Changes in Sensitivity of Tree-Ring Widths to Climate in a Tropical Moist Forest Tree in Bangladesh



Abstract
:1. Introduction
2. Materials and Methods
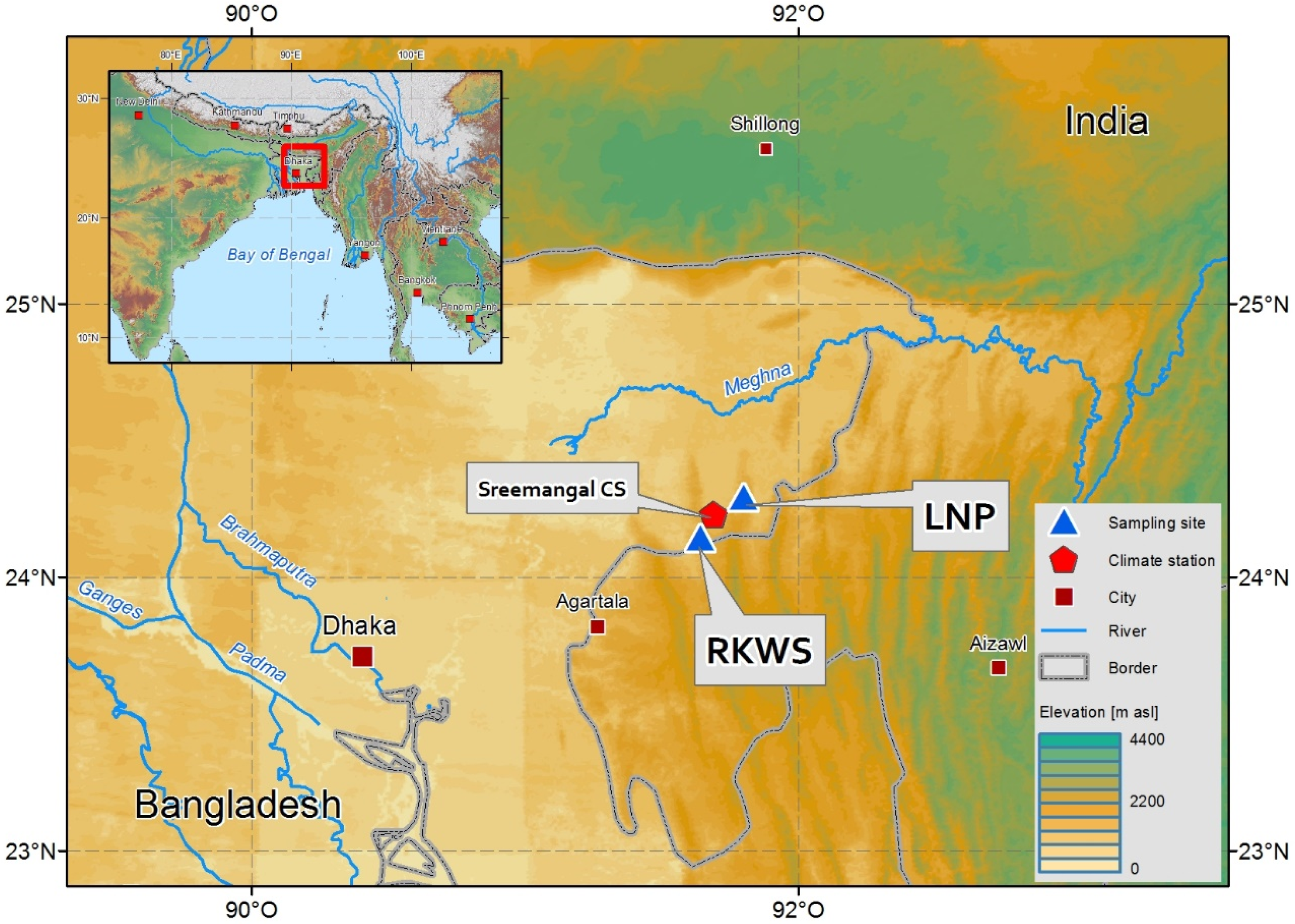
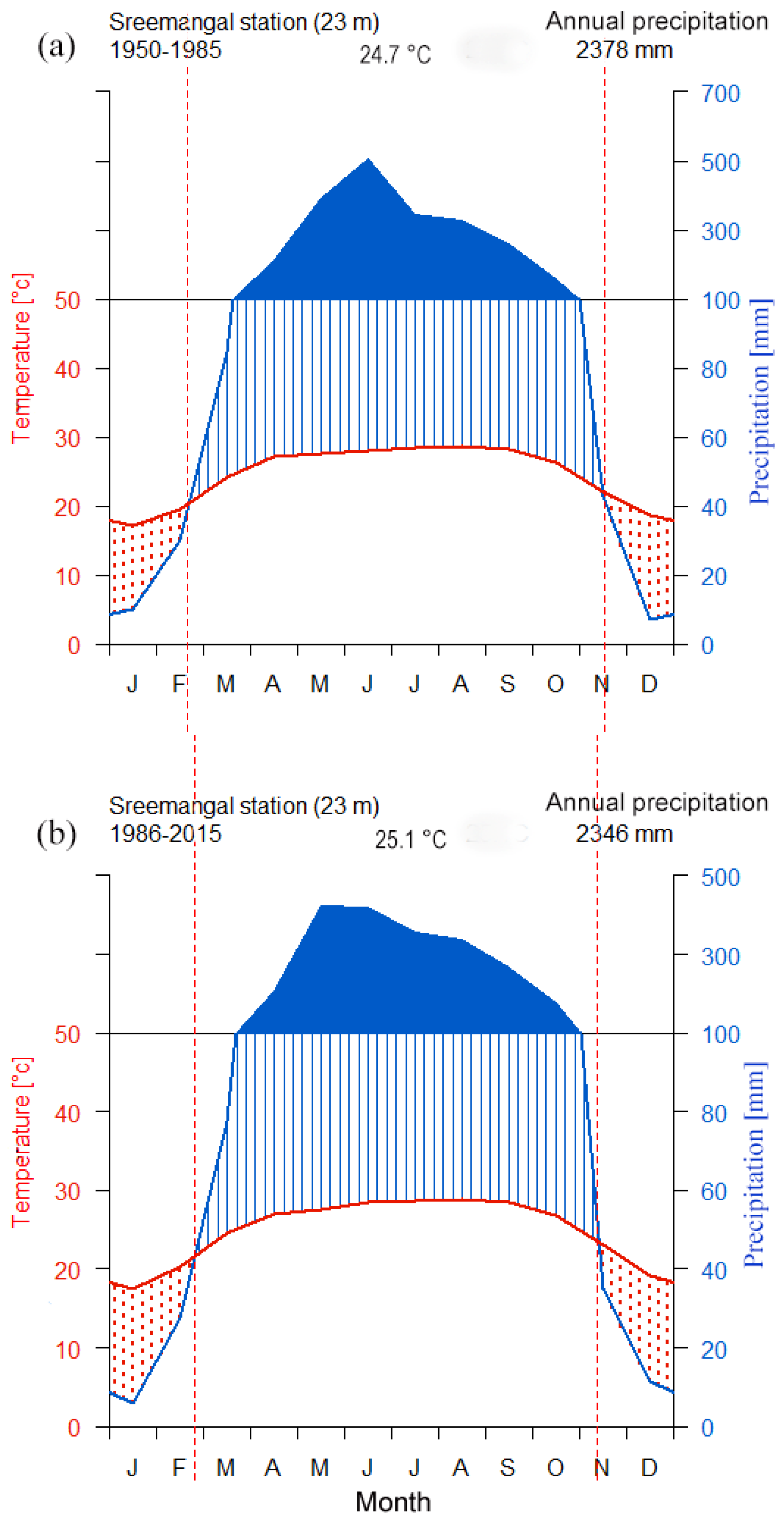
2.1. Study Site, Tree Species, and Local Climate
2.2. Wood Sample Collection and Preparation
2.3. Crossdating and Chronology Development
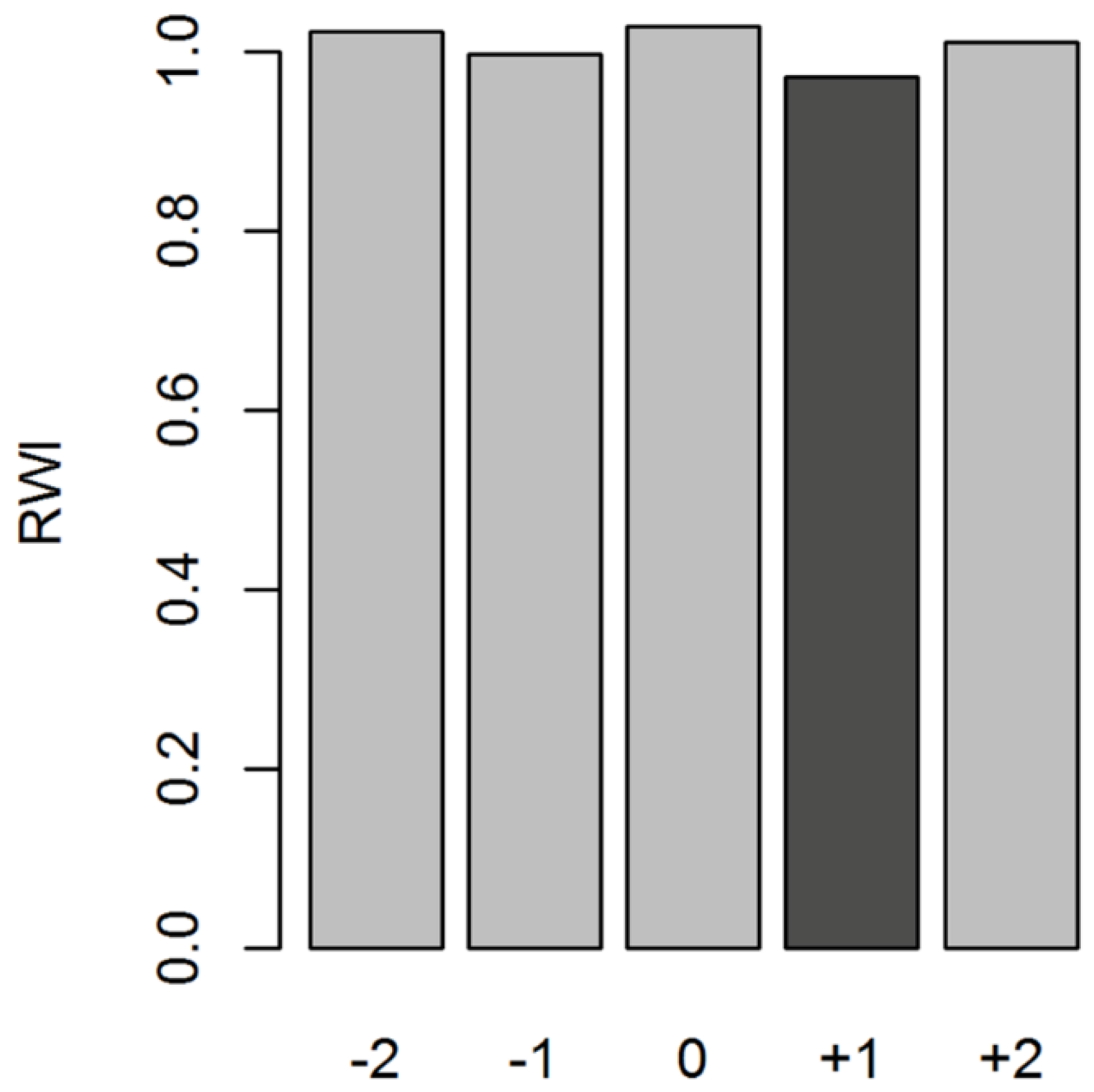
2.4. Data Analyses
3. Results
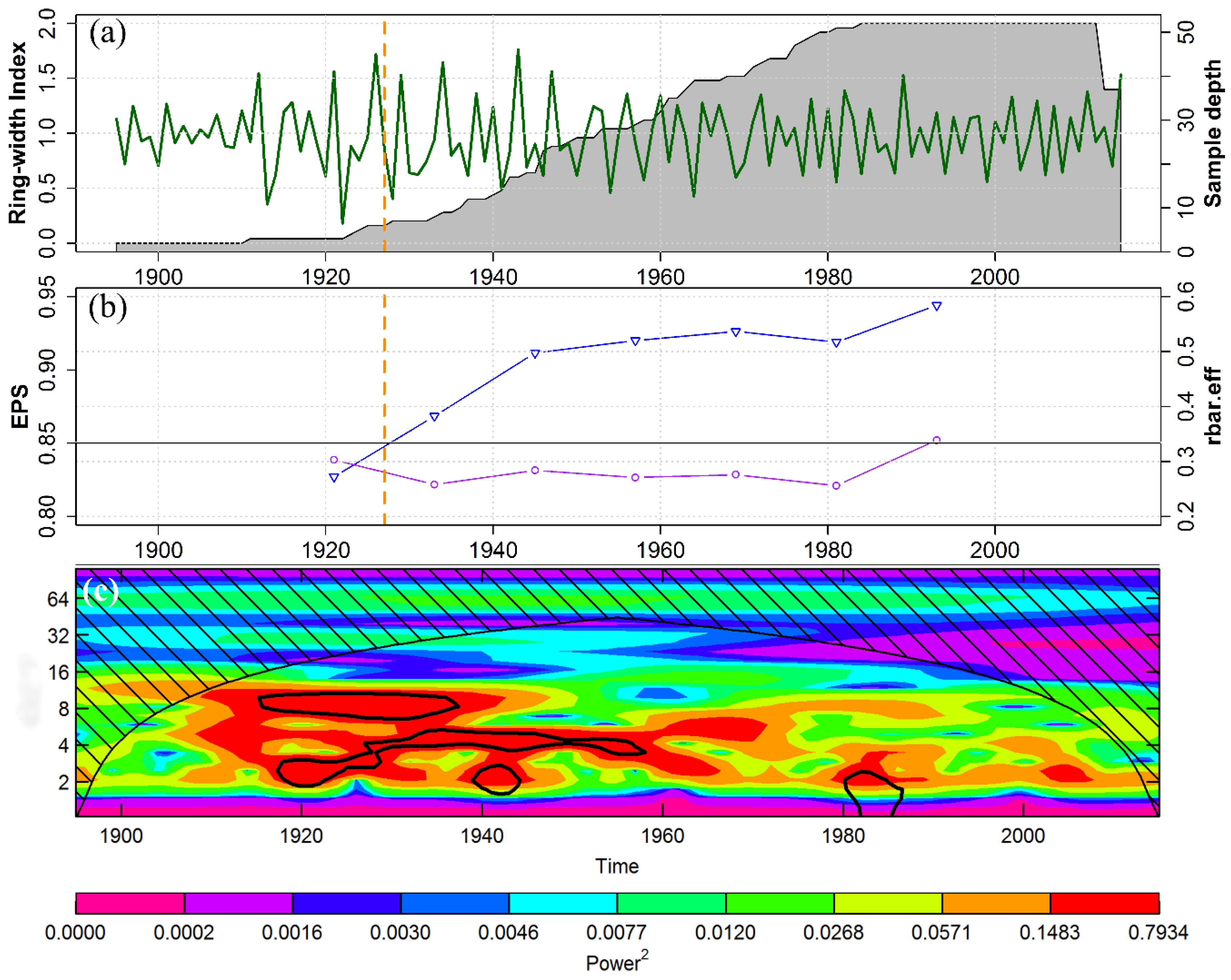
3.1. Chronology Characteristics
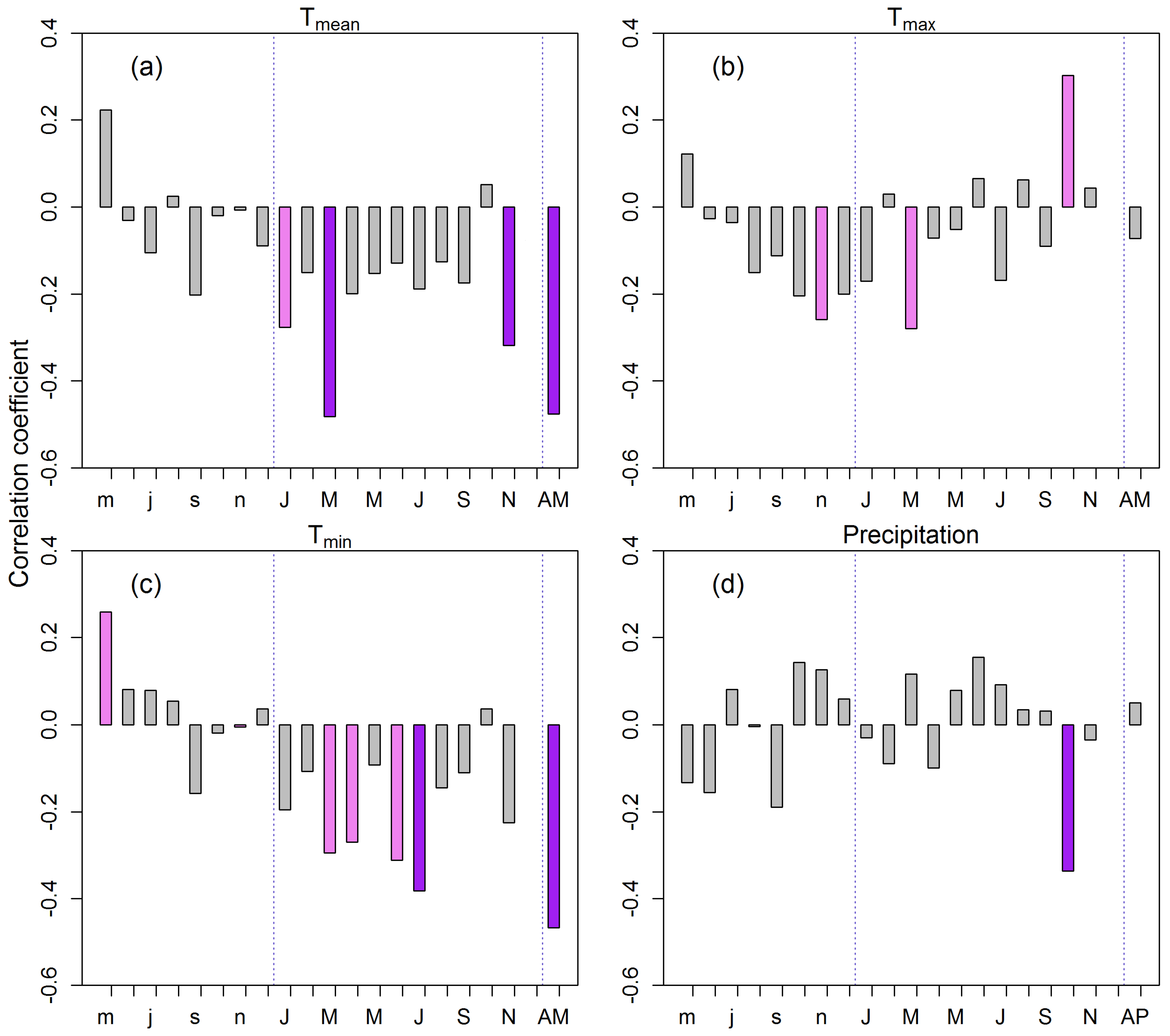
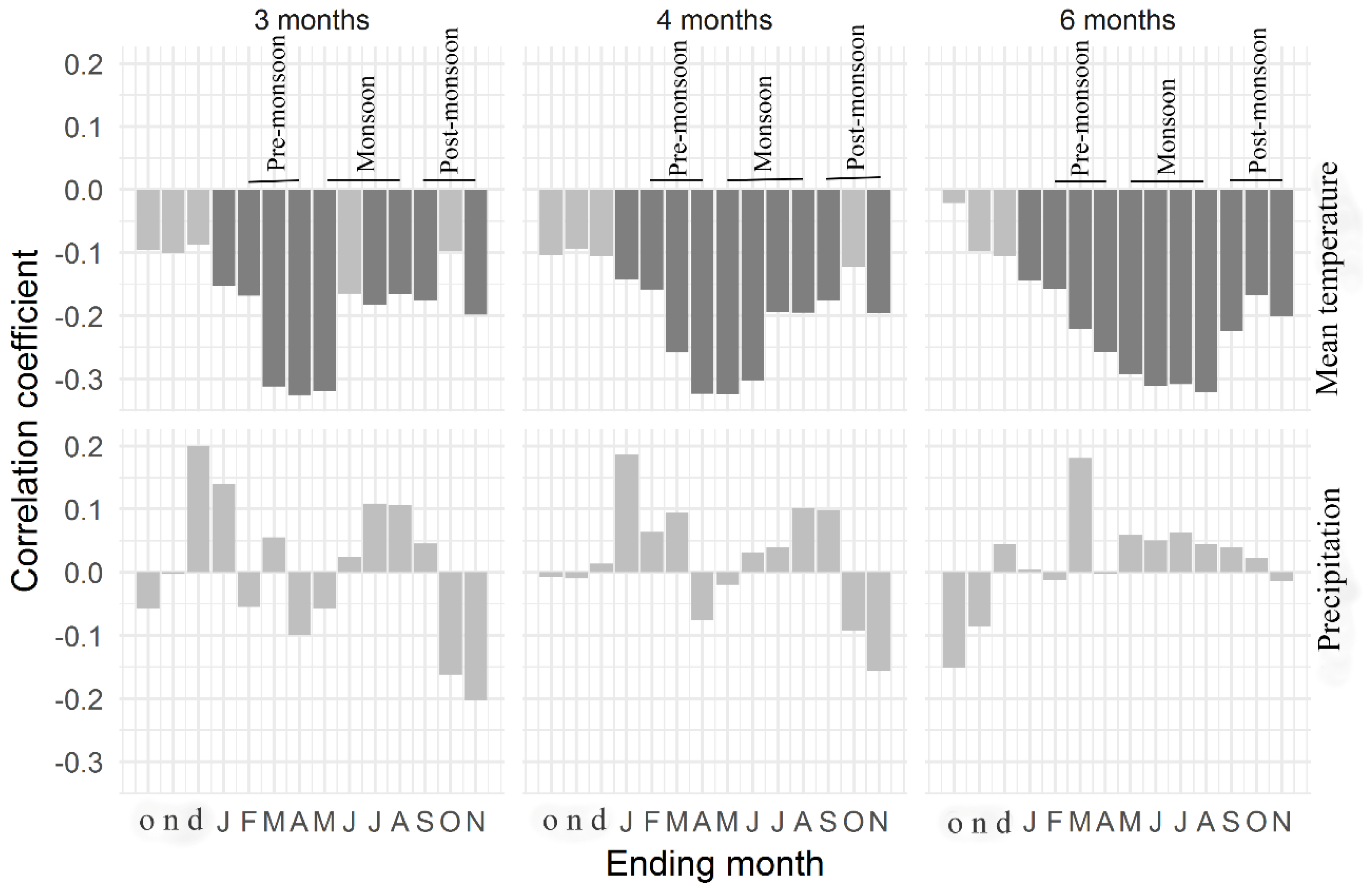
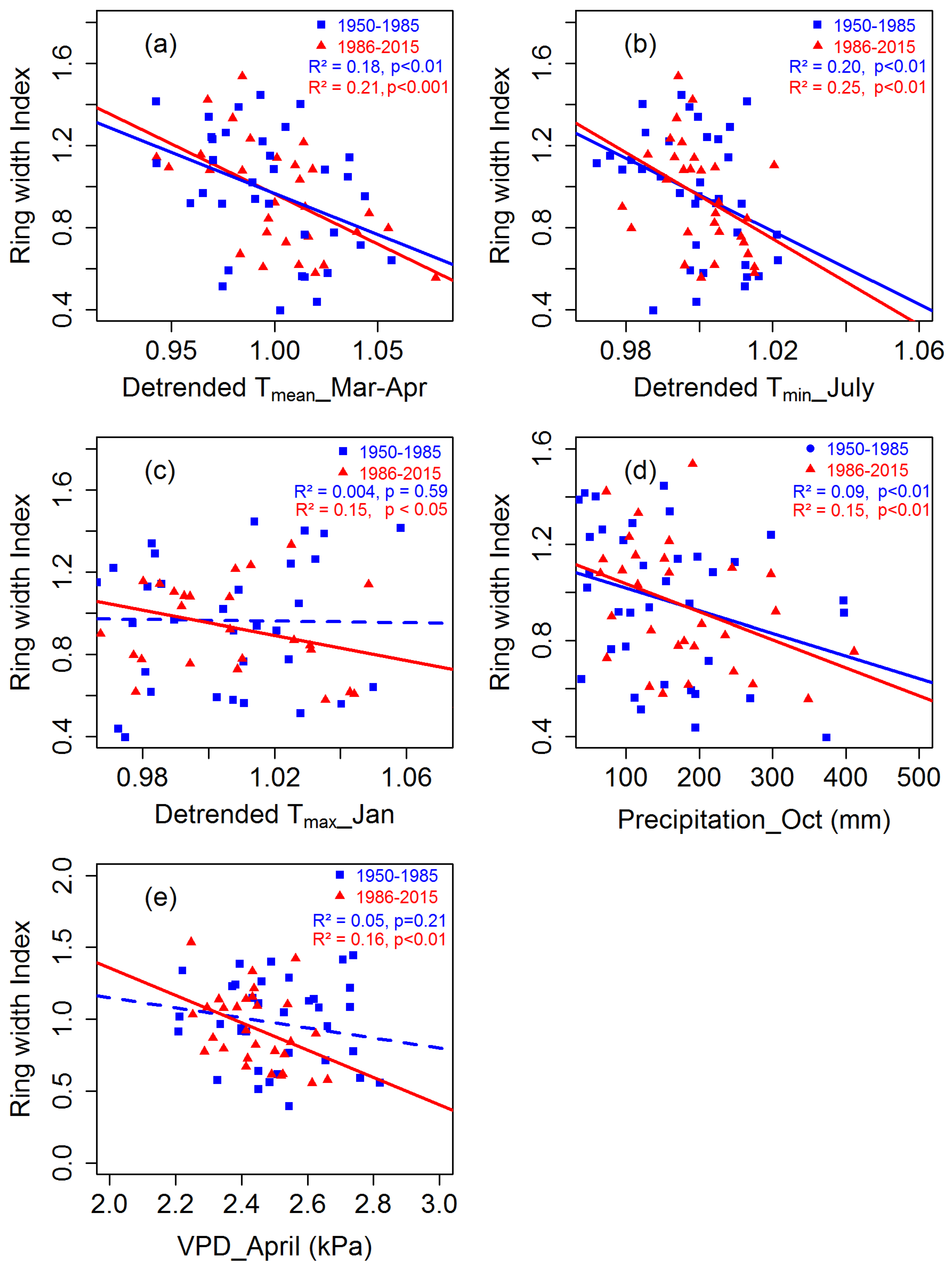
3.2. Tree Growth Responses to Climate Variables
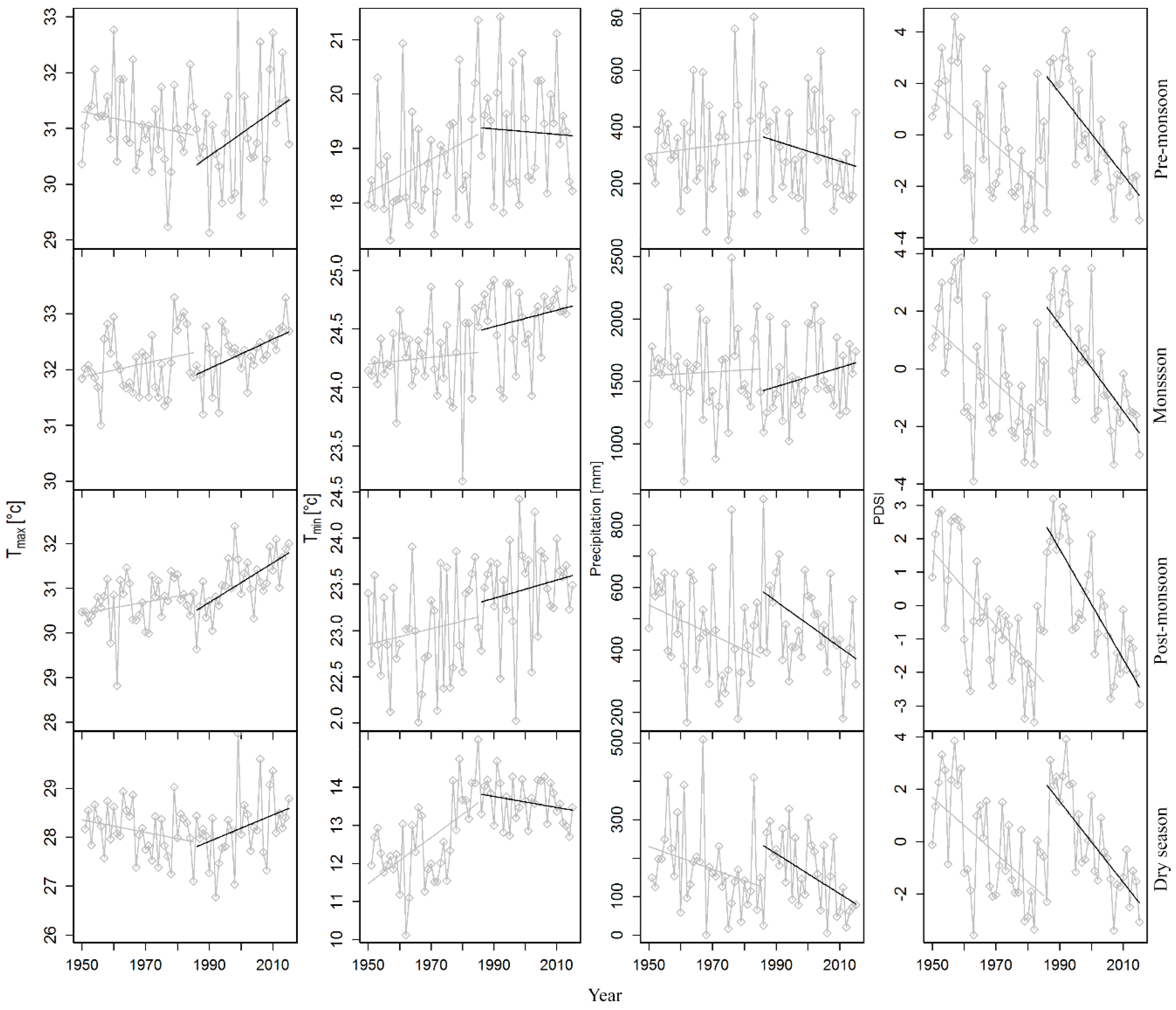
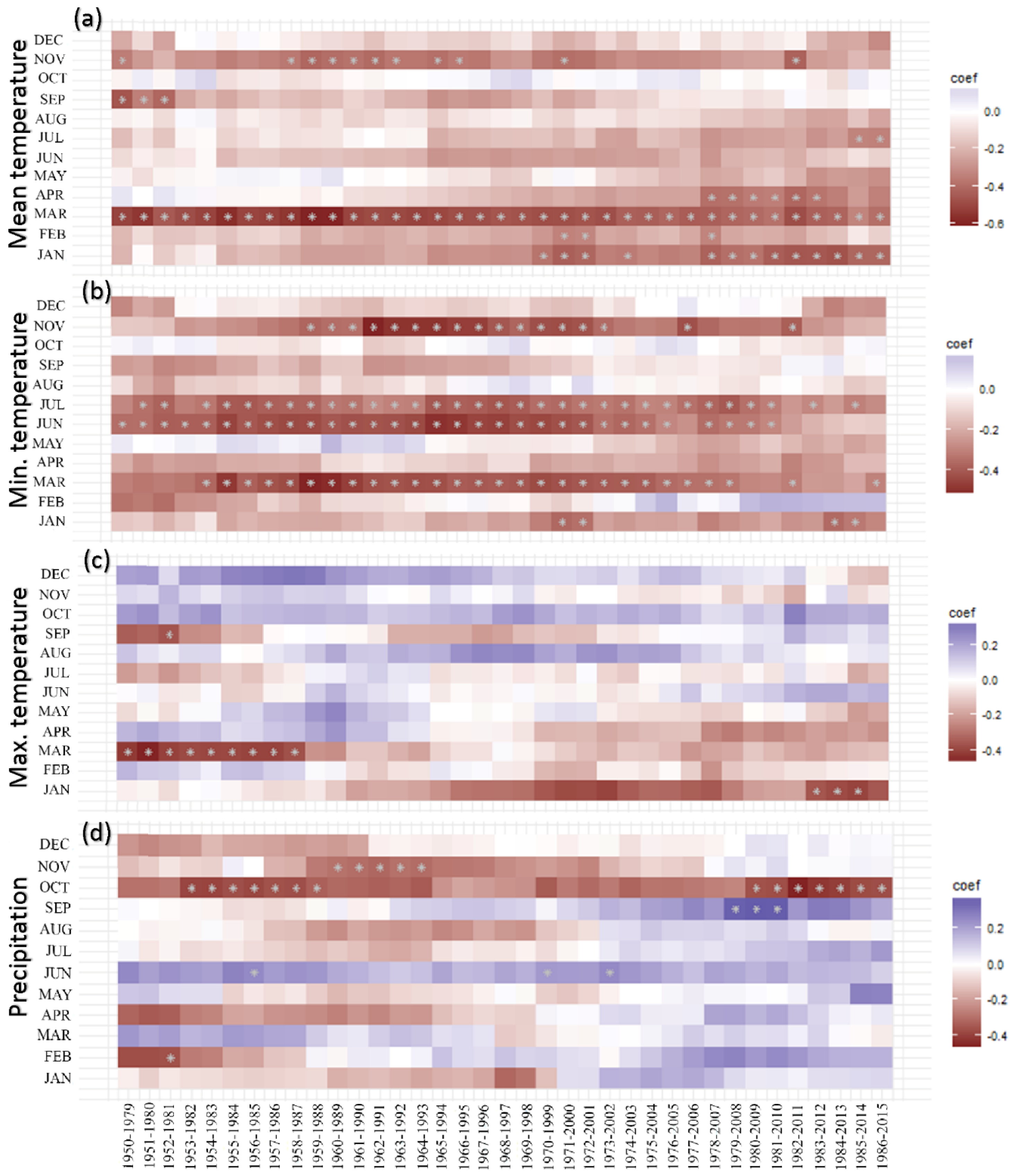
3.3. Trends in Tree Growth Sensitivity to Climate Forcing
4. Discussion
4.1. Signal Strength of the RWI Chronology
4.2. Climate–Growth Relationships
4.3. Stationarity and Consistency in Tree Growth Sensitivity to Climatic Changes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rodenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Grace, J. Understanding and managing the global carbon cycle. J. Ecol. 2004, 92, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Houghton, R.A.; Hall, F.; Goetz, S.J. Importance of biomass in the global carbon cycle. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Le Quéré, C.; Raupach, M.R.; Canadell, J.G.; Al, G.M. Trends in the sources and sinks of carbon dioxide. Nat. Geosci. 2009, 2, 831–836. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Lugo, A.E. Trailblazing the carbon cycle of tropical forests from puerto rico. Forests 2017, 8, 101. [Google Scholar] [CrossRef]
- Djomo, A.N.; Knohl, A.; Gravenhorst, G. Estimations of total ecosystem carbon pools distribution and carbon biomass current annual increment of a moist tropical forest. For. Ecol. Manag. 2011, 261, 1448–1459. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Clare, S.; Mack, M.C.; Brooks, M. A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest. Ecology 2013, 94, 1540–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doughty, C.E.; Metcalfe, D.B.; Girardin, C.A.J.; Amezquita, F.F.; Durand, L.; Huaraca Huasco, W.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, M.C.; da Costa, A.C.L.; et al. Source and sink carbon dynamics and carbon allocation in the Amazon basin. Glob. Biogeochem. Cycles 2015, 29, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.; Mitchard, E.; Gloor, E. Perturbations in the carbon budget of the tropics. Glob. Chang. Biol. 2014, 20, 3238–3255. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonké, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A.; et al. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003–1006. [Google Scholar] [CrossRef]
- Wright, S.J. Tropical forests in a changing environment. Trends Ecol. Evol. 2005, 20, 553–560. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Lebrija-Trejos, E.; Zuidema, P.A.; Martínez-Ramos, M. Climate-growth analysis for a Mexican dry forest tree shows strong impact of sea surface temperatures and predicts future growth declines. Glob. Chang. Biol. 2010, 16, 2001–2012. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Zuidema, P.A. Relating tree growth to rainfall in Bolivian rain forests: A test for six species using tree ring analysis. Oecologia 2005, 146, 1–12. [Google Scholar] [CrossRef]
- Couralet, C.; Sterck, F.J.; Sass-Klaassen, U.; Van Acker, J.; Beeckman, H. Species-specific growth responses to climate variations in understory trees of a central african rain forest. Biotropica 2010, 42, 503–511. [Google Scholar] [CrossRef]
- Dünisch, O.; Montóia, V.R.; Bauch, J. Dendroecological investigations on Swietenia macrophylla King and Cedrela odorata L. (Meliaceae) in the central Amazon Abstract. Trees-Struct. Funct. 2003, 17, 244–250. [Google Scholar]
- Gebrekirstos, A.; Mitlöhner, R.; Teketay, D.; Worbes, M. Climate-growth relationships of the dominant tree species from semi-arid savanna woodland in Ethiopia. Trees-Struct. Funct. 2008, 22, 631–641. [Google Scholar] [CrossRef]
- Locosselli, G.M.; Schöngart, J.; Ceccantini, G. Climate/growth relations and teleconnections for a Hymenaea courbaril (Leguminosae) population inhabiting the dry forest on karst. Trees-Struct. Funct. 2016, 30, 1127–1136. [Google Scholar] [CrossRef]
- Pucha-Cofrep, D.; Peters, T.; Bräuning, A. Wet season precipitation during the past century reconstructed from tree-rings of a tropical dry forest in Southern Ecuador. Glob. Planet. Chang. 2015, 133, 65–78. [Google Scholar] [CrossRef]
- Schöngart, J.; Orthmann, B.; Hennenberg, K.J.; Porembski, S.; Worbes, M. Climate-growth relationships of tropical tree species in West Africa and their potential for climate reconstruction. Glob. Chang. Biol. 2006, 12, 1139–1150. [Google Scholar] [CrossRef]
- Trouet, V.; Coppin, P.; Beeckman, H. Annual growth ring patterns in Brachystegia spiciformis reveal influence of precipitation on tree growth. Biotropica 2006, 38, 375–382. [Google Scholar] [CrossRef]
- Battipaglia, G.; Zalloni, E.; Castaldi, S.; Marzaioli, F.; Cazzolla-Gatti, R.; Lasserre, B.; Tognetti, R.; Marchetti, M.; Valentini, R. Long tree-ring chronologies provide evidence of recent tree growth decrease in a central african tropical forest. PLoS ONE 2015, 10, e0126168. [Google Scholar] [CrossRef]
- Chambers, J.; Silver, W. Some aspects of ecophysiological and biogeochemical responses of tropical forests to atmospheric change. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.B.; Clark, D.A.; Oberbauer, S.F. Annual wood production in a tropical rain forest in NE Costa Rica linked to climatic variation but not to increasing CO2. Glob. Chang. Biol. 2010, 16, 747–759. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, M.; Bräuning, A. Local and regional climatic signals recorded in tree-rings of Chukrasia tabularis in Bangladesh. Dendrochronologia 2017, 45, 1–11. [Google Scholar] [CrossRef]
- Feeley, K.J.; Wright, S.J.; Supardi, M.N.N.; Kassim, A.R.; Davies, S.J. Decelerating growth in tropical forest trees. Ecol. Lett. 2007, 10, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Brienen, R.J.W.; Zuidema, P.A.; Martínez-Ramos, M. Attaining the canopy in dry and moist tropical forests: Strong differences in tree growth trajectories reflect variation in growing conditions. Oecologia 2010, 163, 485–496. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragao, L.E.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; Lopez-Gonzalez, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef] [Green Version]
- Vincent, G.; de Foresta, H.; Mulia, R. Co-occurring tree species show contrasting sensitivity to ENSO-related droughts in planted dipterocarp forests. For. Ecol. Manag. 2009, 258, 1316–1322. [Google Scholar] [CrossRef]
- Briffa, K.R.; Schweingruber, F.H.; Jones, P.D.; Osborn, T.J.; Shiyatov, S.G.; Vaganov, E.A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 1998, 391, 678–682. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Long-term change in the sensitivity of tree-ring growth to climate forcing in Larix decidua. New Phytol. 2006, 170, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Biondi, F. Are climate-tree growth relationships changing in North-Central Idaho, USA? Arct. Antarct. Alp. Res. 2000, 32, 111–116. [Google Scholar] [CrossRef]
- Smith, K.T.; Cufar, K.; Levanic, T. Temporal stability and dendroclimatology in silver fir and red spruce. Phyton Ann. Rei Bot. Austria 1999, 39, 117–122. [Google Scholar]
- Solberg, B.O.; Hofgaard, A.; Hytteborn, H. Shifts in radial growth responses of coastal Picea abies induced by climatic change during the 20th century, central Norway. Ecoscience 2002, 9, 79–88. [Google Scholar] [CrossRef]
- Wilmking, M.; Juday, G.P.; Barber, V.A.; Zald, H.S.J. Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Glob. Chang. Biol. 2004, 10, 1724–1736. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Hughes, M.K.; Kirdyanov, A.V.; Schweingruber, F.H.; Silkin, P.P. Influence of snowfall and melt timing on tree growth in subarctic Eurasia. Nature 1999, 400, 149–151. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Ohta, T.; Maximov, T.C. Growth and physiological responses of larch trees to climate changes deduced from tree-ring widths and δ13C at two forest sites in eastern Siberia. Polar Sci. 2014, 8, 183–195. [Google Scholar] [CrossRef]
- Huq, S.; Ayer, J. Climate Change Impacts and Responses in Bangladesh; A Report of European Parliament, IP/A/CLIM/IC/2007-106; European Parliament: Brussels, Belgium, 2008. [Google Scholar]
- Huq, S.; Reid, H.; Konate, M.; Rahman, A.; Sokona, Y.; Crick, F. Mainstreaming adaptation to climate change in Least Developed Countries (LDCs). Clim. Policy 2004, 4, 25–43. [Google Scholar] [CrossRef]
- IPCC Climate Change. The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Basak, J.K.; Titumir, R.A.M.; Dey, N.C. Climate change in Bangladesh: A Historical analysis of temperature and rainfall data. J. Environ. 2013, 2, 41–46. [Google Scholar]
- Rahman, M.R.; Lateh, H. Climate change in Bangladesh: A spatio-temporal analysis and simulation of recent temperature and rainfall data using GIS and time series analysis model. Theor. Appl. Climatol. 2017, 128, 27–41. [Google Scholar] [CrossRef]
- Rahman, M.R.; Lateh, H. Meteorological drought in Bangladesh: Assessing, analysing and hazard mapping using SPI, GIS and monthly rainfall data. Environ. Earth Sci. 2016, 75, 1026. [Google Scholar] [CrossRef]
- Shahid, S. Rainfall variability and the trends of wet and dry periods in Bangladesh. Int. J. Climatol. 2010, 30, 2299–2313. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Islam, M.; Bräuning, A. Tree radial growth is projected to decline in South Asian moist forest trees under climate change. Glob. Planet. Chang. 2018, 170, 106–119. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Brienen, R.J.W.; Gloor, E.; Phillips, O.L.; Prior, L.D. Detecting trends in tree growth: Not so simple. Trends Plant Sci. 2013, 18, 11–17. [Google Scholar] [CrossRef]
- Chowdhury, M.Q.; De Ridder, M.; Beeckman, H. Climatic signals in tree rings of heritiera fomes Buch.-Ham. in the Sundarbans, Bangladesh. PLoS ONE 2016, 11, e0149788. [Google Scholar] [CrossRef]
- Worbes, M. One hundred years of tree-ring research in the tropics—A brief history and an outlook to future challenges. Dendrochronologia 2002, 20, 217–231. [Google Scholar] [CrossRef]
- Islam, M.; Rahman, M.; Bräuning, A. Growth-ring boundary anatomy and dendrochronological potential in a moist triopical forest in Northeastern Bangladesh. Tree-Ring Res. 2018, 74, 76–93. [Google Scholar] [CrossRef]
- Nishat, A.; Huq, S.M.I.; Barua, S.P.; Reza, A.H.M.A.; Khan, A.S.M. (Eds.) Bio-ecological Zones of Bangladesh; IUCN Bangladesh Country Office: Dhaka, Bangladesh, 2002. [Google Scholar]
- Hassan, M. Forest soils of Bangladesh. J. For. Sci. 1994, 23, 1–11. [Google Scholar]
- FSP First Five Year Management Plan for Lawachara National Park; Volume 1: Management Plan; Forest Department, Ministry of Environment and Forests: Dhaka, Bangladesh, 2000.
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, CA, USA, 1967. [Google Scholar]
- Islam, M.; Feeroz, M.M.; Muzaffar, S.B.; Kabir, M.; Begum, S.; Hassan, K.; Mahmud, S.; Chakma, S. Population Status and Conservation of hoolock gibbons Hylobates hoolock Harlan 1834 in Bangladesh. J. Bombay Nat. Hist. Soc. 2008, 105, 19–23. [Google Scholar]
- Islam, M.; Salim, S.H.; Kawsar, M.H.; Rahman, M. The effect of soil moisture content and forest canopy openness on the regeneration of Dipterocarpus turbinatus C.F. Gaertn. (Dipterocarpaceae) in a protected forest area of Bangladesh. Trop. Ecol. 2016, 57, 455–464. [Google Scholar]
- Kalinganire, A.; Pinyopusarerk, K. Chukrasia: Biology, Cultivation and Utilisation; ACIAR Technical Reports No. 49; Australian Centre for International Agriculatural Reasearch: Canberra, Australia, 2000.
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestry Database: A Tree Reference and Selection Guide Version 4.0; World Agroforestry Centre: Nairobi, Kenya, 2009. [Google Scholar]
- Vlam, M.; Baker, P.J.; Bunyavejchewin, S.; Zuidema, P.A. Temperature and rainfall strongly drive temporal growth variation in Asian tropical forest trees. Oecologia 2014, 174, 1449–1461. [Google Scholar] [CrossRef]
- Baker, P.J.; Bunyavejchewin, S.; Oliver, C.D.; Ashton, P.S. Disturbance history and historical stand dynamics of a seasonal tropical forest in western Thailand. Ecol. Monogr. 2005, 75, 317–343. [Google Scholar] [CrossRef]
- Trouet, V.; Mukelabai, M.; Verheyden, A.; Beeckman, H. Cambial growth season of brevi-deciduous brachystegia spiciformis trees from South central Africa restricted to less than four months. PLoS ONE 2012, 7, e47364. [Google Scholar] [CrossRef]
- Worbes, M. Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. J. Ecol. 1999, 87, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Eckstein, D.; Bauch, J. Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwiss. Cent. 1969, 88, 230–250. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Briffa, K.R.; Jones, P.D. Basic chronology statistics and assessment. In Methods of Dendrochronology: Applications in the Environmental Sciences; Cook, E.R., Kairiuksti, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 137–152. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Cook, E.R.; Briffa, K.R.; Shiyatov, S.; Mazepa, V.; Jones, P.D. Data analysis. In Methods of Dendrochronology: Applications in the Environmental Sciences; Cook, E.R., Kairiuksti, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 97–162. [Google Scholar]
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press Inc.: London, UK, 1976. [Google Scholar]
- Torrence, C.; Gilbert, P.C. A Practical Guide to Wavelet Analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Pederson, N.; Cook, E.R.; Jacoby, G.C.; Peteet, D.M.; Griffin, K.L. The influence of winter temperatures on the annual radial growth of six northern range margin tree species. Dendrochronologia 2004, 22, 7–29. [Google Scholar] [CrossRef]
- Buckley, B.M.; Palakit, K.; Duangsathaporn, K.; Sanguantham, P.; Prasomsin, P. Decadal scale droughts over northwestern Thailand over the past 448 years: Links to the tropical Pacific and Indian Ocean sectors. Clim. Dyn. 2007, 29, 63–71. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A. Time lag and negative responses of forest greenness and tree growth to warming over circumboreal forests. Glob. Chang. Biol. 2018, 24, 4225–4237. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Matsuura, Y.; Osawa, A.; Sato, H.; Fujinuma, J.; Maximov, T. Tree-ring analysis and modeling approaches yield contrary response of circumboreal forest productivity to climate change. Glob. Chang. Biol. 2017, 23, 5179–5188. [Google Scholar] [CrossRef]
- Sideridis, G.D.; Simos, P. What is the actual correlation between expressive and receptive measures of vocabulary? Approximating the sampling distribution of the correlation coefficient using the bootstrapping method. Int. J. Educ. Psychol. Assess. 2010, 5, 117–133. [Google Scholar]
- Grissino-Mayer, H.D.; Romme, W.H.; Floyd, M.L.; Hanna, D.D. Climatic and human influences on fire regimes of the southern San Juan Mountains, Colorado, USA. Ecology 2004, 85, 1708–1724. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. Treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Raihan, F.; Li, G.; Harrison, S.P. Detection of recent changes in climate using meteorological data from South-eastern Bangladesh. J. Climatol. Weather Forecast. 2015, 3, 137. [Google Scholar]
- Stahle, D.W.; Mushove, P.T.; Cleaverland, M.K.; Haynes, G. Management implications of annual growth rings in Ptreocarpus angolensis from Zimbabwe. For. Ecol. Manag. 1999, 124, 217–229. [Google Scholar] [CrossRef]
- Bräuning, A.; Volland-Voigt, F.; Burchardt, I.; Ganzhi, O.; Nauss, T.; Peters, T. Climatic control of radial growth of Cedrela montana in a humid mountain rainforest in southern Ecuador. Erdkunde 2009, 63, 337–345. [Google Scholar] [CrossRef]
- Chen, F.; He, Q.; Bakytbek, E.; Yu, S.; Zhang, R. Climatic signals in tree rings of Juniperus turkestanica in the Gulcha River Basin (Kyrgyzstan), reveals the recent wetting trend of high Asia. Dendrobiology 2015, 74, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.X.; Davies, S.J.; Ashton, P.S.; Bunyavejchewin, S.; Supardi, M.N.N.; Kassim, A.R.; Tan, S.; Moorcroft, P.R. Variability in solar radiation and temperature explains observed patterns and trends in tree growth rates across four tropical forests. Proc. R. Soc. B Biol. Sci. 2012, 279, 3923–3931. [Google Scholar] [CrossRef] [Green Version]
- Galbraith, D.; Levy, P.E.; Sitch, S.; Huntingford, C.; Cox, P.; Williams, M.; Meir, P. Multiple mechanisms of Amazonian forest biomass losses in three dynamic global vegetation models under climate change. New Phytol. 2010, 187, 647–665. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.A. Detecting tropical forests’ responses to global climatic and atmospheric change: Current challenges and a way forward. Biotropica 2007, 39, 4–19. [Google Scholar] [CrossRef]
- Meir, P.; Metcalfe, D.B.; Costa, A.C.L.; Fisher, R.A. The fate of assimilated carbon during drought: Impacts on respiration in Amazon rainforests. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 1849–1855. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. The CO2 dependence of photosynthesis, plant growth responses to elevated atmospheric CO2 concentrations and their interaction with soil nutrient status. I. General principles and forest ecosystems. Funct. Ecol. 1996, 10, 4–32. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. Effects of rising temperatures and CO2 on the physiology of tropical forest trees. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 1811–1817. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bruhn, D.; Hurry, V.M.; Tjoelker, M.G. Evans Review No. 2—The hot and the cold: Unravelling the variable response of plant respiration to temperature. Funct. Plant Biol. 2005, 32, 87–105. [Google Scholar] [CrossRef]
- Will, R.E.; Wilson, S.M.; Zou, C.B.; Hennessey, T.C. Increased vapor pressure deficit due to higher temperature leads to greater transpiration and faster mortality during drought for tree seedlings common to the forest-grassland ecotone. New Phytol. 2013, 200, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Fichtler, E.; Trouet, V.; Beeckman, H.; Coppin, P.; Worbes, M. Climatic signals in tree rings of Burkea africana and Pterocarpus angolensis from semiarid forests in Namibia. Trees-Struct. Funct. 2004, 18, 442–451. [Google Scholar] [CrossRef]
- Ram, S.; Borgaonkar, H.P.; Sikder, A.B. Tree-ring analysis of teak (Tectona grandis L.F.) in central India and its relationship with rainfall and moisture index. J. Earth Syst. Sci. 2008, 117, 637–645. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Cook, E.R.; Wilson, R.J.; Allan, R.; Mann, M.E. On the variability of ENSO over the past six centuries. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef]
- Macias, M.; Andreu, L.; Bosch, O.; Camarero, J.J.; Gutiérrez, E. Increasing aridity is enhancing silver fir (Abies alba Mill.) water stress in its south-western distribution limit. Clim. Chang. 2006, 79, 289–313. [Google Scholar] [CrossRef]
- Andreu, L.; Gutiérrez, E.; Macias, M.; Ribas, M.; Bosch, O.; Camarero, J.J. Climate increases regional tree-growth variability in Iberian pine forests. Glob. Chang. Biol. 2007, 13, 804–815. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Values |
|---|---|
| Time span | 1895–2015 |
| Mean length of series (year) | 64 |
| Trees/cores | 33/52 |
| Mean ring width (mm) ± SD | 3.85 ± 1.33 |
| First order autocorrelation (AC1) | 0.33 |
| Mean GLK * | 0.68 |
| rbar.eff *δ | 0.28 |
| Series inter-correlation (SIC) δ | 0.45 |
| Mean expressed population signal (EPS) | 0.90 |
| Mean sensitivity (MS) | 0.69 |
| Signal-to-noise ratio (SNR) | 9.13 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.; Islam, M.; Wernicke, J.; Bräuning, A. Changes in Sensitivity of Tree-Ring Widths to Climate in a Tropical Moist Forest Tree in Bangladesh. Forests 2018, 9, 761. https://doi.org/10.3390/f9120761
Rahman M, Islam M, Wernicke J, Bräuning A. Changes in Sensitivity of Tree-Ring Widths to Climate in a Tropical Moist Forest Tree in Bangladesh. Forests. 2018; 9(12):761. https://doi.org/10.3390/f9120761
Chicago/Turabian StyleRahman, Mizanur, Mahmuda Islam, Jakob Wernicke, and Achim Bräuning. 2018. "Changes in Sensitivity of Tree-Ring Widths to Climate in a Tropical Moist Forest Tree in Bangladesh" Forests 9, no. 12: 761. https://doi.org/10.3390/f9120761
APA StyleRahman, M., Islam, M., Wernicke, J., & Bräuning, A. (2018). Changes in Sensitivity of Tree-Ring Widths to Climate in a Tropical Moist Forest Tree in Bangladesh. Forests, 9(12), 761. https://doi.org/10.3390/f9120761