Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Data Analysis
- mdlog1 <-glm.nb (DispInvWhole ~ habitat * Hist, data = DE)
- mdlog2 <-glm.nb (DispInvWhole ~ habitat + Hist, data = DE)
3. Results
3.1. General Results
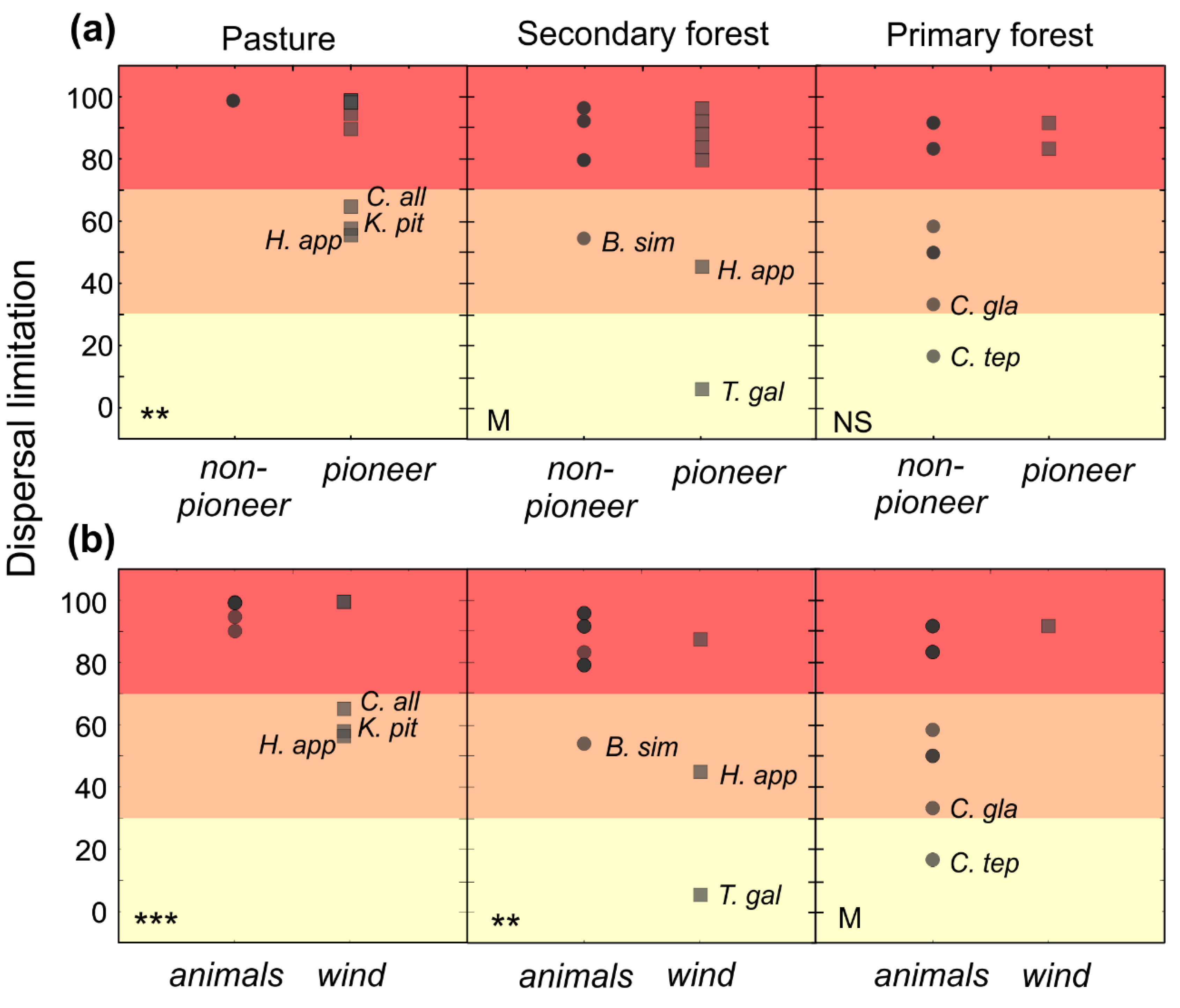
3.2. Dispersal Limitation
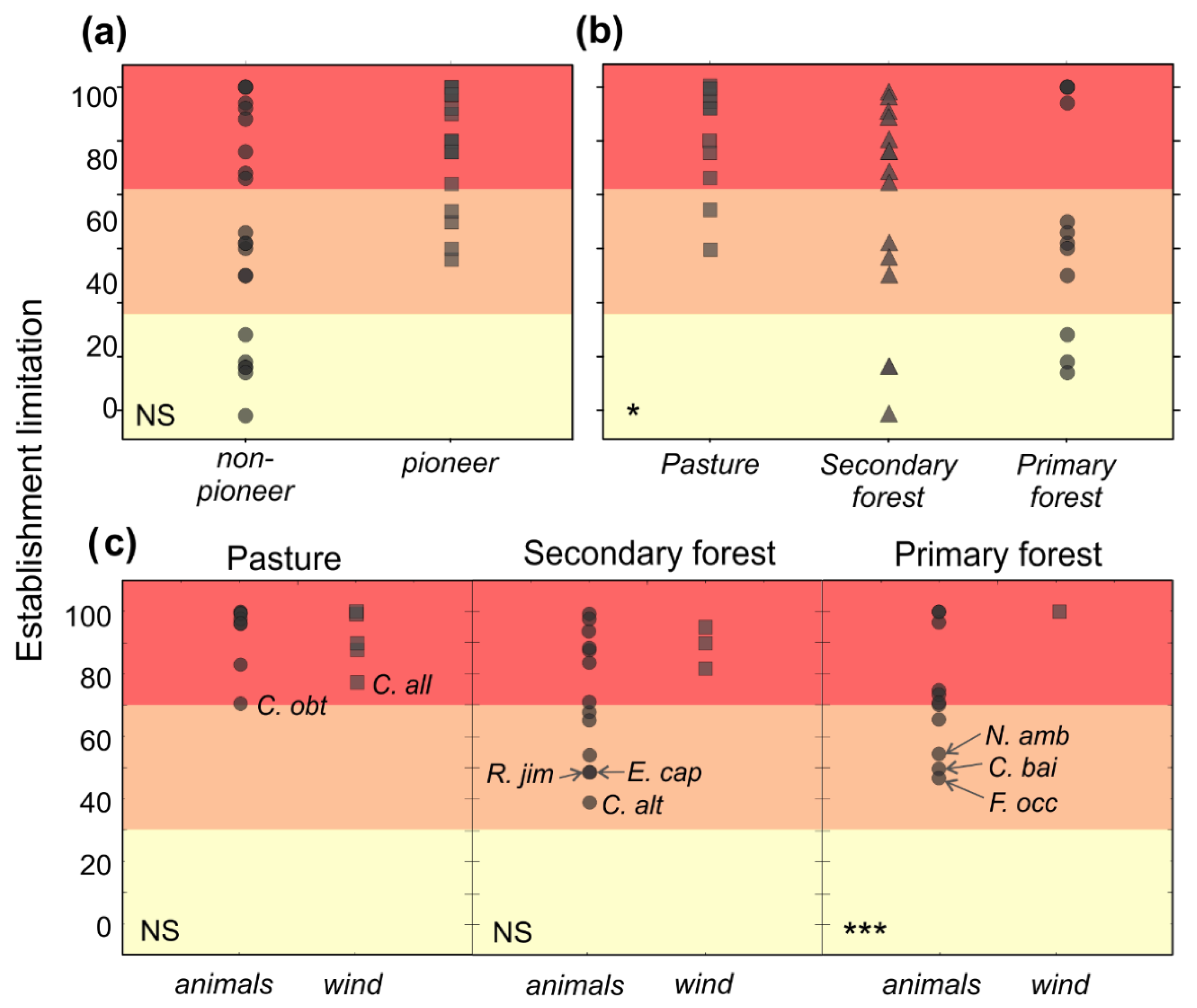
3.3. Establishment Limitation
4. Discussion
4.1. Dispersal Limitation by Habitat
4.2. Establishment Limitation by Habitat
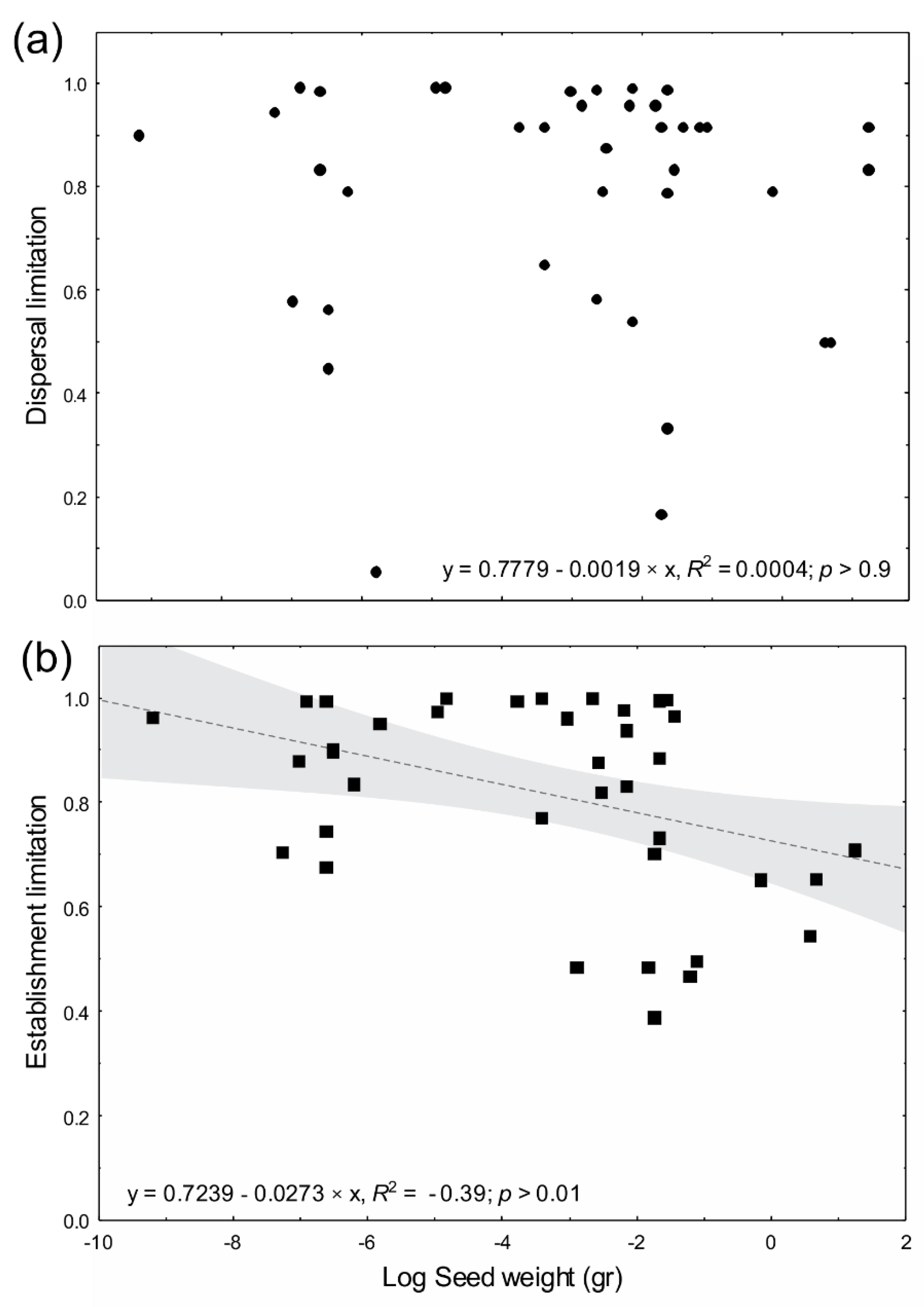
4.3. Limitation by Life History, Dispersal Agent and Seed Size
4.4. Management Implications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wijdeven, S.M.J.; Kuzee, M.E. Seed availability as a limiting factor in forest recovery processes in Costa Rica. Restor. Ecol. 2000, 8, 414–424. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Diversity of neotropical migratory landbird species assemblages in forest fragments and man-made vegetation in Los Tuxtlas, Mexico. Biodivers. Conserv. 2005, 14, 1719–1734. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R. Species composition and reproductive phenology of bats in a tropical landscape at Los Tuxtlas, Mexico. Camb. J. Online 2001, 17, 627–646. [Google Scholar] [CrossRef]
- Laborde, J.; Guevara, S.; Sanchez-Rios, G. Tree and shrub seed dispersal in pastures: The importance of rainforest trees outside forest fragments. EcoScience 2008, 15, 6–16. [Google Scholar] [CrossRef]
- Guevara, S.; Laborde, J. Monitoring seed dispersal at isolated standing trees in tropical pastures: Consequences for local species availability. Vegetatio 1993, 107/108, 319–338. [Google Scholar]
- Nathan, R.; Schurr, F.M.; Spiegel, O.; Steinitz, O.; Trakhtenbrot, A.; Tsoar, A. Mechanisms of long-distance seed dispersal. Trends Ecol. Evolut. 2008, 23, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Garza, C.; González-Montagut, R. Seed rain from forest fragments into tropical pastures in Los Tuxtlas, Mexico. Plant Ecol. 1999, 145, 255–265. [Google Scholar] [CrossRef]
- Bacles, C.F.E.; Lowe, A.J.; Ennos, R.A. Effective seed dispersal across a fragmented landscape. Science 2006, 311, 628. [Google Scholar] [CrossRef] [PubMed]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Clark, C.J.; Poulsen, J.R.; Levey, D.J. Roles of seed and establishment limitation in determining patterns of afrotropical tree recruitment. PLoS ONE 2013, 8, e63330. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.L.; Holl, K.D. Arrival ≠ survival. Restor. Ecol. 2013, 21, 153–155. [Google Scholar] [CrossRef]
- Weiher, E.; Freund, D.; Bunton, T.; Stefanski, A.; Lee, T.; Bentivenga, S. Advances, challenges and a developing synthesis of ecological community assembly theory. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2403–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepstad, D.C.; Uhl, C.; Serrao, E.A.S. Recuperation of a degraded Amazonian landscape: Forest recovery and agricultural restoration. Ambio 1991, 20, 248–255. [Google Scholar]
- Buschbacher, R.J. Tropical deforestation and pasture development. BioScience 1986, 36, 22–28. [Google Scholar] [CrossRef]
- Rhoades, C.C.; Eckert, G.E.; Coleman, D.C. Effect of pasture trees on soil nitrogen and organic matter: Implications for tropical montane forest restoration. Restor. Ecol. 1998, 6, 262–270. [Google Scholar] [CrossRef]
- Myster, R.W. Post-agricultural invasion, establishment, and growth of neotropical trees. Bot. Rev. 2004, 70, 381–402. [Google Scholar] [CrossRef]
- Turner, I.M. The Ecology of Trees in the Tropical Rain Forest; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Kitajima, K. Do shade-tolerant tropical tree seedlings depend longer on seed reserves? Functional growth analysis of three Bignoniaceae species. Funct. Ecol. 2002, 16, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Camargo, J.L.C.; Ferraz, I.D.K.; Imakawa, A.M. Rehabilitation of degraded areas of central Amazonia using direct sowing of forest tree seeds. Restor. Ecol. 2002, 10, 636–644. [Google Scholar] [CrossRef]
- García-Orth, X.; Martínez-Ramos, M. Seed dynamics of early and late successional tree species in tropical abandoned pastures: Seed burial as a way of evading predation. Restor. Ecol. 2008, 16, 435–443. [Google Scholar] [CrossRef]
- Jones, F.A.; Peterson, C.J.; Haines, B.L. Seed predation in neotropical pre-montane pastures: Site, distance, and species effects. Biotropica 2003, 35, 219–225. [Google Scholar] [CrossRef]
- Nepstad, D.; Uhl, C.; Serrão, A.E. Surmounting barriers to forest regeneration in abandoned highly degraded pastures: A case study from Paragominas, Pará, Brazil. In Alternatives to Deforestation: Steps toward Sustainable Use of the Amazon Rain Forest; Anderson, A.B., Ed.; Columbia University Press: New York, NY, USA, 1990; pp. 215–229. [Google Scholar]
- Myster, R.W. Vegetation dynamics of a permanent pasture plot in Puerto Rico. Biotropica 2003, 35, 422–428. [Google Scholar] [CrossRef]
- Sork, V.L. Effects of predation and light on seedling establishment in Gustavia superba. Ecology 1987, 68, 1341–1350. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Scheiner, S.M. Species composition in gaps and structure of a tropical forest. Ecology 1989, 70, 538–541. [Google Scholar] [CrossRef]
- Terborgh, J.; Zhu, K.; Álvarez-Loayza, P.; Cornejo Valverde, F. How many seeds does it take to make a sapling? Ecology 2013, 95, 991–999. [Google Scholar] [CrossRef]
- Kraft, N.J.; Valencia, R.; Ackerly, D.D. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Guevara, S.; Laborde, D.; Sánchez-Ríos, G. Los Tuxtlas: El Paisaje de la Sierra; Instituto de Ecología, A.C.-Unión Europea: Xalapa, Mexico, 2004; p. 288. [Google Scholar]
- Soto, M.; Gama, L. La región de los tuxtlas: Climas. In Historia Natural de Los Tuxtlas; González Soriano, E., Dirzo, R., Vogt, C.R., Eds.; Universidad Nacional Autónoma de México, Instituto de Biología: Mexico City, Mexico, 1997; pp. 7–23. [Google Scholar]
- Tobon, W.; Martinez-Garza, C.; Campo, J. Soil responses to restoration of a tropical pasture in Veracruz, south-eastern Mexico. J. Trop. For. Sci. 2011, 23, 338–344. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México. Rev. de Biol. Trop. 1995, 43, 75–115. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México (Mimosaceae a Verbenaceae). Rev. de Biol. Trop. 1996, 44, 41–60. [Google Scholar]
- Ibarra-Manriquez, G.; Sinaca, S. Lista florística comentada de la Estación de Biología Tropical “Los Tuxtlas”, Veracruz, México (Violaceae a Zingiberaceae). Rev. de Biol. Trop. 1996, 44, 427–447. [Google Scholar]
- Bongers, F.; Popma, J.; Meave-del-Castillo, J.; Carabias, J. Structure and floristic composition on the lowland rain forest of Los Tuxtlas, Mexico. Vegetation 1988, 74, 55–80. [Google Scholar] [CrossRef]
- Estrada, A.; Coates-Estrada, R.; Meritt, D.; Montiel, S.; Curiel, D. Patterns of frugivore species richness and abundance in forest islands and in agricultural habitats at Los Tuxtlas, Mexico. Vegetatio 1993, 107/108, 245–257. [Google Scholar]
- De la Peña-Domene, M.; Howe, H.F.; Cruz-León, E.; Jiménez-Rolland, R.; Lozano-Huerta, C.; Martínez-Garza, C. Seed to seedling transitions in successional habitats across a tropical landscape. Oikos 2017, 126, 410–419. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Martínez-Garza, C.; Howe, H.F. Early recruitment dynamics in tropical restoration. Ecol. Appl. 2013, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- de la Peña-Domene, M. and Martínez-Garza, C. Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests. Forests. 2018, 9, 570. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Flores-Palacios, A.; De La Peña-Domene, M.; Howe, H.F. Seed rain in a tropical agricultural landscape. J. Trop. Ecol. 2009, 25, 541–550. [Google Scholar] [CrossRef]
- Ibarra-Manriquez, G.; Ramos, M.M.; Oyama, K. Seedling functional types in a lowland rain forest in Mexico. Am. J. Bot. 2001, 88, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ramos, M. Claros, ciclos vitales de los árboles tropicales y la regeneración natural de las selvas altas perennifolias. In Investigaciones Sobre la Regeneración de las Selvas Altas en Veracruz, México; Amo, A.G.-P.S.d., Ed.; Editorial Alhambra Mexicana, S.A.: México D.F., Mexico, 1985; pp. 191–239. [Google Scholar]
- Popma, J.; Bongers, F.; Werger, M.J.A. Gap-dependance and leaf characteristics of trees in a tropical lowland rain-forest in Mexico. Oikos 1992, 63, 207–214. [Google Scholar] [CrossRef]
- Kew, R.B.G. Seed Information Database (sid), Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 10 September 2011).
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Hubbell, S.P.; Foster, R.B. Assessing recruitment limitation: Concepts, methods and case-studies from a tropical forest. In Seed Dispersal and Frugivory; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CAB International: Wallingford: Oxon, UK, 2002; pp. 35–53. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- “Statistica”, version 7; Data Analysis Software System; StatSoft Inc.: Tulsa, OK, USA, 2004.
- Caughlin, T.T.; Ferguson, J.M.; Lichstein, J.W.; Bunyavejchewin, S.; Levey, D.J. The importance of long-distance seed dispersal for the demography and distribution of a canopy tree species. Ecology 2013, 95, 952–962. [Google Scholar] [CrossRef]
- Norden, N.; Chave, J.; Belbenoit, P.; Caubere, A.; Chatelet, P.; Forget, P.M.; Riera, B.; Viers, J.; Thebaud, C. Interspecific variation in seedling responses to seed limitation and habitat conditions for 14 neotropical woody species. J. Ecol. 2009, 97, 186–197. [Google Scholar] [CrossRef]
- Cubina, A.; Aide, T.M. The effect of distance from forest edge on seed rain and soil seed bank in a tropical pasture. Biotropica 2001, 33, 260–267. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Pascarella, J.B.; Aide, T.M. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restor. Ecol. 2000, 8, 350–360. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Scariot, A. Principles of natural regeneration of tropical dry forests for restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef]
- Posada, J.M.; Aide, T.M.; Cavelier, J. Cattle and weedy shrubs as restoration tools of tropical montane rainforest. Restor. Ecol. 2000, 8, 370–379. [Google Scholar] [CrossRef]
- Caughlin, T.T.; Elliott, S.; Lichstein, J.W. When does seed limitation matter for scaling up reforestation from patches to landscapes? Ecol. Appl. 2016, 26, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.L.; Holl, K.D.; Zahawi, R.A. Seed dispersal limitations shift over time in tropical forest restoration. Ecol. Appl. 2015, 25, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finegan, B. Pattern and process in neotropical secondary rain forests: The first 100 years of succession. Trends Ecol. Evolut. 1996, 11, 119–124. [Google Scholar] [CrossRef]
- Schupp, E.; Milleron, T.; Russo, S. Dissemination limitation and the origin and maintenance of species-rich tropical forests. In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; Levey, D., Silva, W., Galetti, M., Eds.; CABI Publishing: Oxon, UK, 2002; pp. 19–33. [Google Scholar]
- Engelbrecht, B.M.J.; Wright, S.J.; De Steven, D. Survival and ecophysiology of tree seedlings during El Niño drought in a tropical moist forest in Panama. J. Trop. Ecol. 2002, 18, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Garza, C.; Bongers, F.; Poorter, L. Are functional traits good predictors of species performance in restoration plantings in tropical abandoned pastures? For. Ecol. Manag. 2013, 303, 35–45. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–527. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Pacala, S.W. The consequences of recruitment limitation: reconciling chance, history and competitive differences between plants. J. Theor. Biol. 1995, 176, 1–12. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [PubMed]
- Terborgh, J.; Alvarez-Loayza, P.; Dexter, K.; Cornejo, F.; Carrasco, C. Decomposing dispersal limitation: Limits on fecundity or seed distribution? J. Ecol. 2011, 99, 935–944. [Google Scholar] [CrossRef]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; de Lao, S.L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Popoca-Ortega, L.I. Lluvia de semillas en parcelas de restauración ecológica en la selva tropical de Los Tuxtlas, Veracruz, México. Bachelor Thesis, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos, Mexico, 2016. [Google Scholar]
- Martínez-Ramos, M.; Pingarroni, A.; Rodríguez-Velázquez, J.; Toledo-Chelala, L.; Zermeño-Hernández, I.; Bongers, F. Natural forest regeneration and ecological restoration in human-modified tropical landscapes. Biotropica 2016, 48, 745–757. [Google Scholar] [CrossRef]
- Dalling, J.W.; Muller-Landau, H.C.; Wright, S.J.; Hubbell, S.P. Role of dispersal in the recruitment limitation of neotropical pioneer species. J. Ecol. 2002, 90, 714–727. [Google Scholar] [CrossRef] [Green Version]
- Harms, K.E.; Wright, S.J.; Calderon, O.; Hernandez, A.; Herre, E.A. Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest. Nature 2000, 404, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Dalling, J.W.; Hubbell, S.P. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J. Ecol. 2002, 90, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Seidler, T.G.; Plotkin, J.B. Seed dispersal and spatial pattern in tropical trees. PLoS Biol. 2006, 4, e344. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, N.J.; Howe, H.F. Low recruitment of trees dispersed by animals in African forest fragments. Conserv. Biol. 2001, 15, 1733–1741. [Google Scholar] [CrossRef]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Saavedra, F.; Hensen, I.; Schleuning, M. Deforested habitats lack seeds of late-successional and large-seeded plant species in tropical montane forests. Appl. Veg. Sci. 2015, 18, 603–612. [Google Scholar] [CrossRef]
- Hastings, A. Disturbance, coexistence, history, and competition for space. Theor. Popul. Biol. 1980, 18, 363–373. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Chave, J.; Muller-Landau, H.C.; Levin, S.A. Comparing classical community models: Theoretical consequences for patterns of diversity. Am. Nat. 2002, 159, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Aradottir, A.L.; Halldorsson, G. Colonization of woodland species during restoration: Seed or safe site limitation? Restor. Ecol. 2018, 26, 738. [Google Scholar] [CrossRef]
- Turnbull, L.A.; Crawley, M.J.; Rees, M. Are plant populations seed-limited? A review of seed sowing experiments. Oikos 2000, 88, 225–238. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Luna, A.; Martínez-Garza, C. Performance of 15 tropical tree species recruited or transplanted on restoration settings. Bot. Sci. 2016, 94, 773. [Google Scholar] [CrossRef] [Green Version]
- Grossnickle, S.C.; Ivetić, V. Direct seeding in reforestation—A field performance review. Reforesta 2017, 4, 94–142. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Martínez-Garza, C.; Palmas-Pérez, S.; Rivas-Alonso, E.; Howe, H.F. Roles of birds and bats in early tropical-forest restoration. PLoS ONE 2014, 9, e104656. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Garza, C.; Tobon, W.; Campo, J.; Howe, H.F. Drought mortality of tree seedlings in an eroded tropical pasture. Land Degrad. Dev. 2013, 24, 287–295. [Google Scholar] [CrossRef]
- de la Peña-Domene, M.; Minor, E.S.; Howe, H.F. Restored connectivity facilitates recruitment by an endemic large-seeded tree in a fragmented tropical landscape. Ecology 2016, 97, 2511–2517. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Habitat | Seeds | Seedlings | ||
|---|---|---|---|---|
| Species | Seeds | Species | Seedlings | |
| Pasture | 128 | 29,296 | 35 | 292 |
| Secondary forest | 115 | 81,725 | 35 | 261 |
| Primary forest | 162 | 6160 | 19 | 593 |
| Species | Family | Life History | Dispersal Mode | Seed Mass | Dispersal Limitation | Establishment Limitation |
|---|---|---|---|---|---|---|
| PRIMARY FOREST | ||||||
| Astrocaryum mexicanum Liebm. ex Mart. | ARECACEAE | Non-pioneer | Animal | 3.422 | 0.833 | 0.706 |
| Cymbopetalum baillonii R.E. Fr. | ANNONACEAE | Non-pioneer | Animal | 0.3335 | 0.917 | 0.495 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.333 | 0.731 |
| Chamaedorea tepejilote Liebm. | ARECACEAE | Non-pioneer | Animal | 0.175 | 0.167 | 0.701 |
| Faramea occidentalis (L.) A. Rich. | RUBIACEAE | Non-pioneer | Animal | 0.3 | 0.917 | 0.466 |
| Nectandra ambigens (S.F. Blake) C.K. Allen | LAURACEAE | Non-pioneer | Animal | 1.8 | 0.5 | 0.542 |
| Notopleura chapensis (Steyerm.) C.M. Taylor | RUBIACEAE | Non-pioneer | Animal | 0.21 | 0.833 | 0.996 |
| Trichilia martiana C. DC. | MELIACEAE | Non-pioneer | Animal | 0.2364 | 0.917 | 0.966 |
| Virola guatemalensis (Hemsl.) Warb. | MYRISTICACEAE | Non-pioneer | Animal | 0.1962 | 0.5 | 0.653 |
| Stemmadenia donnell-smithii (Rose) Woodson | APOCYNACEAE | Pioneer | Animal | 0.07 | 0.583 | 0.999 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.833 | 0.746 |
| Cordia alliodora (Ruiz & Pav.) Oken | BORAGINACEAE | Pioneer | Wind | 0.033 | 0.917 | 0.999 |
| SECONDARY FOREST | ||||||
| Astrocaryum mexicanum Liebm. ex Mart. | ARECACEAE | Non-pioneer | Animal | 3.422 | 0.917 | 0.71 |
| Bursera simaruba (L.) Sarg. | BURSERACEAE | Non-pioneer | Animal | 0.117 | 0.54 | 0.938 |
| Cojoba arborea (L.) Britton & Rose | FABACEAE | Non-Pioneer | Animal | 0.85 | 0.791 | 0.652 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.789 | 0.884 |
| Chamaedorea alternans H. Wendl. | ARECACEAE | Non-pioneer | Animal | 0.175 | 0.917 | 0.387 |
| Eugenia capuli (Schltdl. & Cham.) Hook. & Arn. | MYRTACEAE | Non-pioneer | Animal | 0.056 | 0.958 | 0.484 |
| Psychotria veracruzensis Lorence & Dwyer | RUBIACEAE | Non-pioneer | Animal | 0.002 | 0.792 | 0.835 |
| Rollinia jimenezii Saff. | ANNONACEAE | Non-pioneer | Animal | 0.16 | 0.958 | 0.484 |
| Carica papaya L. | CARICACEAE | Pioneer | Animal | 0.023 | 0.917 | 0.992 |
| Hampea nutricia Fryxell | MALVACEAE | Pioneer | Animal | 0.11 | 0.958 | 0.976 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.833 | 0.677 |
| Cnidoscolus multilobus (Pax) I.M. Johnst. | EUPHORBIACEAE | pioneer | Animal | 0.08 | 0.875 | 0.817 |
| Heliocarpus appendiculatus Turcz. | MALVACEAE | Pioneer | Wind | 0.0015 | 0.45 | 0.901 |
| Trichospermum galeottii (Turcz.) Kosterm. | MALVACEAE | Pioneer | Wind | 0.003 | 0.056 | 0.95 |
| Vachellia cornigera (L.) Seigler & Ebinger | FABACEAE | Pioneer | Wind | 0.076 | 0.792 | 0.876 |
| PASTURE | ||||||
| Bursera simaruba (L.) Sarg. | BURSERACEAE | Non-pioneer | Animal | 0.117 | 0.992 | 0.829 |
| Cupania glabra Sw. | SAPINDACEAE | Non-pioneer | Animal | 0.19 | 0.99 | 0.995 |
| Tetrorchidium rotundatum Standl. | EUPHORBIACEAE | Non-pioneer | Animal | 0.048 | 0.987 | 0.961 |
| Cecropia obtusifolia Bertol. | URTICACEAE | Pioneer | Animal | 0.001 | 0.946 | 0.704 |
| Conostegia xalapensis (Bonpl.) D. Don ex DC. | MELASTOMATACEAE | Pioneer | Animal | 0.0001 | 0.9 | 0.962 |
| Piper amalago L. | PIPERACEAE | Pioneer | Animal | 0.0015 | 0.988 | 0.992 |
| Stemmadenia donnell-smithii (Rose) Woodson | APOCYNACEAE | Pioneer | Animal | 0.07 | 0.989 | 0.998 |
| Trema micrantha (L.) Blume | CANNABACEAE | Pioneer | Animal | 0.007 | 0.994 | 0.975 |
| Cordia alliodora (Ruiz & Pav.) Oken | BORAGINACEAE | Pioneer | Wind | 0.033 | 0.651 | 0.771 |
| Koanophyllon pittieri (Klatt) R.M. King & H. Robinson | ASTERACEAE | Pioneer | Wind | 0.0009 | 0.579 | 0.877 |
| Heliocarpus appendiculatus Turcz. | MALVACEAE | Pioneer | Wind | 0.0015 | 0.563 | 0.897 |
| Heliocarpus donnellsmithii Rose | MALVACEAE | Pioneer | Wind | 0.001 | 0.994 | 0.993 |
| Ochroma pyramidale (Cav. ex Lam.) Urb. | MALVACEAE | Pioneer | Wind | 0.008 | 0.994 | 0.998 |
| Interaction | Dispersal Limitation | Establishment Limitation | ||||||
|---|---|---|---|---|---|---|---|---|
| Habitat by Life History | ||||||||
| Estimate | Error | z Value | p Value | Estimate | Error | z Value | p Value | |
| (Intercept) | 2.33 | 0.59 | 3.95 | <0.001 | 8.46 | 2.86 | 2.96 | 0.003 |
| Habitat-Primary forest | 3.52 | 0.67 | 5.28 | <0.001 | 8.10 | 4.19 | 1.93 | 0.05 |
| Habitat-Secondary forest | 2.78 | 0.68 | 4.06 | <0.001 | 9.65 | 4.70 | 2.05 | 0.04 |
| Life history-pioneer | 2.61 | 0.67 | 3.91 | <0.001 | 0.66 | 3.32 | 0.20 | 0.84 |
| Habitat-PrimFor: Hist-pioneer | −3.64 | 1.01 | −3.60 | <0.001 | −5.94 | 6.49 | −0.91 | 0.36 |
| Habitat-SecFor: Hist-pioneer | −2.01 | 0.84 | −2.40 | 0.02 | −8.01 | 5.54 | −1.45 | 0.14 |
| Habitat Type by Dispersal Mode | ||||||||
| Estimate | Error | z Value | p Value | Estimate | Error | z Value | p Value | |
| (Intercept) | 3.29 | 0.31 | 10.31 | <0.001 | 8.50 | 1.70 | 5.014 | <0.001 |
| Habitat-Primary forest | 2.52 | 0.41 | 6.08 | <0.001 | 8.00 | 3.27 | 2.433 | 0.01 |
| Habitat-Secondary forest | 1.74 | 0.40 | 4.27 | <0.001 | 7.54 | 3.12 | 2.420 | 0.01 |
| Dispersal mode-wind | 2.21 | 0.50 | 4.36 | <0.001 | 1.09 | 2.97 | 0.367 | 0.71 |
| Habitat-PrimFor: Disp-wind | −3.60 | 1.05 | −3.41 | <0.001 | −16.60 | 4.14 | −4.005 | <0.001 |
| Habitat-SecFor: Disp-wind | −0.95 | 0.76 | −1.25 | 0.2 | −6.65 | 5.22 | −1.274 | 0.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña-Domene, M.D.l.; Martínez-Garza, C.; Ayestarán-Hernández, L.M.; Howe, H.F. Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes. Forests 2018, 9, 620. https://doi.org/10.3390/f9100620
Peña-Domene MDl, Martínez-Garza C, Ayestarán-Hernández LM, Howe HF. Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes. Forests. 2018; 9(10):620. https://doi.org/10.3390/f9100620
Chicago/Turabian StylePeña-Domene, Marinés De la, Cristina Martínez-Garza, Luz María Ayestarán-Hernández, and Henry F. Howe. 2018. "Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes" Forests 9, no. 10: 620. https://doi.org/10.3390/f9100620
APA StylePeña-Domene, M. D. l., Martínez-Garza, C., Ayestarán-Hernández, L. M., & Howe, H. F. (2018). Plant Attributes that Drive Dispersal and Establishment Limitation in Tropical Agricultural Landscapes. Forests, 9(10), 620. https://doi.org/10.3390/f9100620