Eddy Covariance vs. Biometric Based Estimates of Net Primary Productivity of Pedunculate Oak (Quercus robur L.) Forest in Croatia during Ten Years


 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
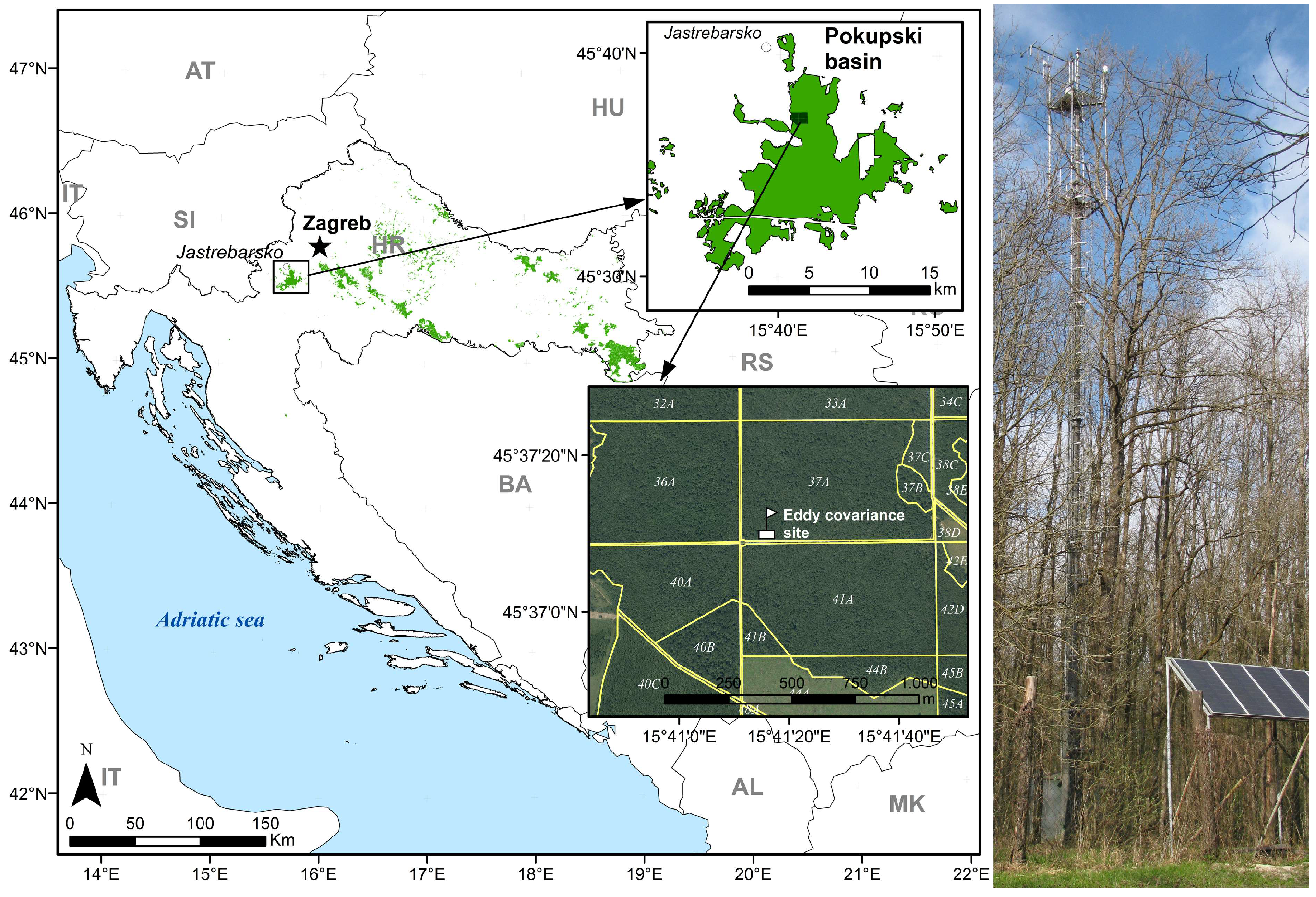
2.1. Research Area and Site Measurement Design
2.2. Eddy Covariance and Meteorological Measurements
2.2.1. EC and Meteorological Instrumentation
2.2.2. EC Data Processing
2.3. NEE Flux Partitioning and the Estimation of NPPEC
2.4. Assessment of Flux Footprint
2.5. Biometric Measurements and Estimation of NPPBM
2.5.1. NPP of Total Woody Biomass, Leaves and Fruits
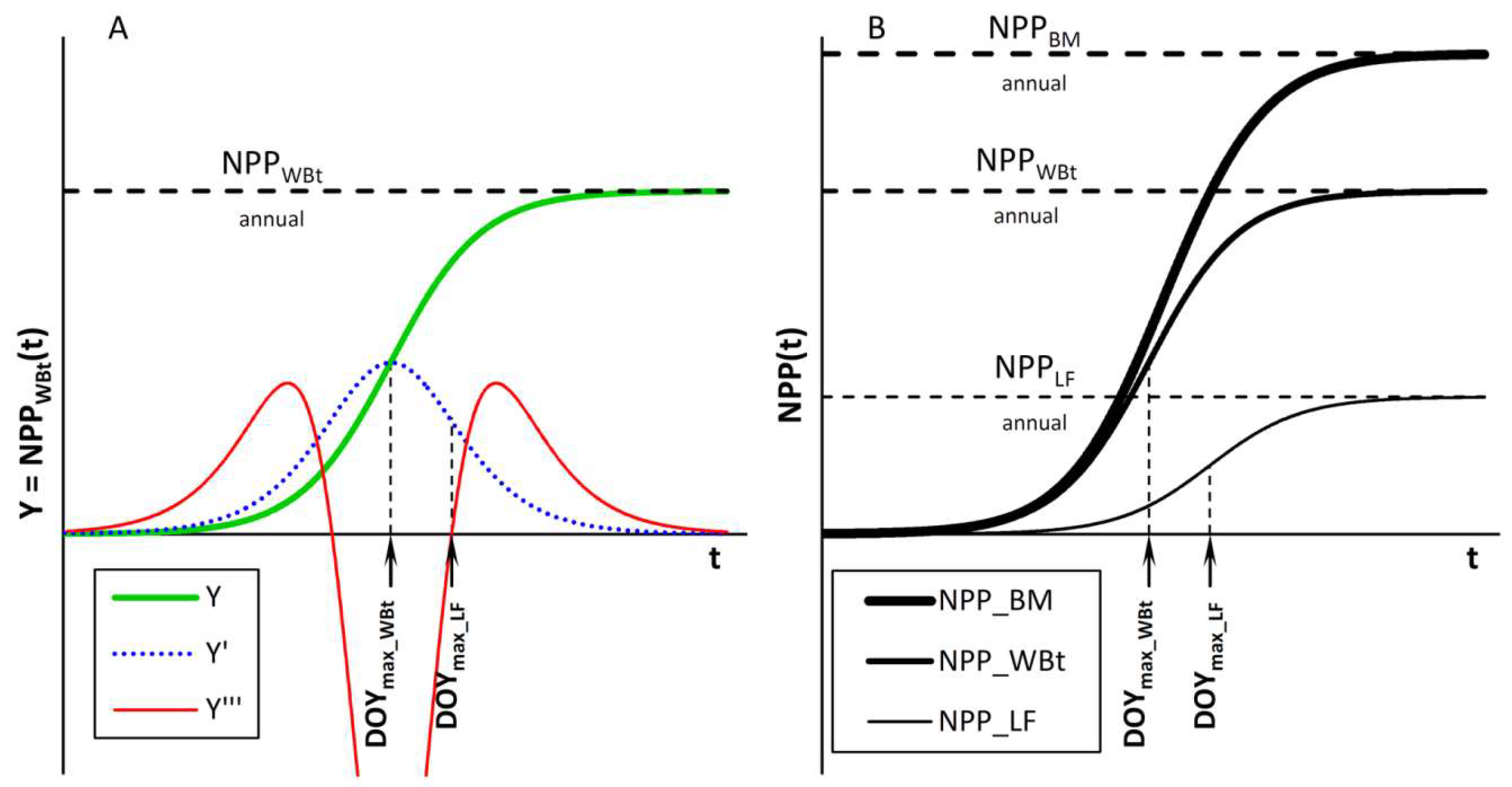
2.5.2. Modelling Seasonal Dynamics of NPPBM
3. Results
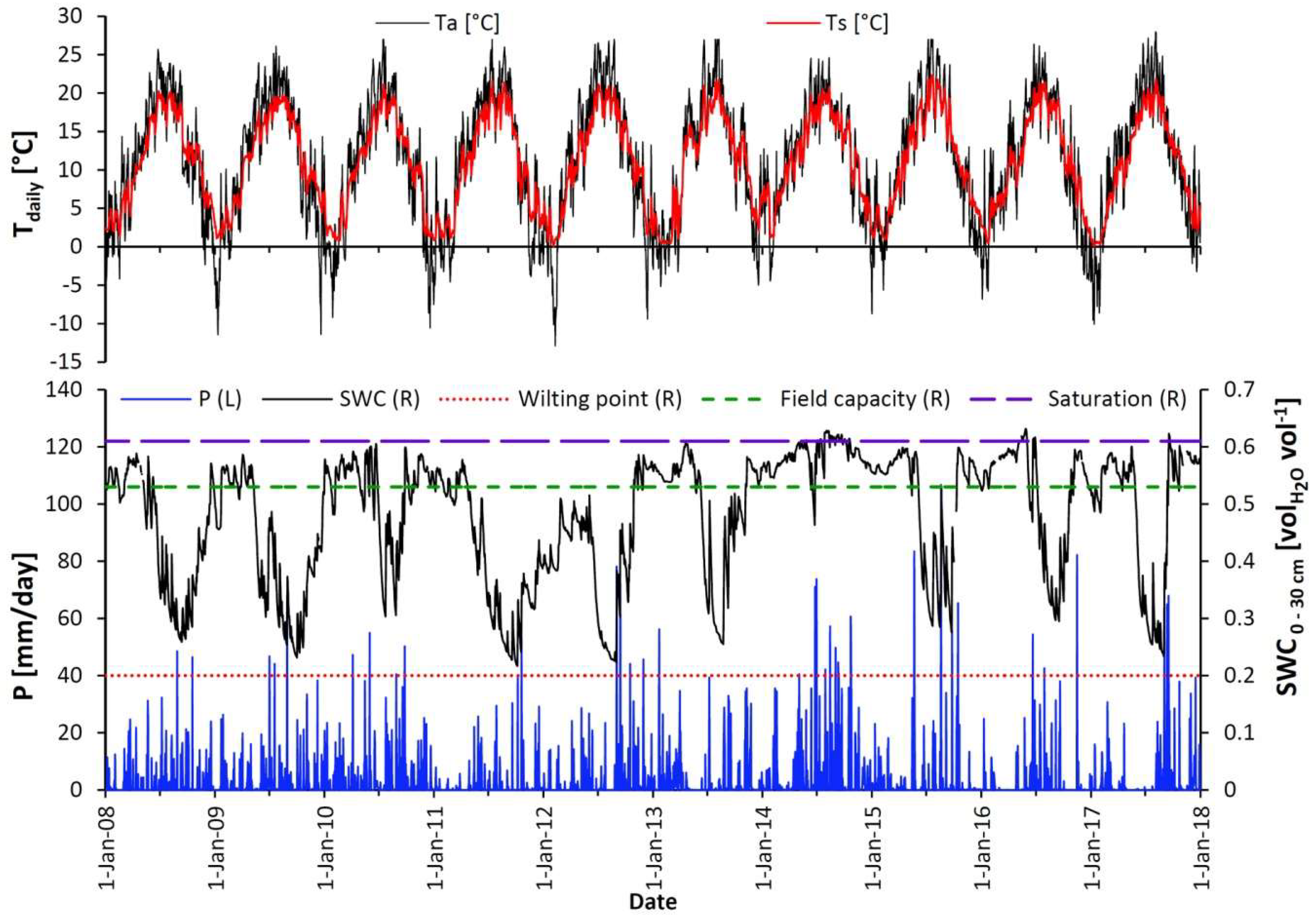
3.1. Meteorological Conditions during the Period 2008–2017
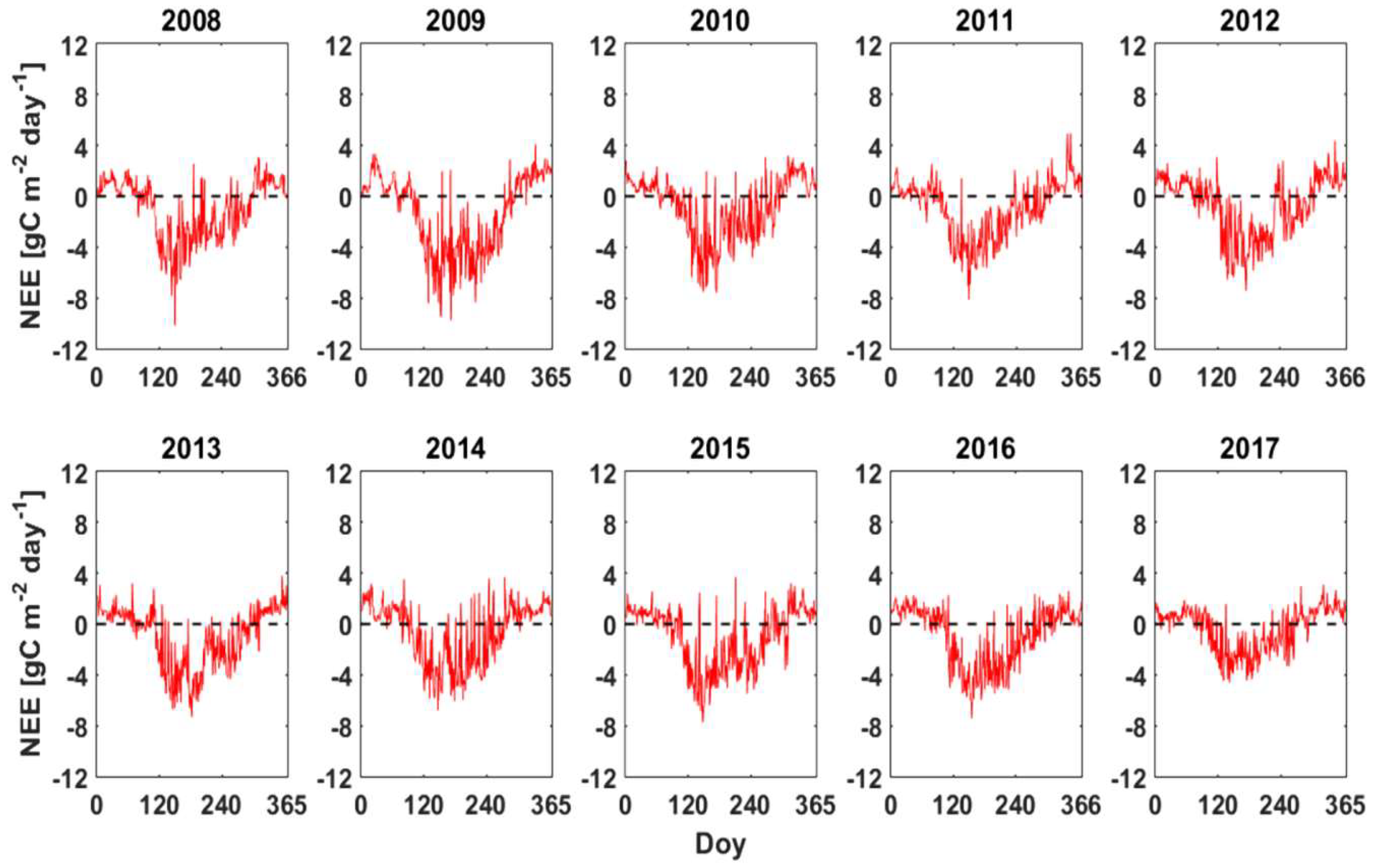
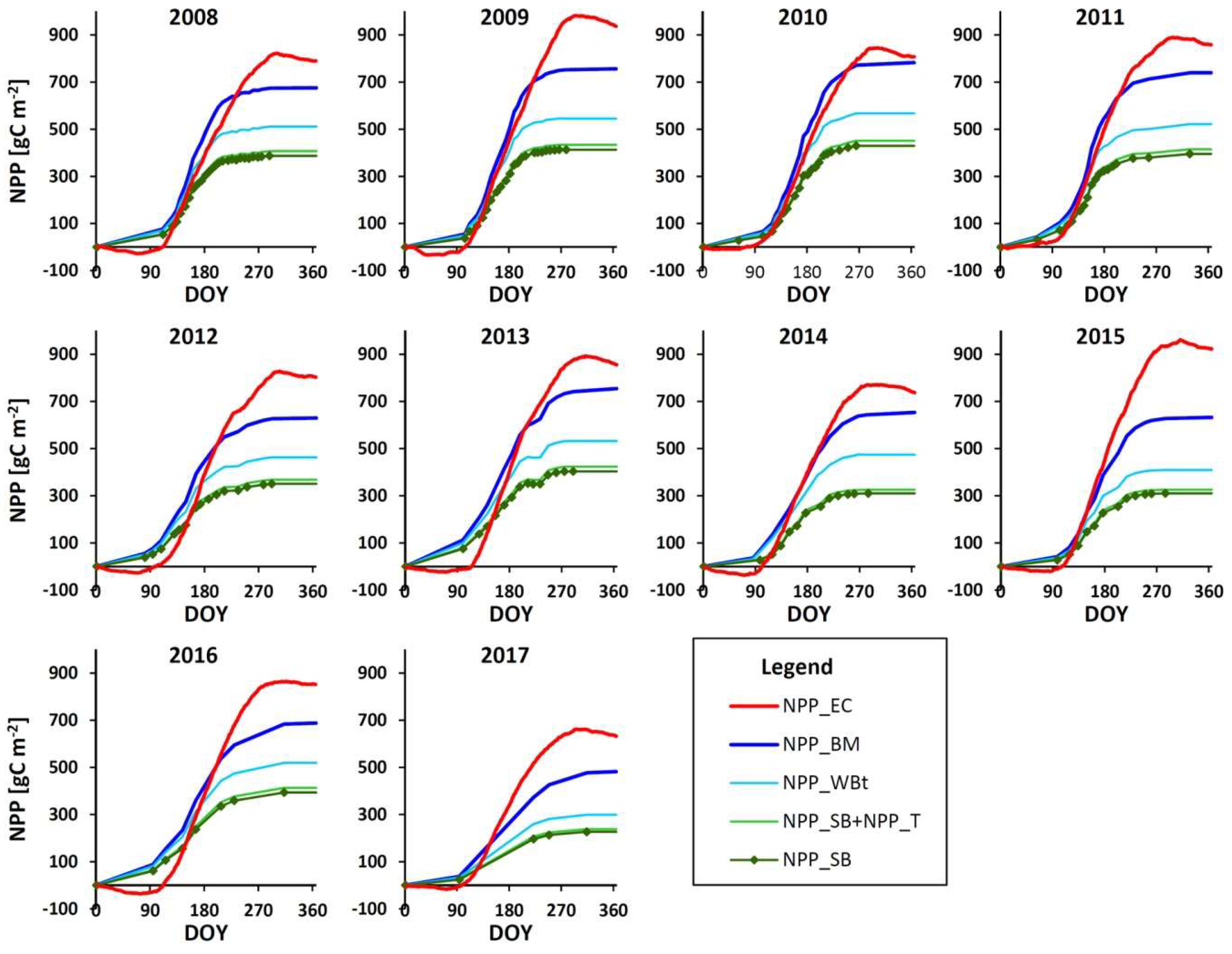
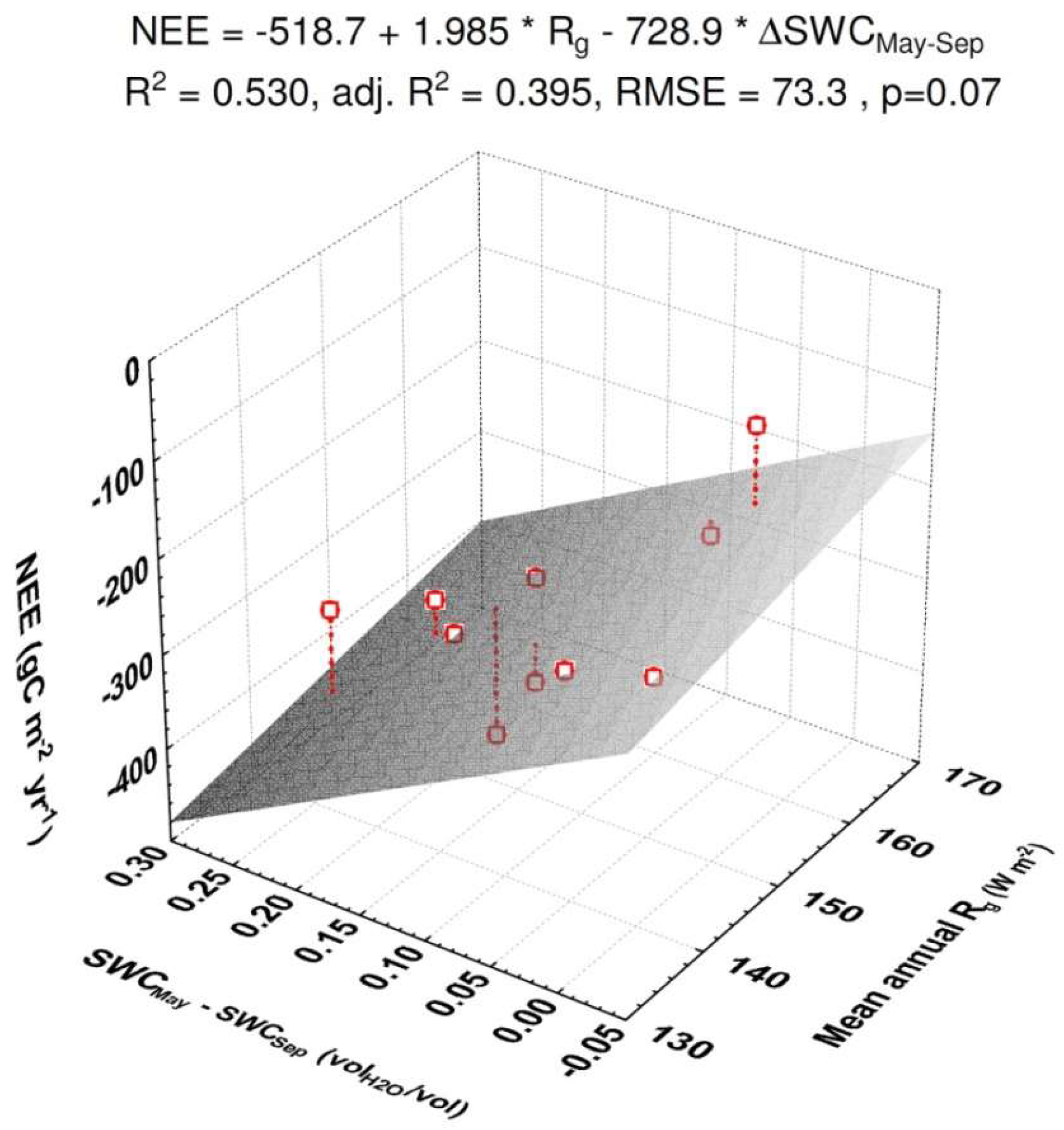
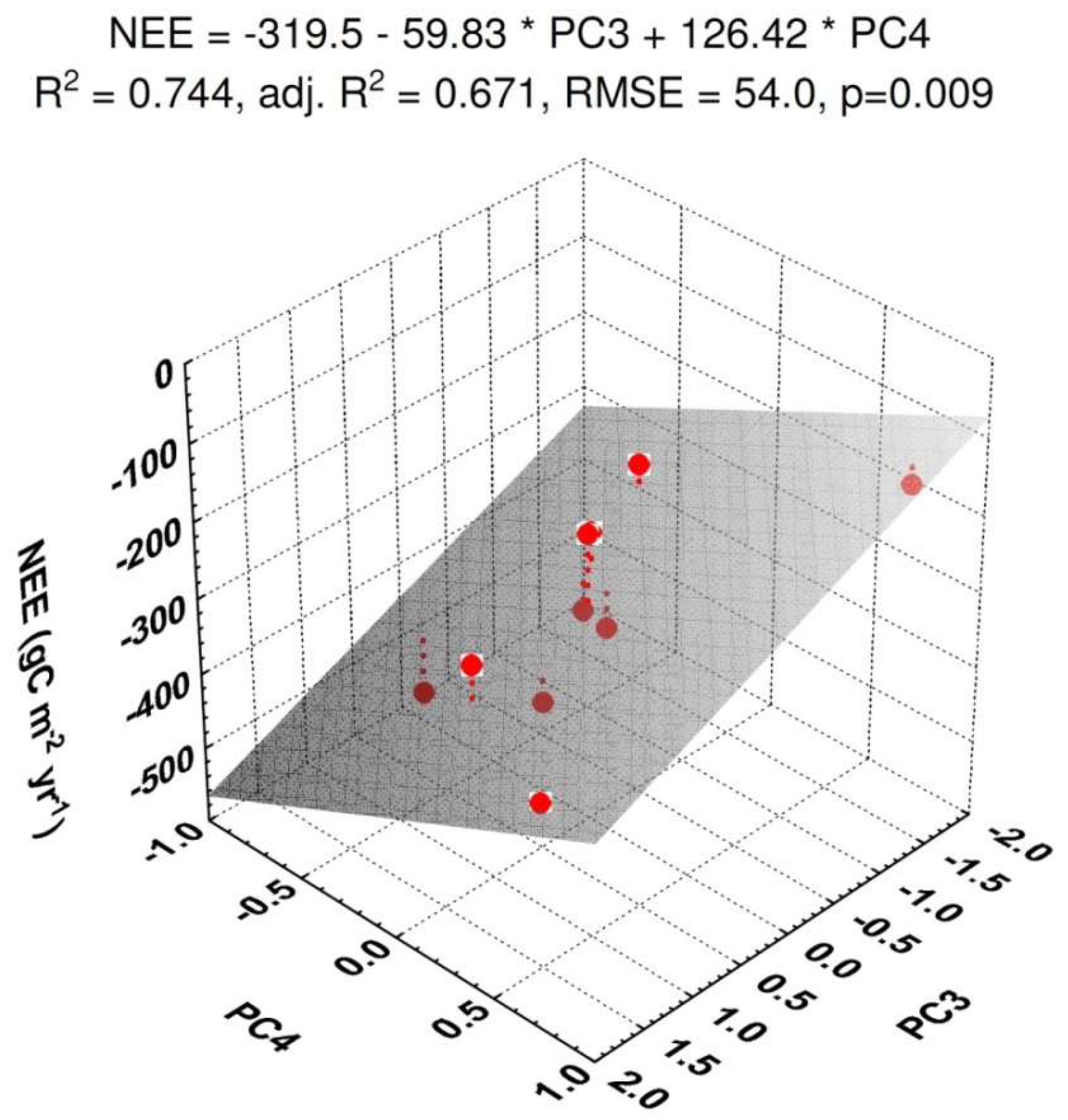
3.2. NEE
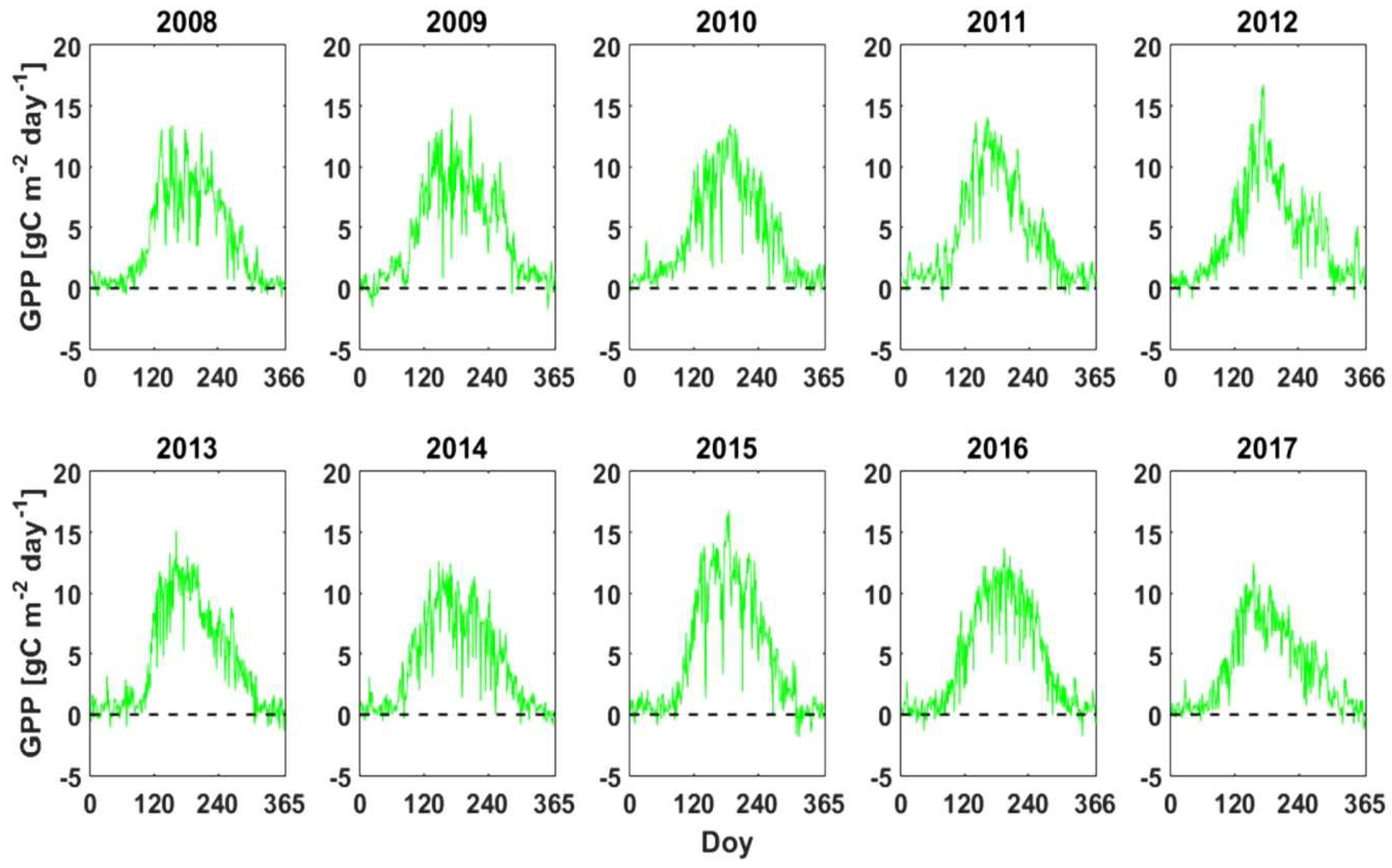
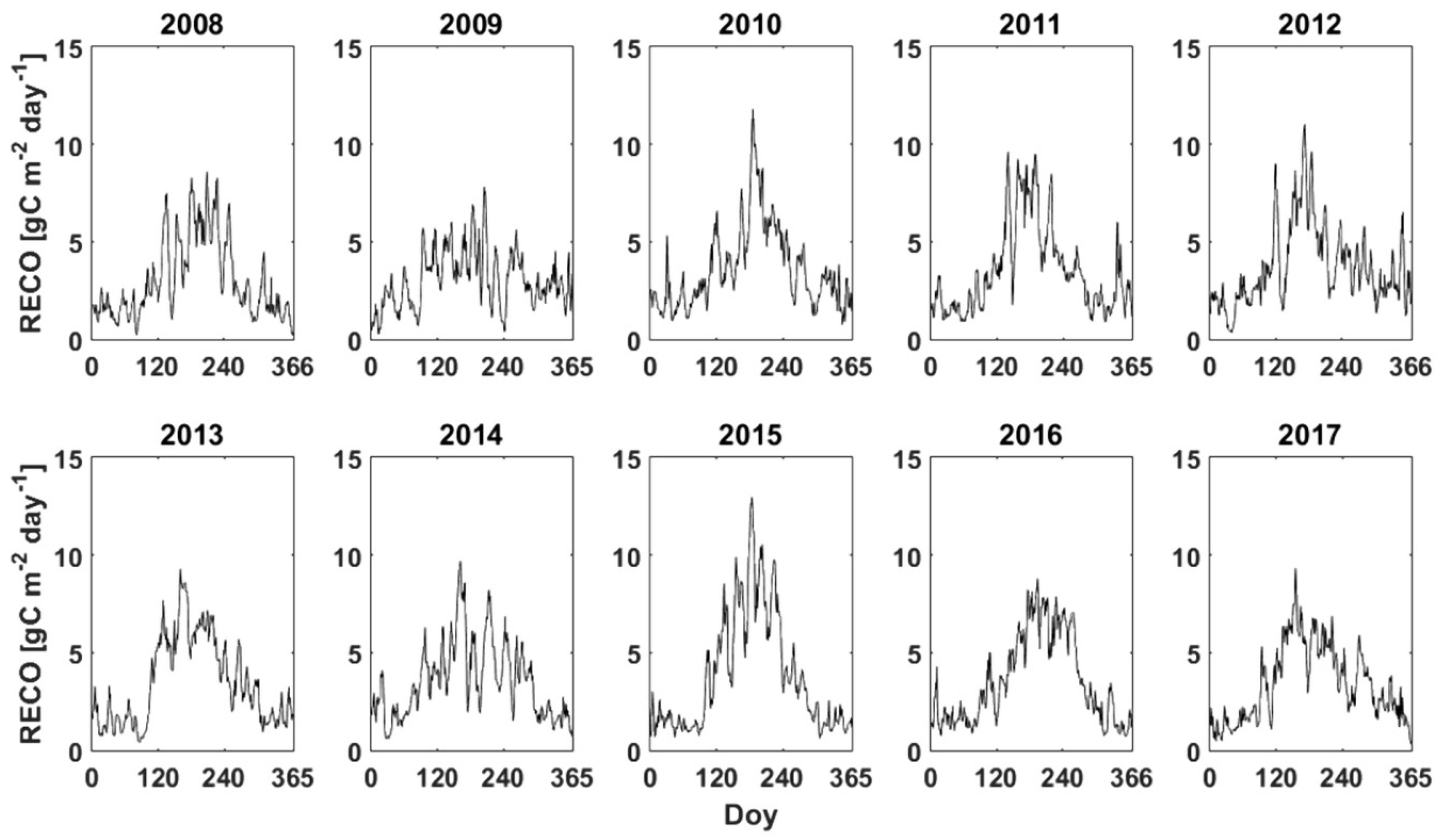
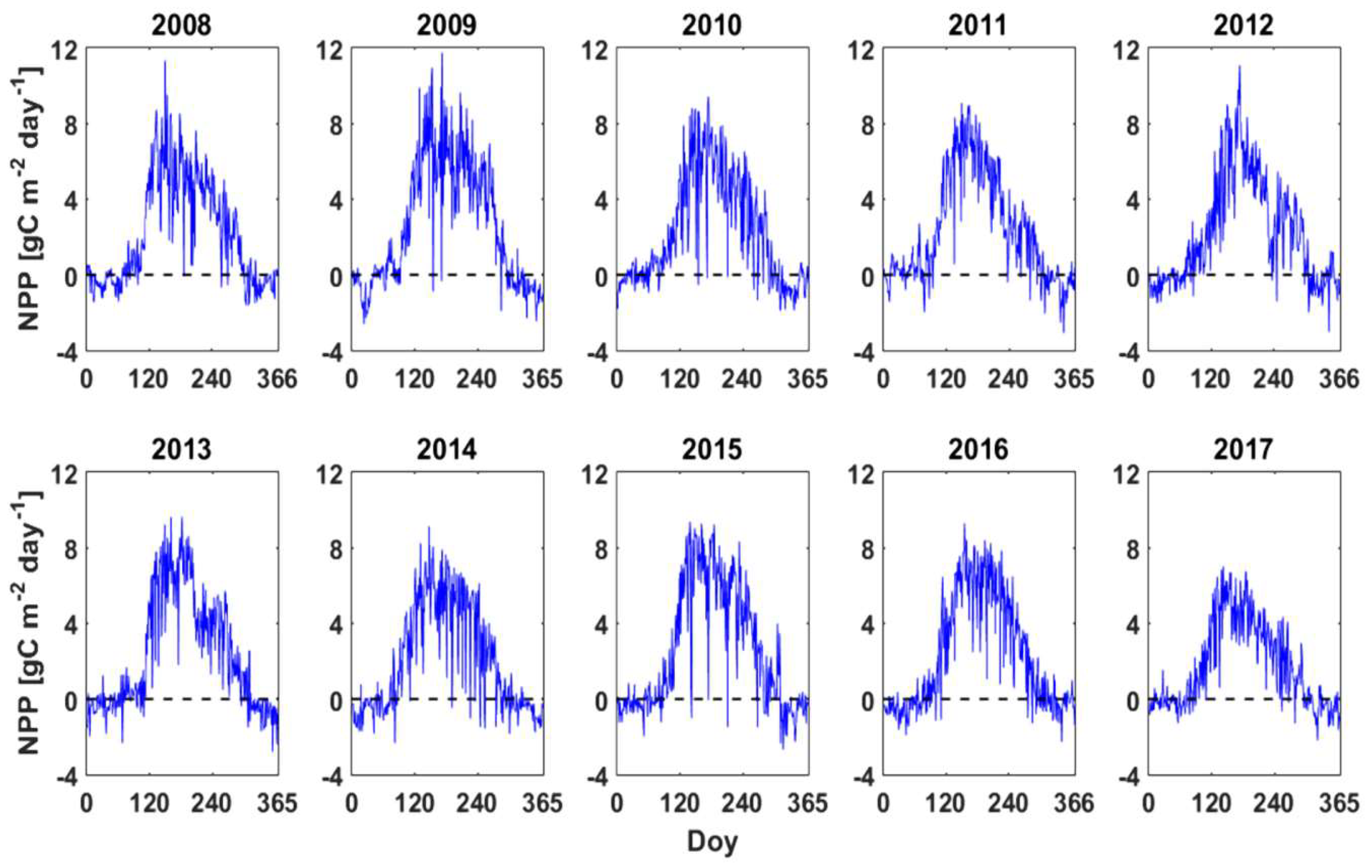
3.3. GPP, RECO and NPPEC
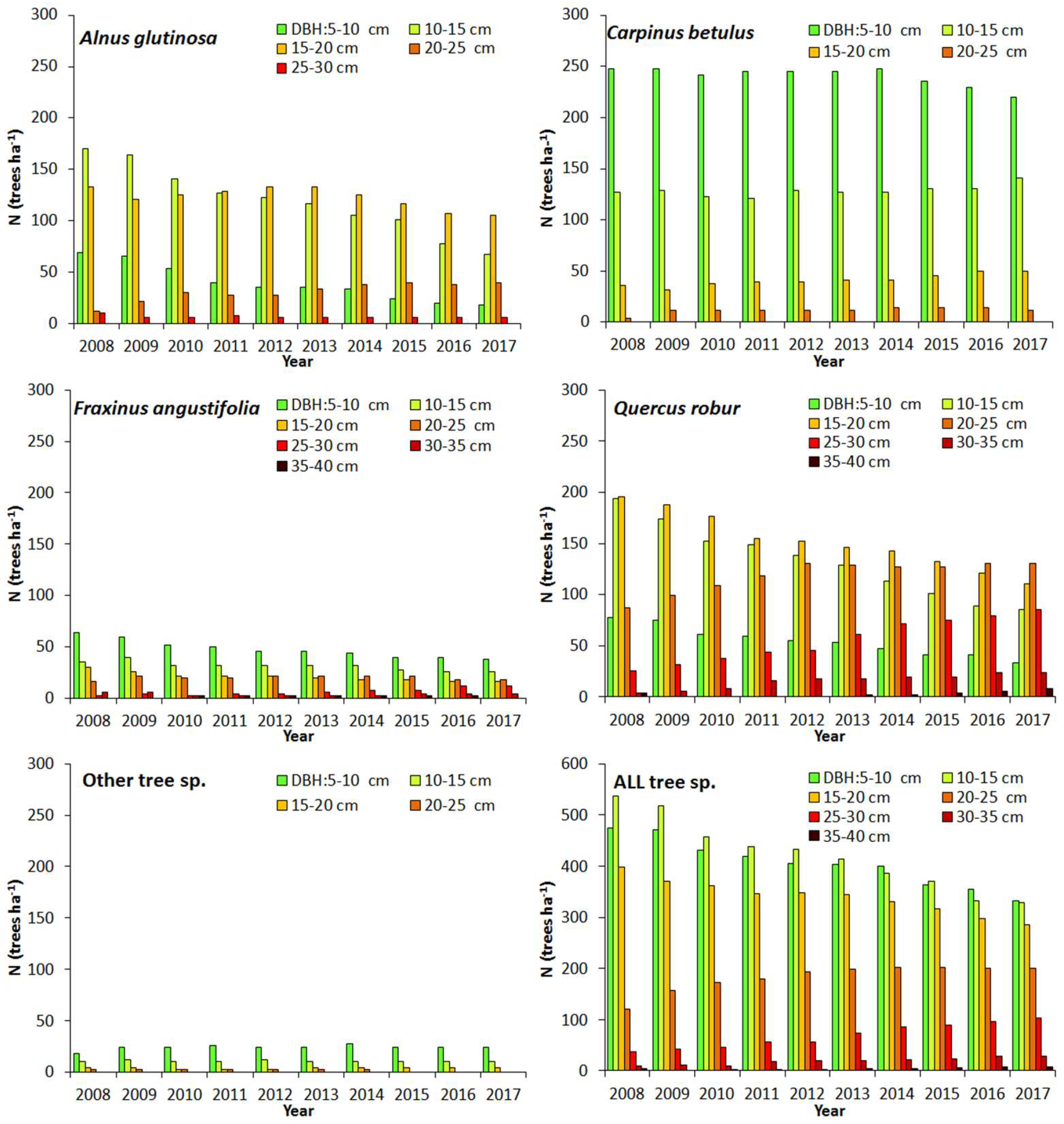
3.4. Stand Characteristics and NPPBM
3.4.1. Stand Development
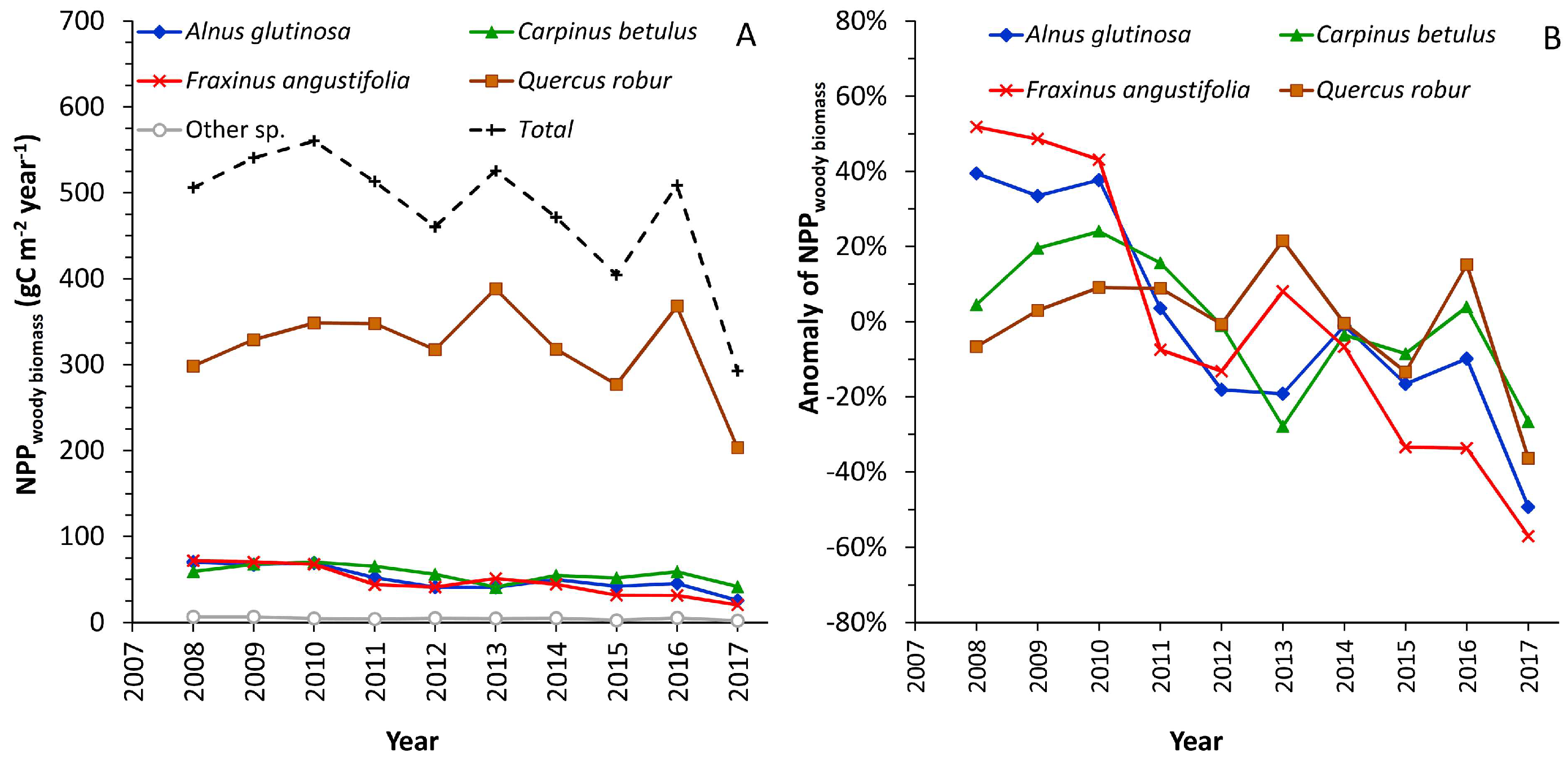
3.4.2. NPPWBt by Tree Species
3.4.3. NPPBM
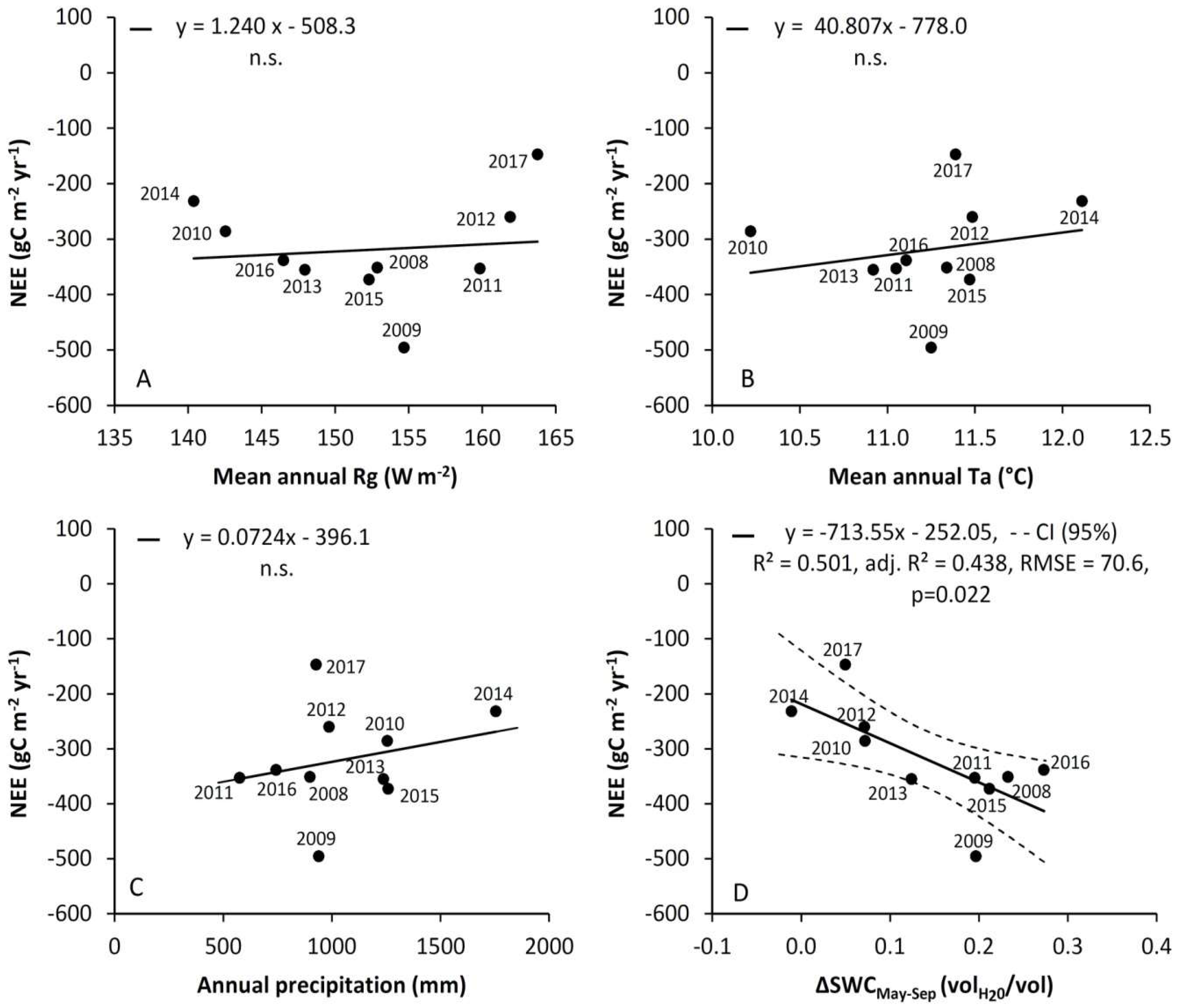
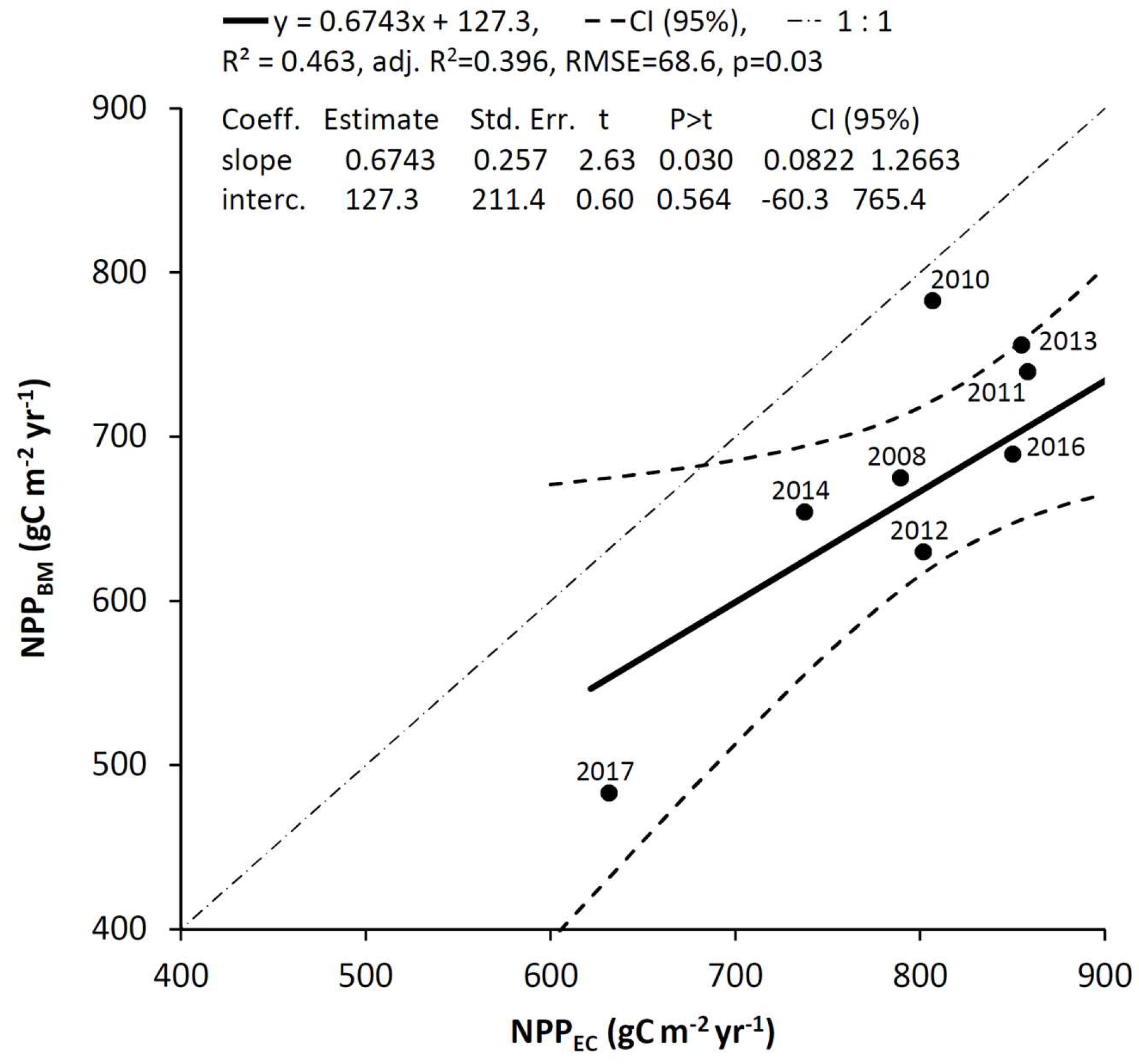
3.5. Comparison of NPP Estimates from Eddy Covariance and Biometric Measurements
4. Discussion
4.1. Variability of CO2 Fluxes from EC
4.2. Dynamic of NPP Estimated with Biometric Method
4.3. Comparison of Biometric and Eddy Covariance NPP Estimate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Data Coverage before Processing [%] | Data Coverage after Quality Control [%] | Data Coverage after u* Filtering [%] |
|---|---|---|---|
| 2008 | 95.5 | 46.6 | 39.4 |
| 2009 | 88.2 | 46.8 | 42.1 |
| 2010 | 91.9 | 47.0 | 45.4 |
| 2011 | 94.3 | 56.5 | 47.7 |
| 2012 | 83.3 | 49.8 | 42.7 |
| 2013 | 92.8 | 53.0 | 48.4 |
| 2014 | 95.7 | 51.4 | 47.3 |
| 2015 | 96.0 | 52.9 | 43.2 |
| 2016 | 83.8 | 45.1 | 41.6 |
| 2017 | 79.0 | 45.0 | 38.0 |
| Species ** | Parameters * | |||
|---|---|---|---|---|
| Alnus glutinosa | 10.31 | 0.37 | 0.59 | 0.1385 |
| Carpinus betulus | 6.60 | 0.59 | −2.31 | 0.2863 |
| Fraxinus angustifolia | 6.25 | 0.58 | −3.89 | 0.3275 |
| Quercus robur | 3.86 | 0.63 | −1.61 | 0.2496 |
| Species *** | Parameter | Source | ||
|---|---|---|---|---|
| b0 | b1 | b2 | ||
| Alnus glutinosa | 4.23243∙10−5 | 2.002354 | 1.001300 | [40] |
| Carpinus betulus | 2.96400∙10−5 | 2.022705 | 1.102119 | [41] |
| Fraxinus angustifolia | 3.95282∙10−5 | 1.974875 | 1.001444 | [42] |
| Quercus robur | 4.96820∙10−5 | 2.048384 | 0.892124 | [41] |
| Parameter | Year | Estimate | Std. Err. | t | p > |t| | CI95lower | CI95uper |
|---|---|---|---|---|---|---|---|
| DOYmax_WBt | 2008 | 151.6 | 0.5 | 317.43 | 0.000 | 150.7 | 152.6 |
| 2009 | 154.1 | 0.7 | 227.61 | 0.000 | 152.7 | 155.5 | |
| 2010 | 158.3 | 1.1 | 138.99 | 0.000 | 155.9 | 160.7 | |
| 2011 | 144.7 | 1.6 | 91.91 | 0.000 | 141.3 | 148.0 | |
| 2012 | 143.2 | 1.3 | 108.31 | 0.000 | 140.3 | 146.1 | |
| 2013 | 152.0 | 2.0 | 75.94 | 0.000 | 147.6 | 156.4 | |
| 2014 | 156.3 | 1.2 | 126.73 | 0.000 | 153.5 | 159.0 | |
| 2015 | 155.3 | 1.7 | 93.04 | 0.000 | 151.5 | 159.1 | |
| 2016 | 154.4 | 2.7 | 57.64 | 0.000 | 146.9 | 161.8 | |
| 2017 | 163.5 | 0.3 | 590.49 | 0.001 | 160.0 | 167.0 | |
| k | 2008 | 0.0476 | 0.0011 | 42.77 | 0.000 | 0.0453 | 0.0499 |
| 2009 | 0.0426 | 0.0012 | 36.81 | 0.000 | 0.0402 | 0.0450 | |
| 2010 | 0.0397 | 0.0015 | 26.67 | 0.000 | 0.0365 | 0.0428 | |
| 2011 | 0.0431 | 0.0027 | 15.83 | 0.000 | 0.0373 | 0.0490 | |
| 2012 | 0.0356 | 0.0015 | 23.36 | 0.000 | 0.0323 | 0.0389 | |
| 2013 | 0.0287 | 0.0017 | 16.5 | 0.000 | 0.0249 | 0.0325 | |
| 2014 | 0.0322 | 0.0010 | 31.4 | 0.000 | 0.0299 | 0.0345 | |
| 2015 | 0.0404 | 0.0026 | 15.63 | 0.000 | 0.0346 | 0.0463 | |
| 2016 | 0.0293 | 0.0019 | 15.24 | 0.000 | 0.0240 | 0.0346 | |
| 2017 | 0.0307 | 0.0001 | 248.18 | 0.003 | 0.0291 | 0.0322 | |
| NPPWBt annual | 2008 | 504.9 | 2.3 | 216.26 | 0.000 | 500.1 | 509.8 |
| 2009 | 550.9 | 3.7 | 150.55 | 0.000 | 543.3 | 558.6 | |
| 2010 | 571.5 | 7.5 | 76.41 | 0.000 | 555.7 | 587.3 | |
| 2011 | 513.1 | 9.7 | 52.89 | 0.000 | 492.3 | 533.9 | |
| 2012 | 457.0 | 5.3 | 85.69 | 0.000 | 445.3 | 468.6 | |
| 2013 | 540.4 | 9.1 | 59.64 | 0.000 | 520.5 | 560.4 | |
| 2014 | 486.5 | 4.8 | 102.17 | 0.000 | 475.9 | 497.1 | |
| 2015 | 409.9 | 6.3 | 65.16 | 0.000 | 395.7 | 424.1 | |
| 2016 | 526.9 | 11.6 | 45.46 | 0.000 | 494.8 | 559.1 | |
| 2017 | 302.0 | 0.3 | 900.4 | 0.001 | 297.7 | 306.2 |
| Season | Annual | Season | Annual | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Year | DJF | MAM | JJA | SON | Values | DJF | MAM | JJA | SON | Values |
| Ta (°C) | Ts (°C) | |||||||||
| 2008 | 2.55 | 11.30 | 20.08 | 10.90 | 11.29 | 3.65 | 9.69 | 18.09 | 12.25 | 11.12 |
| 2009 | 0.87 | 12.45 | 20.02 | 11.49 | 11.28 | 3.70 | 10.53 | 17.65 | 12.56 | 11.24 |
| 2010 | 0.75 | 10.91 | 20.10 | 10.13 | 10.27 | 3.80 | 9.59 | 17.69 | 11.42 | 10.31 |
| 2011 | 0.24 | 11.65 | 20.47 | 10.14 | 11.04 | 2.20 | 9.82 | 18.12 | 11.23 | 10.63 |
| 2012 | 0.63 | 12.16 | 21.81 | 11.89 | 11.49 | 3.03 | 10.31 | 18.58 | 12.73 | 11.06 |
| 2013 | 0.73 | 10.36 | 20.62 | 11.26 | 10.92 | 2.01 | 9.63 | 17.89 | 12.07 | 10.52 |
| 2014 | 3.85 | 12.42 | 19.30 | 12.20 | 12.10 | 4.86 | 11.19 | 17.93 | 12.90 | 11.82 |
| 2015 | 2.74 | 11.67 | 21.27 | 10.75 | 11.54 | 3.43 | 10.29 | 18.78 | 11.73 | 11.11 |
| 2016 | 2.91 | 11.36 | 20.19 | 10.82 | 11.11 | 3.78 | 9.99 | 18.51 | 11.96 | 11.23 |
| 2017 | −0.40 | 12.57 | 21.62 | 10.65 | 11.57 | 2.06 | 10.48 | 18.49 | 11.98 | 10.83 |
| Rg (Wm−2) | P (mm) | |||||||||
| 2008 | 62 | 189 | 256 | 107 | 153 | 106 | 279 | 245 | 223 | 899 |
| 2009 | 48 | 193 | 262 | 113 | 155 | 228 | 164 | 345 | 204 | 940 |
| 2010 | 41 | 174 | 257 | 93 | 143 | 292 | 306 | 305 | 382 | 1255 |
| 2011 | 49 | 209 | 265 | 112 | 160 | 100 | 94 | 193 | 166 | 576 |
| 2012 | 63 | 205 | 281 | 96 | 162 | 170 | 184 | 137 | 526 | 987 |
| 2013 | 43 | 178 | 270 | 98 | 148 | 310 | 207 | 197 | 355 | 1024 |
| 2014 | 45 | 181 | 246 | 86 | 140 | 247 | 325 | 527 | 603 | 1755 |
| 2015 | 51 | 194 | 268 | 96 | 152 | 215 | 230 | 288 | 325 | 991 |
| 2016 | 48 | 168 | 261 | 106 | 146 | 102 | 95 | 338 | 208 | 744 |
| 2017 | 57 | 209 | 286 | 98 | 164 | 130 | 85 | 99 | 501 | 927 |
| NEE | GPP | RECO | Rg | Ta | Ts | P | SWC | ΔSWCMay–Sep | SGS | EGS | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NEE | 1 | ||||||||||
| GPP | −0.578 | 1 | |||||||||
| RECO | 0.327 | 0.582 | 1 | ||||||||
| Rg | 0.107 | −0.184 | −0.107 | 1 | |||||||
| Ta | 0.209 | −0.213 | −0.038 | 0.105 | 1 | ||||||
| Ts | −0.030 | −0.108 | −0.155 | −0.176 | 0.902 * | 1 | |||||
| P | 0.255 | −0.004 | 0.250 | −0.644 * | 0.328 | 0.398 | 1 | ||||
| SWC | 0.421 | −0.118 | 0.281 | −0.778 * | 0.191 | 0.272 | 0.776 * | 1 | |||
| ΔSWCMay–Sep | −0.707 * | 0.403 | −0.240 | 0.089 | −0.219 | −0.096 | −0.645 * | −0.422 | 1 | ||
| SGS | −0.021 | 0.221 | 0.238 | −0.075 | 0.054 | 0.019 | 0.160 | 0.025 | −0.228 | 1 | |
| EGS | −0.165 | −0.189 | −0.382 | 0.672 * | −0.458 | −0.550 | −0.749 * | −0.834 * | 0.247 | 0.131 | 1 |
| (a) Eigenvalues of the Correlation Matrix | |||||||
| Principal Component (PC) | Eigenvalue | Difference | Proportion of Variance | Cumulative Proportion | |||
| PC1 | 3.0422 | 1.3715 | 0.507 | 0.507 | |||
| PC2 | 1.6706 | 0.7073 | 0.278 | 0.786 | |||
| PC3 | 0.9633 | 0.7660 | 0.161 | 0.946 | |||
| PC4 | 0.1973 | 0.0945 | 0.033 | 0.979 | |||
| PC5 | 0.1028 | 0.0791 | 0.017 | 0.996 | |||
| PC6 | 0.0237 | - | 0.004 | 1.000 | |||
| (b) Relation between the Physical Driver Variables and the Principal Components | |||||||
| Principal Components (Eigenvectors) | |||||||
| Variable | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | Unexplained |
| P | 0.5313 | −0.1376 | −0.1502 | −0.4509 | 0.6785 | 0.1121 | 0 |
| Ta | 0.3025 | 0.6464 | 0.0065 | 0.2386 | 0.1597 | −0.6389 | 0 |
| Ts | 0.3513 | 0.5572 | 0.2789 | −0.2349 | −0.3487 | 0.5581 | 0 |
| Rg | −0.3844 | 0.4151 | −0.4819 | 0.2968 | 0.4023 | 0.4446 | 0 |
| SWC | 0.4925 | −0.2747 | 0.1317 | 0.7710 | 0.0573 | 0.2587 | 0 |
| ΔSWCMay–Sep | −0.3352 | 0.0694 | 0.8062 | 0.0468 | 0.4769 | 0.0565 | 0 |
| (c) Correlation between the NEE and the Principal Components (PCs) | |||||||
| NEE | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
| NEE | 1 | ||||||
| PC1 | 0.3391 | 1 | |||||
| PC2 | −0.0284 | 0 | 1 | ||||
| PC3 | −0.6233 # | 0 | 0 | 1 | |||
| PC4 | 0.5962 # | 0 | 0 | 0 | 1 | ||
| PC5 | −0.1664 | 0 | 0 | 0 | 0 | 1 | |
| PC6 | −0.0355 | 0 | 0 | 0 | 0 | 0 | 1 |
| Species | DBH | Year (at the End of the Season) | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (cm) | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | |||||||||||||
| N—stand density (trees ha−1) | ||||||||||||||||||||||||
| Quercus robur | <10 | 119 | ±25 | 104 | ±24 | 100 | ±23 | 85 | ±22 | 81 | ±21 | 76 | ±20 | 74 | ±20 | 67 | ±18 | 61 | ±17 | 59 | ±17 | 52 | ±15 | |
| ≥10 | 502 | ±61 | 516 | ±63 | 505 | ±62 | 488 | ±61 | 486 | ±61 | 491 | ±61 | 491 | ±61 | 480 | ±57 | 464 | ±53 | 453 | ±51 | 447 | ±48 | ||
| All | 620 | ±64 | 620 | ±64 | 605 | ±64 | 574 | ±60 | 568 | ±59 | 566 | ±59 | 564 | ±59 | 548 | ±56 | 525 | ±51 | 512 | ±50 | 499 | ±47 | ||
| Fraxinus angustifolia | <10 | 91 | ±34 | 89 | ±33 | 81 | ±30 | 73 | ±28 | 68 | ±26 | 64 | ±24 | 64 | ±24 | 60 | ±22 | 56 | ±20 | 56 | ±20 | 54 | ±19 | |
| ≥10 | 90 | ±25 | 92 | ±26 | 100 | ±29 | 82 | ±25 | 84 | ±24 | 86 | ±25 | 86 | ±25 | 86 | ±25 | 84 | ±25 | 80 | ±24 | 80 | ±24 | ||
| All | 181 | ±54 | 181 | ±54 | 181 | ±54 | 155 | ±46 | 153 | ±44 | 151 | ±43 | 151 | ±43 | 146 | ±41 | 140 | ±39 | 136 | ±38 | 134 | ±37 | ||
| Carpinus betulus | <10 | 489 | ±97 | 474 | ±95 | 468 | ±94 | 457 | ±92 | 455 | ±92 | 447 | ±91 | 447 | ±91 | 444 | ±91 | 431 | ±88 | 417 | ±87 | 406 | ±86 | |
| ≥10 | 154 | ±80 | 169 | ±80 | 175 | ±81 | 175 | ±81 | 175 | ±81 | 183 | ±84 | 183 | ±84 | 185 | ±82 | 193 | ±82 | 197 | ±82 | 205 | ±81 | ||
| All | 643 | ±145 | 643 | ±145 | 643 | ±145 | 633 | ±140 | 630 | ±139 | 630 | ±139 | 630 | ±139 | 628 | ±138 | 624 | ±136 | 614 | ±132 | 612 | ±130 | ||
| Alnus glutinosa | <10 | 90 | ±46 | 90 | ±46 | 86 | ±45 | 74 | ±38 | 56 | ±27 | 52 | ±24 | 52 | ±24 | 50 | ±25 | 39 | ±20 | 35 | ±17 | 33 | ±15 | |
| ≥10 | 325 | ±67 | 325 | ±67 | 315 | ±68 | 305 | ±67 | 293 | ±66 | 293 | ±67 | 293 | ±67 | 278 | ±65 | 267 | ±63 | 232 | ±57 | 222 | ±56 | ||
| All | 415 | ±99 | 415 | ±99 | 401 | ±99 | 378 | ±92 | 349 | ±84 | 345 | ±84 | 345 | ±84 | 328 | ±81 | 307 | ±75 | 268 | ±67 | 255 | ±65 | ||
| Other broadleaf | <10 | 158 | ±52 | 156 | ±52 | 154 | ±53 | 149 | ±51 | 147 | ±51 | 145 | ±50 | 145 | ±50 | 145 | ±50 | 141 | ±49 | 141 | ±49 | 141 | ±49 | |
| ≥10 | 15 | ±6 | 17 | ±6 | 19 | ±8 | 15 | ±6 | 15 | ±6 | 17 | ±6 | 17 | ±6 | 17 | ±6 | 15 | ±7 | 15 | ±7 | 15 | ±7 | ||
| All | 172 | ±53 | 172 | ±53 | 172 | ±53 | 164 | ±51 | 162 | ±51 | 162 | ±51 | 162 | ±51 | 162 | ±51 | 156 | ±51 | 156 | ±51 | 156 | ±51 | ||
| All species | <10 | 946 | ±109 | 913 | ±108 | 888 | ±108 | 839 | ±104 | 808 | ±95 | 784 | ±93 | 782 | ±93 | 766 | ±94 | 729 | ±89 | 708 | ±86 | 686 | ±84 | |
| ≥10 | 1085 | ±53 | 1118 | ±55 | 1114 | ±59 | 1065 | ±62 | 1054 | ±62 | 1070 | ±64 | 1070 | ±64 | 1046 | ±64 | 1023 | ±65 | 977 | ±64 | 969 | ±66 | ||
| All | 2031 | ±136 | 2031 | ±136 | 2002 | ±136 | 1903 | ±136 | 1862 | ±128 | 1854 | ±129 | 1852 | ±129 | 1812 | ±128 | 1752 | ±125 | 1686 | ±120 | 1655 | ±119 | ||
| G—basal area (m2 ha−1) | ||||||||||||||||||||||||
| Quercus robur | <10 | 0.5 | ±0.1 | 0.4 | ±0.1 | 0.4 | ±0.1 | 0.4 | ±0.1 | 0.4 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.0 | |
| ≥10 | 11.7 | ±1.3 | 12.4 | ±1.4 | 12.7 | ±1.4 | 13.2 | ±1.4 | 13.9 | ±1.5 | 14.5 | ±1.5 | 15.3 | ±1.6 | 15.7 | ±1.6 | 15.9 | ±1.6 | 16.4 | ±1.7 | 16.6 | ±1.7 | ||
| All | 12.2 | ±1.3 | 12.9 | ±1.4 | 13.1 | ±1.4 | 13.6 | ±1.4 | 14.3 | ±1.5 | 14.9 | ±1.5 | 15.6 | ±1.6 | 16.0 | ±1.6 | 16.2 | ±1.6 | 16.6 | ±1.7 | 16.8 | ±1.7 | ||
| Fraxinus angustifolia | <10 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | |
| ≥10 | 2.2 | ±0.6 | 2.4 | ±0.6 | 2.6 | ±0.7 | 2.2 | ±0.6 | 2.3 | ±0.6 | 2.4 | ±0.6 | 2.5 | ±0.7 | 2.6 | ±0.7 | 2.7 | ±0.7 | 2.7 | ±0.7 | 2.7 | ±0.7 | ||
| All | 2.5 | ±0.7 | 2.7 | ±0.7 | 2.9 | ±0.7 | 2.5 | ±0.6 | 2.6 | ±0.6 | 2.6 | ±0.7 | 2.8 | ±0.7 | 2.8 | ±0.7 | 2.9 | ±0.7 | 2.8 | ±0.7 | 2.9 | ±0.7 | ||
| Carpinus betulus | <10 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.3 | ±0.3 | 1.2 | ±0.2 | 1.2 | ±0.2 | |
| ≥10 | 2.3 | ±1.4 | 2.5 | ±1.4 | 2.6 | ±1.5 | 2.7 | ±1.5 | 2.8 | ±1.6 | 3.0 | ±1.6 | 3.0 | ±1.7 | 3.1 | ±1.7 | 3.2 | ±1.7 | 3.3 | ±1.7 | 3.4 | ±1.7 | ||
| All | 3.6 | ±1.5 | 3.8 | ±1.6 | 3.9 | ±1.6 | 4.0 | ±1.7 | 4.2 | ±1.7 | 4.3 | ±1.8 | 4.4 | ±1.8 | 4.4 | ±1.8 | 4.5 | ±1.8 | 4.6 | ±1.8 | 4.6 | ±1.8 | ||
| Alnus glutinosa | <10 | 0.4 | ±0.2 | 0.4 | ±0.2 | 0.4 | ±0.2 | 0.3 | ±0.2 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.1 | ±0.1 | 0.1 | ±0.1 | 0.1 | ±0.0 | |
| ≥10 | 6.0 | ±1.2 | 6.2 | ±1.3 | 6.0 | ±1.3 | 6.1 | ±1.3 | 6.1 | ±1.3 | 6.1 | ±1.3 | 6.1 | ±1.4 | 6.0 | ±1.4 | 5.8 | ±1.3 | 5.4 | ±1.3 | 5.3 | ±1.3 | ||
| All | 6.4 | ±1.3 | 6.6 | ±1.4 | 6.4 | ±1.4 | 6.4 | ±1.4 | 6.3 | ±1.4 | 6.3 | ±1.4 | 6.3 | ±1.4 | 6.1 | ±1.4 | 5.9 | ±1.4 | 5.5 | ±1.3 | 5.4 | ±1.3 | ||
| Other broadleaf | <10 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | |
| ≥10 | 0.2 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.2 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.3 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | 0.2 | ±0.1 | ||
| All | 0.5 | ±0.2 | 0.5 | ±0.2 | 0.5 | ±0.2 | 0.4 | ±0.1 | 0.4 | ±0.1 | 0.5 | ±0.1 | 0.5 | ±0.2 | 0.5 | ±0.2 | 0.4 | ±0.1 | 0.4 | ±0.1 | 0.4 | ±0.1 | ||
| All species | <10 | 2.8 | ±0.4 | 2.6 | ±0.4 | 2.6 | ±0.4 | 2.4 | ±0.4 | 2.3 | ±0.3 | 2.2 | ±0.3 | 2.2 | ±0.3 | 2.2 | ±0.3 | 2.0 | ±0.3 | 1.9 | ±0.2 | 1.8 | ±0.2 | |
| ≥10 | 22.5 | ±1.0 | 23.8 | ±1.0 | 24.3 | ±0.9 | 24.5 | ±1.0 | 25.4 | ±1.0 | 26.3 | ±1.1 | 27.3 | ±1.1 | 27.7 | ±1.1 | 27.9 | ±1.1 | 28.0 | ±1.2 | 28.3 | ±1.2 | ||
| All | 25.3 | ±1.0 | 26.5 | ±1.0 | 26.9 | ±1.0 | 26.9 | ±1.1 | 27.7 | ±1.1 | 28.5 | ±1.1 | 29.5 | ±1.2 | 29.9 | ±1.2 | 29.9 | ±1.2 | 29.9 | ±1.3 | 30.1 | ±1.3 | ||
| V—wood ** volume (m3 ha−1) | ||||||||||||||||||||||||
| Quercus robur | <10 | 3 | ±1 | 3 | ±1 | 3 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±0 | 2 | ±0 | 2 | ±0 | 1 | ±0 | |
| ≥10 | 115 | ±13 | 125 | ±14 | 130 | ±14 | 139 | ±15 | 149 | ±16 | 159 | ±17 | 171 | ±18 | 179 | ±18 | 185 | ±19 | 194 | ±20 | 200 | ±20 | ||
| All | 118 | ±13 | 128 | ±14 | 133 | ±14 | 142 | ±15 | 152 | ±16 | 161 | ±17 | 173 | ±18 | 181 | ±18 | 187 | ±19 | 196 | ±20 | 201 | ±20 | ||
| Fraxinus angustifolia | <10 | 2 | ±1 | 2 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±0 | 1 | ±0 | 1 | ±0 | 1 | ±0 | 1 | ±0 | 1 | ±0 | |
| ≥10 | 19 | ±5 | 21 | ±6 | 24 | ±6 | 20 | ±5 | 22 | ±6 | 23 | ±6 | 25 | ±6 | 26 | ±7 | 27 | ±7 | 27 | ±7 | 28 | ±7 | ||
| All | 21 | ±5 | 23 | ±6 | 25 | ±7 | 22 | ±5 | 23 | ±6 | 24 | ±6 | 26 | ±7 | 27 | ±7 | 28 | ±7 | 28 | ±7 | 29 | ±8 | ||
| Carpinus betulus | <10 | 7 | ±1 | 7 | ±1 | 7 | ±1 | 7 | ±1 | 7 | ±1 | 7 | ±1 | 7 | ±1 | 7 | ±2 | 7 | ±1 | 7 | ±1 | 6 | ±1 | |
| ≥10 | 20 | ±13 | 22 | ±13 | 24 | ±14 | 26 | ±15 | 27 | ±16 | 29 | ±17 | 30 | ±17 | 31 | ±18 | 33 | ±19 | 35 | ±19 | 36 | ±19 | ||
| All | 27 | ±13 | 29 | ±14 | 31 | ±15 | 33 | ±16 | 34 | ±17 | 36 | ±17 | 37 | ±18 | 39 | ±19 | 40 | ±19 | 42 | ±20 | 42 | ±20 | ||
| Alnus glutinosa | <10 | 3 | ±2 | 3 | ±2 | 3 | ±2 | 2 | ±1 | 2 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±0 | 1 | ±0 | |
| ≥10 | 58 | ±12 | 60 | ±12 | 59 | ±12 | 60 | ±13 | 61 | ±13 | 61 | ±13 | 62 | ±14 | 61 | ±14 | 60 | ±14 | 56 | ±14 | 56 | ±14 | ||
| All | 60 | ±13 | 63 | ±13 | 61 | ±13 | 62 | ±14 | 62 | ±14 | 62 | ±14 | 63 | ±14 | 62 | ±14 | 61 | ±14 | 56 | ±14 | 56 | ±14 | ||
| Other broadleaf | <10 | 1 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | 0 | ±0 | |
| ≥10 | 2 | ±1 | 2 | ±1 | 3 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±1 | ||
| All | 3 | ±1 | 3 | ±1 | 3 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 2 | ±1 | 1 | ±1 | 1 | ±1 | 1 | ±1 | ||
| All species | <10 | 15 | ±2 | 14 | ±2 | 14 | ±2 | 13 | ±2 | 13 | ±2 | 12 | ±2 | 12 | ±2 | 12 | ±2 | 10 | ±2 | 10 | ±1 | 9 | ±1 | |
| ≥10 | 214 | ±10 | 231 | ±10 | 240 | ±9 | 247 | ±11 | 261 | ±11 | 274 | ±11 | 290 | ±12 | 300 | ±12 | 306 | ±13 | 313 | ±14 | 321 | ±14 | ||
| All | 230 | ±10 | 245 | ±10 | 253 | ±10 | 260 | ±11 | 274 | ±11 | 286 | ±12 | 302 | ±12 | 311 | ±13 | 317 | ±13 | 323 | ±14 | 330 | ±15 | ||
References
- Canadell, J.G.; Le Quere, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.A.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar] [CrossRef] [PubMed]
- Nabuurs, G.J.; Lindner, M.; Verkerk, P.J.; Gunia, P.J.; Deda, P.; Michalak, R.; Grassi, G. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Chang. 2013, 3, 792–796. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Cullman, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Baldocchi, D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 1–14. [Google Scholar] [CrossRef]
- Carrara, A.; Janssens, I.A.; Curiel Yuste, J.; Ceulemans, R. Seasonal changes in photosynthesis, respiration and nee of a mixed temperate forest. Agric. For. Meteorol. 2004, 126, 15–31. [Google Scholar] [CrossRef]
- Granier, A.; Breda, N.; Longdoz, B.; Gross, P.; Ngao, J. Ten years of fluxes and stand growth in a young beech forest at Hesse, North-Eastern France. Ann. For. Sci. 2008, 65, 704–717. [Google Scholar] [CrossRef]
- Baldocchi, D.; Chu, H.; Reichstein, M. Inter-annual variability of net and gross ecosystem carbon fluxes: A review. Agric. For. Meteorol. 2018, 249, 520–533. [Google Scholar] [CrossRef]
- Teets, A.; Fraver, S.; Hollinger, D.Y.; Weiskittel, A.R.; Seymour, R.S.; Richardson, A.D. Linking annual tree growth with eddy-flux measures of net ecosystem productivity across twenty years of observation in a mixed conifer forest. Agric. For. Meteorol. 2018, 249, 479–487. [Google Scholar] [CrossRef]
- Running, S.W.; Hunt, E.R.J. Generalization of a forest ecosystem process model for other biomes, BIOME-BGC, and an application for global-scale models. In Scaling Physiological Processes: Leaf to Globe; Ehleringer, J.R., Field, C., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 141–158. ISBN 012233440X. [Google Scholar]
- Hidy, D.; Barcza, Z.; Marjanović, H.; Ostrogović Sever, M.Z.; Dobor, L.; Gelybo, G.; Fodor, N.; Pinter, K.; Churkina, G.; Running, S.; et al. Terrestrial Ecosystem Process Model Biome-BGCMuSo v4.0: Summary of improvements and new modelling possibilities. Geosci. Model. Dev. 2016, 9, 4405–4437. [Google Scholar] [CrossRef]
- Running, S.W.; Nemani, R.R.; Heinsch, F.A.; Zhao, M.; Reeves, M.; Hashimoto, H. A Continuous Satellite-Derived Measure of Global Terrestrial Primary Production. BioScience 2004, 54, 547–560. [Google Scholar] [CrossRef]
- FLUXNET2015 Dataset. Available online: http://fluxnet.fluxdata.org/data/ (accessed on 7 September 2018).
- Burba, G. Eddy Covariance Method for Scientific, Industrial, Agricultural and Regulatory Applications. A Field Book on Measuring Ecosystem Gas. Exchange and Areal Emission Rates; LI-COR Biosciences: Lincoln, NE, USA, 2013; p. 332. ISBN 978-0-615-76827-4. [Google Scholar]
- Schulze, E.-D. The Carbon and Nitrogen Cycle of Forest Ecosystems. In Carbon and Nitrogen Cycling in European Forest Ecosystems, Ecological Studies; Schulze, E.-D., Ed.; Springer: Berlin/Heidelberg, Germany, 2000; Volume 142, pp. 3–13. ISBN 978-3-642-57219-7. [Google Scholar]
- Brunner, I.; Godbold, D.L. Tree roots in a changing world. J. For. Res. 2007, 12, 78–82. [Google Scholar] [CrossRef]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, P.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Saigusa, N.; Koizumi, H. On linking multiyear biometric measurements of tree growth with eddy covariance-based net ecosystem production. Glob. Chang. Biol. 2009, 15, 1015–1024. [Google Scholar] [CrossRef]
- Curtis, P.S.; Hanson, P.J.; Bolstad, P.; Barford, C.; Randolph, J.C.; Schmid, H.P.; Wilson, K.B. Biometric and eddy-covariance based estimates of annual carbon storage in five Eastern North American deciduous forests. Agric. For. Meteorol. 2002, 113, 3–19. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Tschaplinski, T.J.; Wullschleger, S.D.; Joslin, J.D. Estimating the Net Primary and Net Ecosystem Production of a Southeastern Upland Quercus Forest from an 8-Year Biometric Record. In North American Temperate Deciduous Forest Responses to Changing Precipitation Regimes; Ecological Studies (Analysis and Synthesis), Volume 166; Hanson, P.J., Wullschleger, S.D., Eds.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Rocha, A.V.; Goulden, M.L.; Dunn, A.L.; Wofsy, S.C. On linking interannual tree ring variability with observations of whole-forest CO2 flux. Glob. Chang. Biol. 2006, 12, 1378–1389. [Google Scholar] [CrossRef]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Su, H.B.; Curtis, P.S. Multi-year convergence of biometric and meteorological estimates of forest carbon storage. Agric. For. Meteorol. 2008, 148, 158–170. [Google Scholar] [CrossRef]
- Gough, C.M.; Flower, C.E.; Vogel, C.S.; Dragoni, D.; Curtis, P.S. Whole-ecosystem labile carbon production in a north temperate deciduous forest. Agric. For. Meteorol. 2009, 149, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Saleska, S.R.; Miller, S.D.; Matross, D.M.; Goulden, M.L.; Wofsy, S.C.; da Rocha, H.R.; de Camargo, P.B.; Crill, P.; Daube, B.C.; de Freitas, H.C.; et al. Carbon in amazon forests: Unexpected seasonal fluxes and disturbance-induced losses. Science 2003, 302, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Gough, C.M.; Vogel, C.S.; Schmid, H.P.; Curtis, P.S. Controls on annual forest carbon storage: Lessons from the past and predictions for the future. Bioscience 2008, 58, 609–622. [Google Scholar] [CrossRef]
- Urbanski, S.; Barford, C.; Wofsy, S.; Kucharik, C.; Pyle, E.; Budney, J.; McKain, K.; Fitzjarrald, D.; Czikowsky, M.; Munger, W. Factors controlling CO2 exchange on timescales from hourly to decadal at Harvard Forest. J. Geophys. Res. 2007, 112, G02020. [Google Scholar] [CrossRef]
- Van der Molen, M.K.; Dolman, A.J.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.E.; Meir, P.; Peters, W.; Phillips, O.L.; Reichstein, M.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol. 2011, 2011 151, 765–773. [Google Scholar] [CrossRef]
- Carbone, M.S.; Czimczik, C.I.; Keenan, T.F.; Murakami, P.F.; Pederson, N.; Schaberg, P.G.; Xu, X.; Richardson, A.D. Age, allocation and availability of nonstructural carbon in mature red maple trees. New Phytol. 2013, 200, 1145–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, A.; Marjanović, H.; Dobor, L.; Anić, M.; Hlásny, T.; Barcza, Z. Identification of Years with Extreme Vegetation State in Central Europe Based on Remote Sensing and Meteorological Data. South.-East Eur. For. 2017, 8, 1–20. [Google Scholar] [CrossRef]
- Ostrogović Sever, M.Z.; Paladinić, E.; Barcza, Z.; Hidy, D.; Kern, A.; Anić, M.; Marjanović, H. Biogeochemical Modelling vs. Tree-Ring Measurements—Comparison of Growth Dynamic Estimates at Two Distinct Oak Forests in Croatia. South.-East Eur. For. 2017, 8, 71–84. [Google Scholar] [CrossRef]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Babst, F.; Bouriaud, O.; Papale, D.; Gielen, B.; Janssens, I.A.; Nikinmaa, E.; Ibrom, A.; Wu, J.; Bernhofer, C.; Kostner, B.; et al. Above-ground woody carbon sequestration measured from tree rings is coherent with net ecosystem productivity at five eddy-covariance sites. New Phytol. 2014, 201, 1289–1303. [Google Scholar] [CrossRef]
- Barford, C.C.; Wofsy, S.C.; Goulden, M.L.; Munger, J.W.; Pyle, E.H.; Urbanski, S.P.; Hutyra, L.; Saleska, S.R.; Fitzjarrald, D.; Moore, K. Factors controlling long- and short-term sequestration of atmospheric CO2 in a mid-latitude forest. Science 2001, 294, 1688–1691. [Google Scholar] [CrossRef]
- Klepac, D.; Fabijanić, G. Management of Pedunculate Oak Forests. In Pedunculate Oak in Croatia, 1st ed.; Klepac, D., Dundović, J., Gračan, J., Eds.; Croatian Academy of Arts and Sciences, Centre for Scientific Work, Vinkovci and Croatian Forests Ltd.: Zagreb, Croatia, 1996; pp. 452–457. ISBN 953-154-079-9. [Google Scholar]
- Ministry of Agriculture of the Republic of Croatia, Forest Management Area Plan of the Republic of Croatia; Croatian Forests Ltd.: Zagreb, Croatia, 2017; p. 859. Available online: http://www.mps.hr/hr/sume/sumarstvo/sumskogospodarska-osnova-2016-2025 (accessed on 12 September 2018).
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, Update 2015; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; p. 192. ISBN 978-92-5-108369-7. [Google Scholar]
- Mayer, B. Hydropedological relations in the region of lowland forests of the Pokupsko basin. Rad. Šum. Inst. Jastrebarsko 1996, 31, 37–89. (In Croatian) [Google Scholar]
- Marjanović, H.; Ostrogović, M.Z.; Alberti, G.; Balenović, I.; Paladinić, E.; Indir, K.; Peressotti, A.; Vuletić, D. Carbon dynamics in younger stands of Pedunculate oak during two vegetation periods. Šum. List (Spec. Issue) 2011, 135, 59–73, (In Croatian with English summary). [Google Scholar]
- Ostrogović Sever, M.Z. Carbon Stocks and Carbon Balance of an Even-Aged Pedunculate Oak (Quercus robur L.) Forest in Kupa River Basin. Ph.D. Thesis, Faculty of Forestry, University of Zagreb, Zagreb, Croatia, 2013. [Google Scholar]
- Wilkinson, M.; Eaton, E.L.; Broadmeadow, M.S.J.; Morison, J.I.L. Inter-annual variation of carbon uptake by a plantation oak woodland in south-eastern England. Biogeosciences 2012, 9, 5373–5389. [Google Scholar] [CrossRef] [Green Version]
- Vickers, D.; Mahrt, L. Quality Control and Flux Sampling Problems for Tower and Aircraft Data. J. Atmos. Ocean. Technol. 1997, 14, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Aubinet, M.; Grelle, A.; Ibrom, A.; Rannik, U.; Moncrieff, J.; Foken, T.; Kowalski, A.S.; Martin, P.H.; Berbigier, P.; Bernhofer, C.; et al. Estimates of the annual net carbon and water exchange of forests: The euroflux methodology. Adv. Ecol. Res. 2000, 30, 113–175. [Google Scholar]
- Wilczak, J.M.; Oncley, S.P. Sonic anemometer tilt correction algorithms. Bound.-Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef]
- Baldocchi, D.; Valentini, R.; Running, S.; Oechel, W.; Dahlman, R. Strategies for measuring and modelling carbon dioxide and water vapour fluxes over terrestrial ecosystems. Glob. Chang. Biol. 1996, 2, 159–168. [Google Scholar] [CrossRef]
- Eddy Covariance Gap-Filling & Flux-Partitioning Tool. Available online: https://www.bgc-jena.mpg.de/bgi/index.php/Services/REddyProcWeb (accessed on 1 June 2018).
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Sigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the temperature-dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Van Dijk, A.J.I.M.; Dolman, A.J. Estimates of CO2 uptake and release among European forests based on eddy covariance data. Glob. Chang. Biol. 2004, 10, 1445–1459. [Google Scholar] [CrossRef]
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A simple two-dimensional parameterisation for Flux Footprint Prediction (FFP). Geosci. Model. Dev. 2015, 8, 3695–3713. [Google Scholar] [CrossRef] [Green Version]
- Flesch, T.K.; Wilson, J.D.; Yee, E. Backward-time lagrangian stochastic dispersion models and their application to estimate gaseous emissions. J. Appl. Meteorol. Climatol. 1995, 34, 1320–1332. [Google Scholar] [CrossRef]
- Kljun, N.; Rotach, M.W.; Schmid, H.P. A three-dimensional backward Lagrangian footprint model for a wide range of boundary-layer stratifications. Bound.-Lay Meteorol. 2002, 103, 205–226. [Google Scholar] [CrossRef]
- Flux Footprint Prediction (FFP) Online Data Processing. Available online: http://geography.swansea.ac.uk/nkljun/ffp/www/ (accessed on 13 April 2018).
- Keeland, B.D.; Young, P.J. Installation of Traditional Dendrometer Bands; National Wetlands Research Center: Lafayette, LA, USA, 2015. Available online: https://www.nwrc.usgs.gov/topics/Dendrometer/ (accessed on 15 August 2018).
- Marjanović, H. Modelling tree development and elements of stand structure in young stands of Pedunculate oak (Quercus Robur L.). Ph.D. Thesis, Faculty of Forestry, University of Zagreb, Zagreb, Croatia, 10 May 2009; p. 213, (In Croatian with English Summary). [Google Scholar]
- Michailoff, I. Zahlenmäßiges Verfahren für die Ausführung der Bestandeshöhenkurven. [Numerical algorithm for the implementation of stand height curves]. Forstw. Cbl. u Thar. Jahrb. 1943, 6, 273–279. (In German) [Google Scholar]
- Schumacher, F.X.; Hall, F.D.S. Logarithmic expression of timber-tree volume. J. Agric. Res. 1933, 47, 719–734. [Google Scholar]
- Cestar, D.; Kovačić, Đ. Wood volume tables for Black Alder and Black Locust. Rad. Šum. Inst. Jastrebarsko 1982, 49, 1–149, (In Croatian with English Summary). [Google Scholar]
- Špiranec, M. Wood volume tables. Rad. Šum. Inst. Jastrebarsko 1975, 22, 1–262, (In Croatian with German Summary). [Google Scholar]
- Cestar, D.; Kovačić, Đ. Wood volume tables for Narrow-leaved Ash (Fraxinus parvifolia Auct.). Rad. Šum. Inst. Jastrebarsko 1984, 60, 1–178, (In Croatian with English Summary). [Google Scholar]
- Balboa-Murias, M.A.; Rojo, A.; Álvarez, J.G.; Merino, A. Carbon and nutrient stocks in mature Quercus robur L. stands in NW Spain. Ann. For. Sci. 2006, 63, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef]
- Šumarska Enciklopedija, 1st ed.; Leksikografski zavod FNRJ: Zagreb, Croatia, 1959.
- IPCC. Good Practice Guidance for Land Use, Land-Use Change and Forestry; Penman, J., Gytarsky, M., Hiraishi, T., Kruger, D., Pipatti, R., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Wagner, F., et al., Eds.; IPCC/IGES: Hayama, Japan, 2003. [Google Scholar]
- IPCC. 2006 IPCC Guidelines for National Greenhouse Inventories; Eggleston, H.S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; IGES: Kobe, Japan, 2006. [Google Scholar]
- Martínez-Vilalta, J. Carbon storage in trees: Pathogens have their say. Tree Physiol. 2014, 34, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Locosselli, G.M.; Buckeridge, M.S. Dendrobiochemistry, a missing link to further understand carbon allocation during growth and decline of trees. Trees-Struct. Funct. 2017, 31, 1745–1758. [Google Scholar] [CrossRef]
- Jolliffe, I.T. A note on the use of principal components in regression. J. R. Stat. Soc. Ser. C Appl. Stat. 1982, 31, 300–303. [Google Scholar] [CrossRef]
- Pretzsch, H. Forest Dynamics, Growth and Yield: From Measurement to Model; Springer: Berlin, Germany, 2009. [Google Scholar]
- Pilegaard, K.; Ibrom, A.; Courtney, M.S.; Hummelshøj, P.; Jensen, N.O. Increasing net CO2 uptake by a Danish beech forest during the period from 1996 to 2009. Agric. For. Meteorol. 2011, 151, 934–946. [Google Scholar] [CrossRef]
- Herbst, M.; Mund, M.; Tamrakar, R.; Knohl, A. Differences in carbon uptake and water use between a managed and an unmanaged beech forest in central Germany. For. Ecol. Manag. 2015, 355, 101–108. [Google Scholar] [CrossRef]
- Novick, K.A.; Oishi, A.C.; Ward, E.J.; Siqueira, M.B.S.; Juang, J.-Y.; Stoy, P.C. On the difference in the net ecosystem exchange of CO2 between deciduous and evergreen forests in the southeastern United States. Glob. Chang. Biol. 2015, 21, 827–842. [Google Scholar] [CrossRef]
- Sulman, B.N.; Roman, D.T.; Scanlon, T.M.; Wang, L.; Novick, K.A. Comparing methods for partitioning a decade of carbon dioxide and water vapor fluxes in a temperate forest. Agirc. For. Meteorol. 2016, 226–227, 229–245. [Google Scholar] [CrossRef]
- Malhi, Y.; Baldocchi, D.; Jarvis, P.G. The carbon balance of tropical, temperate and boreal forests. Plant Cell Environ. 1999, 22, 715–740. [Google Scholar] [CrossRef] [Green Version]
- Law, B.E.; Falge, E.; Gu, L.; Baldocchi, D.D.; Bakwin, P.; Berbigier, P.; Davis, K.; Dolman, A.J.; Falk, M.; Fuentes, J.D.; et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol. 2002, 113, 97–120. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Berveiller, D.; Granda, E.; Dufrene, E. Wood phenology, not carbon input, controls the interannual variability of wood growth in a temperate oak forest. New Phytol. 2016, 210, 459–470. [Google Scholar] [CrossRef]
- Froelich, N.; Croft, H.; Chen, J.M.; Gonsamo, A.; Staebler, R.M. Trends of carbon fluxes and climate over a mixed temperate-boreal transition forest in southern Ontario, Canada. Agric. For. Meteorol. 2015, 211, 72–84. [Google Scholar] [CrossRef]
- Saigusa, N.; Yamamoto, S.; Murayama, S.; Kondo, H. Inter-annual variability of carbon budget components in an AsiaFlux forest site estimated by long-term flux measurements. Agric. For. Meteorol. 2005, 134, 4–16. [Google Scholar] [CrossRef]
- Zscheischler, J.; Fatichi, S.; Wolf, S.; Blanken, P.D.; Bohrer, G.; Clark, K.; Desai, A.R.; Hollinger, D.; Keenan, T.; Novick, K.A.; et al. Short-term favorable weather conditions are an important control of interannual variability in carbon and water fluxes. J. Geophys. Res.-Biogeosci. 2016, 121, 2186–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odum, E.P. Strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.S.; Gough, C.M. Forest aging, disturbance and the carbon cycle. New Phytol. 2018, 219, 1188–1193. [Google Scholar] [CrossRef]
- Gough, C.M.; Vogel, C.S.; Hardiman, B.; Curtis, P.S. Wood net primary production resilience in an unmanaged forest transitioning from early to middle succession. For. Ecol. Manag. 2010, 260, 36–41. [Google Scholar] [CrossRef]
- Peterken, G.F. Woodland Conservation and Management, 2nd ed.; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Ortmann-Ajkai, A.; Csicsek, G.; Lukacs, M.; Horvath, F. Regeneration patterns in a pedunculate oak (Quercus robur L.) strict forest reserve in southern Hungary. Šum. List 2017, 141, 39–46. [Google Scholar]
- Gower, S.T.; McMurtrie, R.E.; Murty, D. Aboveground net primary production decline with stand age: Potential causes. Trends Ecol. Evol. 1996, 11, 378–382. [Google Scholar] [CrossRef]
- Xu, C.Y.; Turnbull, M.H.; Tissue, D.T.; Lewis, J.D.; Carson, R.; Schuster, W.S.F.; Whitehead, D.; Walcroft, A.S.; Li, J.B.; Griffin, K.L. Age-related decline of stand biomass accumulation is primarily due to mortality and not to reduction in NPP associated with individual tree physiology, tree growth or stand structure in a quercus-dominated forest. J. Ecol. 2012, 100, 428–440. [Google Scholar] [CrossRef]
- Barić, L.; Županić, M.; Pernek, M.; Diminić, D. First records of Chalara fraxinea in Croatia—A new agent of ash dieback (Fraxinus spp.). Šum. list 2012, 136, 461–469. [Google Scholar]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef]
- Black, K.; Bolger, T.; Davis, P.; Nieuwenhuis, M.; Reidy, B.; Saiz, G.; Tobin, B.; Osborne, B. Inventory and eddy covariance-based estimates of annual carbon sequestration in a sitka spruce (Picea sitchensis (Bong.) Carr.) forest ecosystem. Eur. J. For. Res. 2007, 126, 167–178. [Google Scholar] [CrossRef]
- Peichl, M.; Brodeur, J.J.; Khomik, M.; Arain, M.A. Biometric and eddy-covariance based estimates of carbon fluxes in an age-sequence of temperate pine forests. Agric. For. Meteorol. 2010, 150, 952–965. [Google Scholar] [CrossRef]
- Campioli, M.; Malhi, Y.; Vicca, S.; Luyssaert, S.; Papale, D.; Penuelas, J.; Reichstein, M.; Migliavacca, M.; Arain, M.A.; Janssens, I.A. Evaluating the convergence between eddy-covariance and biometric methods for assessing carbon budgets of forests. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Kutsch, W.; Bahn, M.; Heinemeyer, A. Soil Carbon Dynamics: An Integrated Methodology; Cambridge University Press: Cambridge, UK, 2009; p. 286. ISBN 978-0-521-86561-6. [Google Scholar]
- Ohtsuka, T.; Mo, W.H.; Satomura, T.; Inatomi, M.; Koizumi, H. Biometric based carbon flux measurements and net ecosystem production (NEP) in a temperate deciduous broad-leaved forest beneath a flux tower. Ecosystems 2007, 10, 324–334. [Google Scholar] [CrossRef]
- Subke, J.A.; Inglima, I.; Cotrufo, M.F. Trends and methodological impacts in soil CO2 efflux partitioning: A meta-analytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Bailey, V.L.; Chen, M.; Gough, C.M.; Vargas, R. Globally rising soil heterotrophic respiration over recent decades. Nature 2018, 560, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Yamaji, K.; Ishida, A.; Prokushkin, S.G.; Masyagina, O.V.; Hagihara, A.; Hoque, A.; Suwa, R.; Osawa, A.; Nishizono, T. Mixed-power scaling of whole-plant respiration from seedlings to giant trees. Proc. Natl. Acad. Sci. USA 2010, 107, 1447–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, D.P.; Ritts, W.D.; Cohen, W.B.; Maeirsperger, T.K.; Gower, S.T.; Kirschbaum, A.A.; Running, S.W.; Zhao, M.; Wofsy, S.C.; Dunn, A.L.; et al. Site-level evaluation of satellite-based global terrestrial gross primary production and net primary production monitoring. Glob. Chang. Biol. 2005, 11, 666–684. [Google Scholar] [CrossRef]
- Neumann, M.; Zhao, M.; Kindermann, G.; Hasenauer, H. Comparing MODIS net primary production estimates with terrestrial national forest inventory data in Austria. Remote Sens. 2015, 7, 3878. [Google Scholar] [CrossRef]
- Kern, A.; Marjanović, H.; Barcza, Z. Evaluation of the Quality of NDVI3g Dataset against Collection 6 MODIS NDVI in Central Europe between 2000 and 2013. Remote Sens. 2016, 8, 955. [Google Scholar] [CrossRef]
- Gaudinski, J.B.; Torn, M.S.; Riley, W.J.; Swanston, C.; Trumbore, S.E.; Joslin, J.D.; Majdi, H.; Dawson, T.E.; Hanson, P.J. Use of stored carbon reserves in growth of temperate tree roots and leaf buds: Analyses using radiocarbon measurements and modeling. Glob. Chang. Biol. 2009, 15, 992–1014. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands; London, UK; Boston, MA, USA, 2008. [Google Scholar]
- Barbaroux, C.; Breda, N.; Dufrene, E. Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (Quercus petraea and Fagus sylvatica). New Phytol. 2003, 157, 605–615. [Google Scholar] [CrossRef]
- Palacio, S.; Camarero, J.J.; Maestro, M.; Alla, A.Q.; Lahoz, E.; Montserrat-Martí, G. Are storage and tree growth related? Seasonal nutrient and carbohydrate dynamics in evergreen and deciduous Mediterranean oaks. Trees 2018, 32, 777–790. [Google Scholar] [CrossRef]
- Begum, S.; Kudo, K.; Rahman, M.H.; Nakaba, S.; Yamagishi, Y.; Nabeshima, E.; Nugroho, W.D.; Oribe, Y.; Kitin, P.; Jin, H.O.; et al. Climate change and the regulation of wood formation in trees by temperature. Trees-Struct. Funct. 2018, 32, 3–15. [Google Scholar] [CrossRef]












| Year | Age | P | Ta | Ts | Rg | SWC | ΔSWCMay–Sep | SGS | EGS | NEE ± SE | GPP | RECO | Ra | Rh | NPPEC | NPPSB ± SE | NPPT | NPPR | NPPL ± SE | NPPF ± SE | NPPBM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2008 | 35 | 900 | 11.34 | 11.12 | 153 | 0.448 | 0.233 | 77 | 295 | −352 ± 13 | 1469 | 1117 | 679 | 438 | 790 | 387 ± 15 | 19 | 104 | 164 ± 20 | 1 ± 0 | 675 |
| 2009 | 36 | 940 | 11.25 | 11.23 | 155 | 0.420 | 0.197 | 98 | 293 | −496 ± 15 | 1622 | 1126 | 685 | 441 | 937 | 413 ± 18 | 21 | 111 | 207 ± 10 | 4 ± 1 | 756 |
| 2010 | 37 | 1255 | 10.22 | 10.26 | 143 | 0.518 | 0.072 | 86 | 287 | −286 ± 14 | 1615 | 1329 | 808 | 521 | 807 | 430 ± 18 | 21 | 116 | 207 ± 15 | 9 ± 4 | 783 |
| 2011 | 38 | 576 | 11.05 | 10.63 | 160 | 0.403 | 0.195 | 88 | 295 | −353 ± 14 | 1642 | 1289 | 784 | 505 | 858 | 395 ± 17 | 20 | 107 | 199 ± 11 | 18 ± 5 | 739 |
| 2012 | 39 | 987 | 11.49 | 11.06 | 162 | 0.419 | 0.071 | 90 | 296 | −261 ± 14 | 1642 | 1382 | 840 | 541 | 802 | 351 ± 16 | 18 | 95 | 165 ± 8 | 1 ± 0 | 630 |
| 2013 | 40 | 1238 | 10.92 | 10.52 | 148 | 0.493 | 0.124 | 111 | 291 | −356 ± 13 | 1630 | 1275 | 775 | 500 | 855 | 403 ± 26 | 20 | 109 | 204 ± 12 | 20 ± 6 | 756 |
| 2014 | 41 | 1755 | 12.11 | 11.82 | 140 | 0.586 | −0.011 | 90 | 272 | −232 ± 14 | 1522 | 1290 | 785 | 506 | 738 | 359 ± 17 | 18 | 97 | 174 ± 8 | 6 ± 2 | 654 |
| 2015 | 42 | 1260 | 11.47 | 11.10 | 152 | 0.502 | 0.212 | 86 | 278 | −373 ± 13 | 1775 | 1402 | 852 | 549 | 923 | 310 ± 13 | 16 | 84 | 162 ± 13 | 60 ± 25 | 632 |
| 2016 | 43 | 744 | 11.11 | 11.01 | 146 | 0.512 | 0.273 | 101 | 282 | −339 ± 13 | 1644 | 1305 | 794 | 512 | 850 | 393 ± 19 | 20 | 106 | 166 ± 5 | 4 ± 1 | 689 |
| 2017 | 44 | 927 | 11.39 | 10.76 | 164 | 0.487 | 0.049 | 100 | 292 | −147 ± 13 | 1384 | 1236 | 752 | 484 | 632 | 227 ± 16 | 11 | 61 | 183 ± 5 | 1 ± 0 | 483 |
| Aver. | 1058 | 11.24 | 10.95 | 152 | 0.479 | 0.141 | 93 | 288 | −319 | 1594 | 1275 | 775 | 500 | 819 | 367 | 18 | 99 | 183 | 12 | 680 | |
| SD | (332) | (0.48) | (0.43) | (8) | (0.053) | (0.089) | (10) | (8) | (94) | (109) | (94) | (57) | (37) | (89) | (60) | (3) | (16) | (19) | (18) | (88) |
| DBH * (cm) | Year (at the End of the Season) | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | |||||||||||||||||||||||
| Stand age (years) | |||||||||||||||||||||||||||||||||
| 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | 45 | |||||||||||||||||||||||
| d100—quadratic mean diameter of 100 thickest trees per hectare (cm) | |||||||||||||||||||||||||||||||||
| 25.8 | 26.5 | 26.4 | 26.7 | 27.4 | 28.5 | 28.5 | 29.8 | 29.4 | 30.0 | 30.3 | |||||||||||||||||||||||
| h100—top height ** (m) | |||||||||||||||||||||||||||||||||
| 20.8 | 21.3 | 21.6 | 21.9 | 22.3 | 22.7 | 23.1 | 23.5 | 23.9 | 24.3 | 24.6 | |||||||||||||||||||||||
| N—stand density (trees ha−1) | |||||||||||||||||||||||||||||||||
| <10 | 946 | ±109 | 913 | ±108 | 888 | ±108 | 839 | ±104 | 808 | ±95 | 784 | ±93 | 782 | ±93 | 766 | ±94 | 729 | ±89 | 708 | ±86 | 686 | ±84 | |||||||||||
| ≥10 | 1085 | ±53 | 1118 | ±55 | 1114 | ±59 | 1065 | ±62 | 1054 | ±62 | 1070 | ±64 | 1070 | ±64 | 1046 | ±64 | 1023 | ±65 | 977 | ±64 | 969 | ±66 | |||||||||||
| All | 2031 | ±136 | 2031 | ±136 | 2002 | ±136 | 1903 | ±136 | 1862 | ±128 | 1854 | ±129 | 1852 | ±129 | 1812 | ±128 | 1752 | ±125 | 1686 | ±120 | 1655 | ±119 | |||||||||||
| G—basal area (m2 ha−1) | |||||||||||||||||||||||||||||||||
| <10 | 2.8 | ±0.4 | 2.6 | ±0.4 | 2.6 | ±0.4 | 2.4 | ±0.4 | 2.3 | ±0.3 | 2.2 | ±0.3 | 2.2 | ±0.3 | 2.2 | ±0.3 | 2.0 | ±0.3 | 1.9 | ±0.2 | 1.8 | ±0.2 | |||||||||||
| ≥10 | 22.5 | ±1.0 | 23.8 | ±1.0 | 24.3 | ±0.9 | 24.5 | ±1.0 | 25.4 | ±1.0 | 26.3 | ±1.1 | 27.3 | ±1.1 | 27.7 | ±1.1 | 27.9 | ±1.1 | 28.0 | ±1.2 | 28.3 | ±1.2 | |||||||||||
| All | 25.3 | ±1.0 | 26.5 | ±1.0 | 26.9 | ±1.0 | 26.9 | ±1.1 | 27.7 | ±1.1 | 28.5 | ±1.1 | 29.5 | ±1.2 | 29.9 | ±1.2 | 29.9 | ±1.2 | 29.9 | ±1.3 | 30.1 | ±1.3 | |||||||||||
| V—wood *** volume (m3 ha−1) | |||||||||||||||||||||||||||||||||
| <10 | 15 | ±2 | 14 | ±2 | 14 | ±2 | 13 | ±2 | 13 | ±2 | 12 | ±2 | 12 | ±2 | 12 | ±2 | 10 | ±2 | 10 | ±1 | 9 | ±1 | |||||||||||
| ≥10 | 214 | ±10 | 231 | ±10 | 240 | ±9 | 247 | ±11 | 261 | ±11 | 274 | ±11 | 290 | ±12 | 300 | ±12 | 306 | ±13 | 313 | ±14 | 321 | ±14 | |||||||||||
| All | 230 | ±10 | 245 | ±10 | 253 | ±10 | 260 | ±11 | 274 | ±11 | 286 | ±12 | 302 | ±12 | 311 | ±13 | 317 | ±13 | 323 | ±14 | 330 | ±15 | |||||||||||
| Country, Site | Genus | Period | NEE | GPP | RECO | Reference |
|---|---|---|---|---|---|---|
| gC m−2 year−1 | ||||||
| Denmark, Sorø | Fagus | 1996–2009 | −156 ± 103 | 1727 ± 136 | 1570 ± 97 | [72] |
| France, Hesse | Fagus | 1995–2005 | −386 ± 171 | 1397 ± 192 | 1011 ± 138 | [6] |
| Germany, Hainich | Fagus, Fraxinus | 2003–2012 | −483 ± 70 | 1498 ± 83 | 1015 ± 51 | [73] |
| UK, Straits Inclosure | Quercus | 1999–2010 | −486 ± 115 | 2034 ± 228 | 1548 ± 192 | [40] |
| US, Duke Forest | Quercus, Carya | 2001–2008 | −402 ± 96 | 1982 ± 300 | 1580 ± 237 | [74] |
| US, Harvard Forest | Acer, Quercus | 1992–2004 | −242 ± 100 | 1400 ± 164 | 1153 ± 105 | [26] |
| Croatia, Jastrebarsko | Quercus | 2008–2017 | −319 ± 94 | 1594 ± 109 | 1275 ± 94 | this study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anić, M.; Ostrogović Sever, M.Z.; Alberti, G.; Balenović, I.; Paladinić, E.; Peressotti, A.; Tijan, G.; Večenaj, Ž.; Vuletić, D.; Marjanović, H. Eddy Covariance vs. Biometric Based Estimates of Net Primary Productivity of Pedunculate Oak (Quercus robur L.) Forest in Croatia during Ten Years. Forests 2018, 9, 764. https://doi.org/10.3390/f9120764
Anić M, Ostrogović Sever MZ, Alberti G, Balenović I, Paladinić E, Peressotti A, Tijan G, Večenaj Ž, Vuletić D, Marjanović H. Eddy Covariance vs. Biometric Based Estimates of Net Primary Productivity of Pedunculate Oak (Quercus robur L.) Forest in Croatia during Ten Years. Forests. 2018; 9(12):764. https://doi.org/10.3390/f9120764
Chicago/Turabian StyleAnić, Mislav, Maša Zorana Ostrogović Sever, Giorgio Alberti, Ivan Balenović, Elvis Paladinić, Alessandro Peressotti, Goran Tijan, Željko Večenaj, Dijana Vuletić, and Hrvoje Marjanović. 2018. "Eddy Covariance vs. Biometric Based Estimates of Net Primary Productivity of Pedunculate Oak (Quercus robur L.) Forest in Croatia during Ten Years" Forests 9, no. 12: 764. https://doi.org/10.3390/f9120764
APA StyleAnić, M., Ostrogović Sever, M. Z., Alberti, G., Balenović, I., Paladinić, E., Peressotti, A., Tijan, G., Večenaj, Ž., Vuletić, D., & Marjanović, H. (2018). Eddy Covariance vs. Biometric Based Estimates of Net Primary Productivity of Pedunculate Oak (Quercus robur L.) Forest in Croatia during Ten Years. Forests, 9(12), 764. https://doi.org/10.3390/f9120764