Spatial Segregation Facilitates the Coexistence of Tree Species in Temperate Forests



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
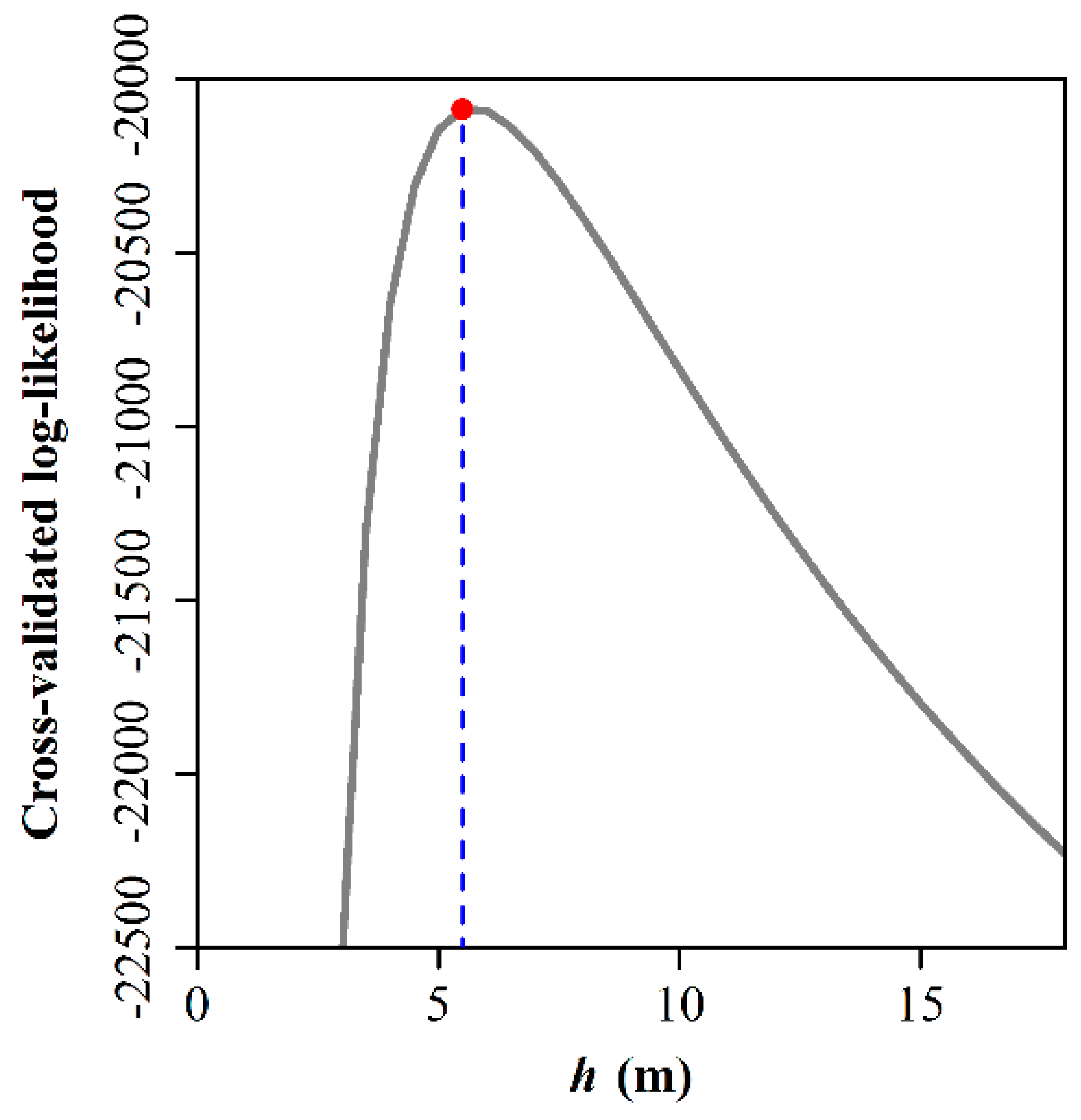
2.2. Statistical Methods
2.2.1. Species-Specific Probabilities for Spatial Segregation
2.2.2. Significant Test for Spatial Segregation
2.3. Analysis and Test of Spatial Relationship
3. Results
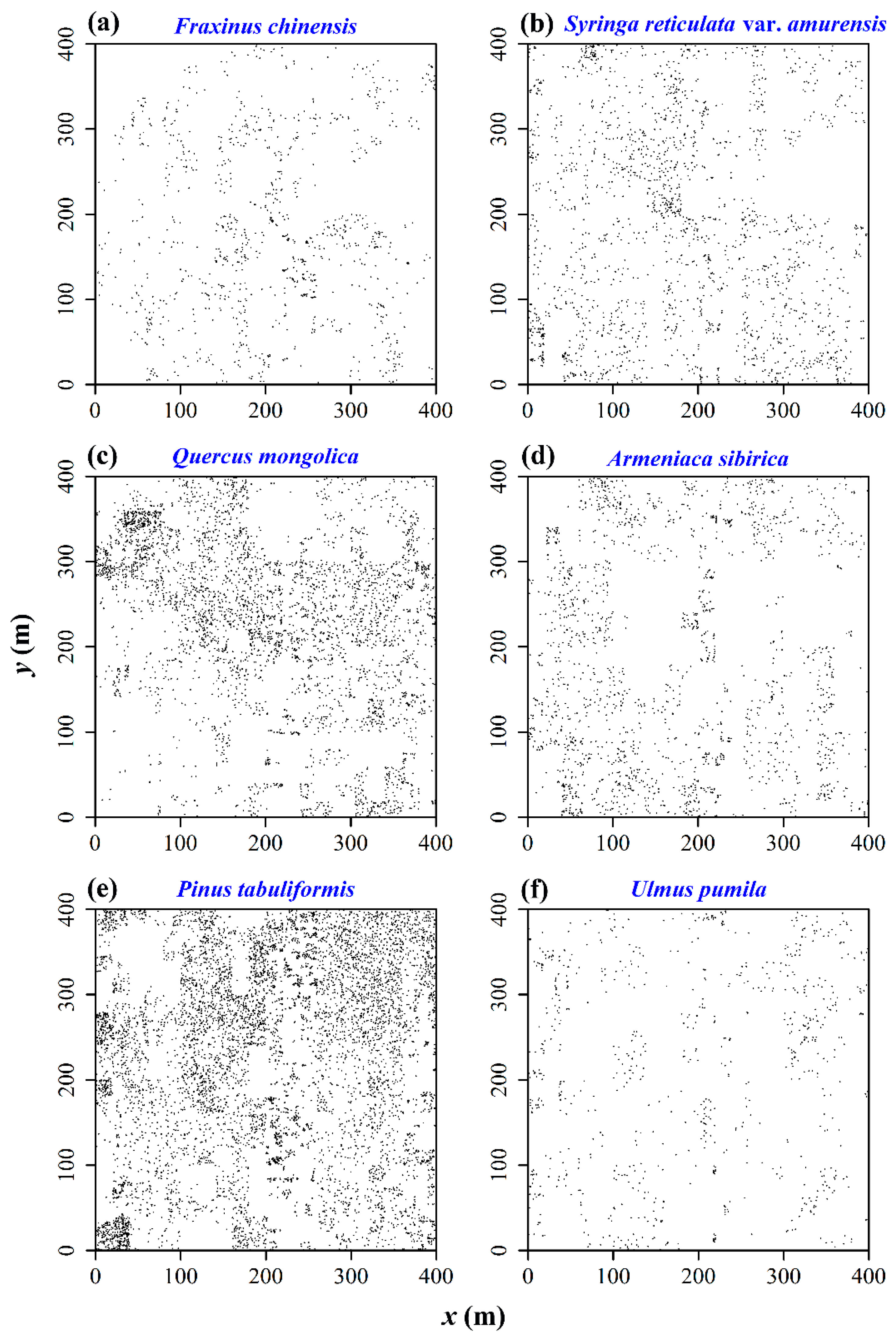
3.1. Abundances of Tree Species
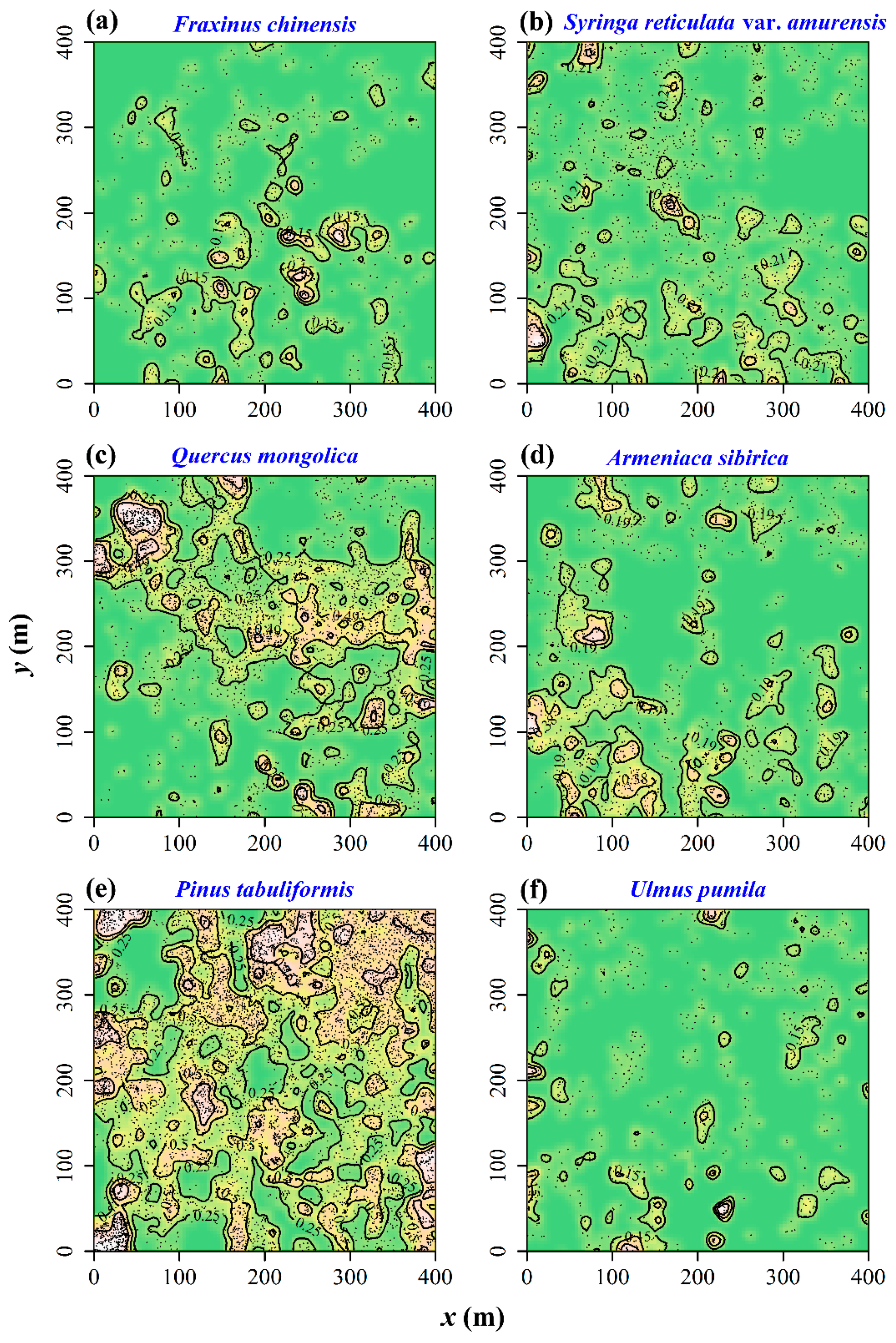
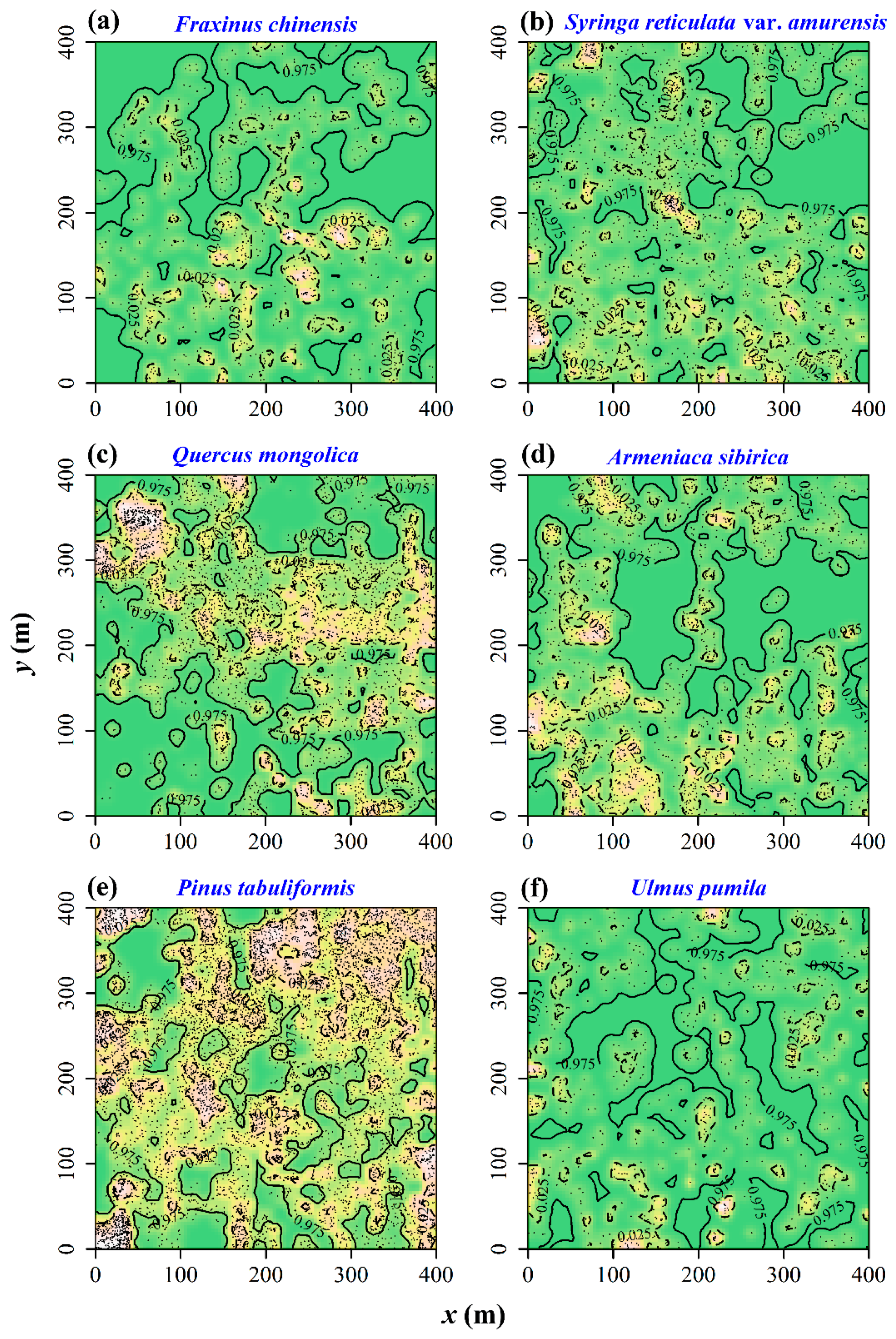
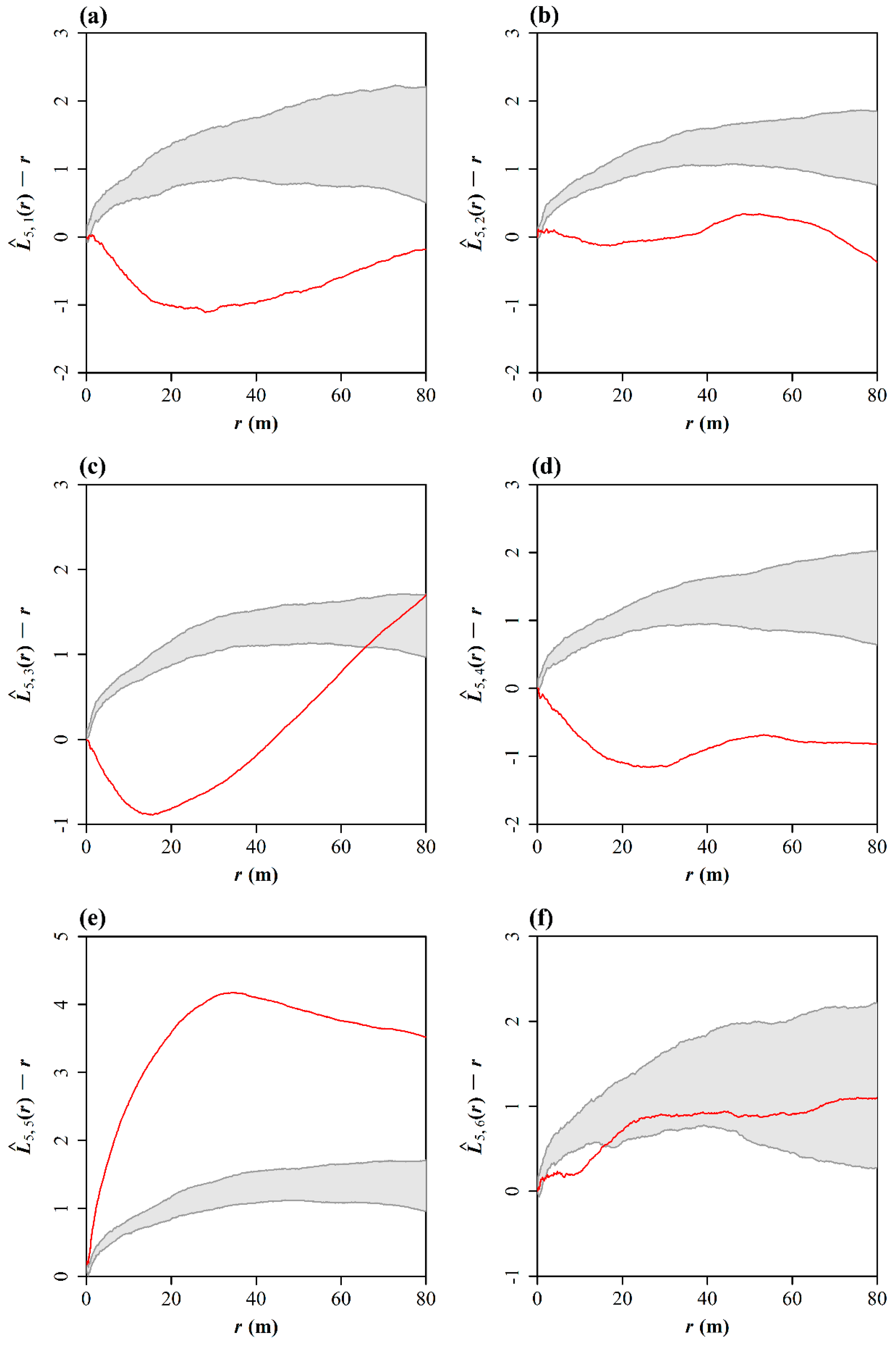
3.2. Test of Spatial Segregation
3.3. Spatial Correlations between the Chinese Pine and Other Five Tree Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Sork, V.L.; Wu, J.; Ge, J. Effect of patch size and isolation on mating patterns and seed production in an urban population of Chinese pine (Pinus tabuliformis Carr.). For. Ecol. Manag. 2010, 260, 965–974. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Godron, M. Patches and structural components for a landscape ecology. Bioscience 1981, 31, 733–740. [Google Scholar]
- DeCesare, N.J.; Hebblewhite, M.; Robinson, H.S.; Musiani, M. Endangered, apparently: The role of apparent competition in endangered species conservation. Anim. Conserv. 2010, 13, 353–362. [Google Scholar] [CrossRef]
- Hui, C.; Richardson, D.M. Invasion Dynamics; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Chu, C.; Kleinhesselink, A.R.; Havstad, K.M.; McClaran, M.P.; Peters, D.P.; Vermeire, L.T.; Wei, H.Y.; Adler, P.B. Direct effects dominate responses to climate perturbations in grassland plant communities. Nat. Commun. 2016, 7, 11766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Lehman, C.L.; Yin, C.J. Habitat destruction, dispersal, and deterministic extinction in competitive communities. Am. Nat. 1997, 149, 407–435. [Google Scholar] [CrossRef]
- Lehman, C.L.; Tilman, D. Biodiversity, stability, and productivity in competitive communities. Am. Nat. 2000, 156, 534–552. [Google Scholar] [CrossRef]
- Parry, G.D. The meanings of r- and K-selection. Oecologia 1981, 48, 260–264. [Google Scholar] [CrossRef]
- Adler, P.B.; HilleRisLambers, J.; Levine, J.M. A niche for neutrality. Ecol. Lett. 2007, 10, 95–104. [Google Scholar] [CrossRef]
- Chesson, P. General theory of competitive coexistence in spatially-varying environments. Theor. Popul. Biol. 2000, 58, 211–237. [Google Scholar] [CrossRef]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. Dyn. Popul. 1971, 298, 298–312. [Google Scholar]
- Diggle, P.J. Statistical Analysis of Spatial and Spatio-temporal Point Patterns; CRC Press: London, UK, 2014. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; Chapman & Hall/CRC: London, UK, 2015. [Google Scholar]
- Hui, C.; Landi, P.; Minoarivelo, H.O.; Ramanantoanina, A. Ecological and Evolutionary Modelling; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Baddeley, A.; Diggle, P.J.; Hardegen, A.; Lawrence, T.; Milne, R.K.; Nair, G. On tests of spatial pattern based on simulation envelopes. Ecol. Monogr. 2014, 84, 477–489. [Google Scholar] [CrossRef]
- Ripley, B.D. Modelling spatial patterns (with discussion). J. R. Stat. Soc. B 1977, 39, 172–212. [Google Scholar]
- Hui, C.; Veldtman, R.; McGeoch, M.A. Measures, perceptions and scaling patterns of aggregated species distributions. Ecography 2010, 33, 95–102. [Google Scholar] [CrossRef]
- Hui, C.; Vermeulen, W.; Durrheim, G. Quantifying multiple-site compositional turnover in an Afrotemperate forest, using zeta diversity. For. Ecosyst. 2018, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Diggle, P.; Zheng, P.P.; Durr, P. Nonparametric estimation of spatial segregation in a multivariate point process: Bovine tuberculosis in Cornwall, UK. J. R. Stat. Soc. Ser. C Appl. Stat. 2005, 54, 645–658. [Google Scholar] [CrossRef]
- Sandhu, H.S.; Shi, P.; Yang, Q. Intraspecific spatial niche differentiation: Evidence from Phyllostachys edulis. Acta Ecol. Sin. 2013, 33, 287–292. [Google Scholar] [CrossRef]
- Serra, L.; Juan, P.; Varga, D.; Mateu, J.; Saez, M. Spatial pattern modelling of wildfires in Catalonia, Spain 2004–2008. Environ. Model. Softw. 2013, 40, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.B.; Shi, P.J.; Reddy, G.V.P.; Li, L.M.; Men, X.Y.; Ge, F. Nonparametric estimation of interspecific spatio-temporal niche separation between two lady beetles (Coleoptera: Coccinellidae) in Bt cotton fields. Ann. Entomol. Soc. Am. 2015, 108, 807–813. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Y. Age-independent climate-growth response of Chinese pine (Pinus tabuliformis Carrière) in North China. Trees Struc. Funct. 2015, 29, 397–406. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Song, H.; Li, Q.; Burr, G.S.; Wang, L.; Hu, S. A monsoon-related 174-year relative humidity record from tree-ring in δ18O the Yaoshan region, eastern central China. Sci. Total Environ. 2017, 593−594, 523–534. [Google Scholar] [CrossRef]
- Kelsall, J.E.; Diggle, P.J. Spatial variation in risk of disease: A nonparametric binary regression approach. Appl. Stat. 1998, 47, 559–573. [Google Scholar] [CrossRef]
- Hui, C.; McGeoch, M.A.; Warren, M. A spatially explicit approach to estimating species occupancy and spatial correlation. J. Anim. Ecol. 2006, 75, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goreaud, F.; Pélissier, R. Avoiding misinterpretation of biotic interactions with the intertype K12-function: Population independence vs. random labelling hypotheses. J. Veg. Sci. 2003, 14, 681–692. [Google Scholar] [CrossRef]
- Grabarnik, P.; Myllymäki, M.; Stoyan, D. Correct testing of mark independence for marked point patterns. Ecol. Model. 2011, 222, 3888–3894. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R.-project.org (accessed on 17 April 2018).
- Ghalandarayeshi, S.; Nord-Larsen, T.; Johannsen, V.K.; Larsen, J.B. Spatial patterns of tree species in Suserup Skov—A semi-natural forest in Denmark. For. Ecol. Manag. 2017, 406, 391–401. [Google Scholar] [CrossRef]
- Xiang, W.; Liu, S.; Lei, X.; Frank, S.C.; Tian, D.; Wang, G.; Deng, X. Secondary forest floristic composition, structure, and spatial pattern in subtropical China. J. For. Res. 2013, 18, 111–120. [Google Scholar] [CrossRef]
- Legendre, P.; Fortin, M.-J. Spatial pattern and ecological analysis. Plant Ecol. 1989, 80, 107–138. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yang, H.; Dong, B.; Zhou, M.; Ma, L.; Jia, Z.; Duan, J. Effects of canopy gap size on growth and spatial patterns of Chinese pine (Pinus tabuliformis) regeneration. For. Ecol. Manag. 2017, 385, 46–56. [Google Scholar] [CrossRef]
- Diggle, P.J.; Gómez-Rubio, V.; Brown, P.E.; Chetwynd, A.G.; Gooding, S. Second-order analysis of inhomogeneous spatial point processes using case–control data. Biometrics 2007, 63, 550–557. [Google Scholar] [CrossRef]
- Shi, P.J.; Sandhu, H.S.; Reddy, G.V.P. Dispersal distance determines the exponent of the spatial Taylor’s power law. Ecol. Model. 2016, 335, 48–53. [Google Scholar] [CrossRef]
- Hou, H.; Ding, L.; Xu, Z.; Wang, L.; Zhao, Y. Root distribution of young trees of typical species in the northern region of Yanshan Mountains. For. Resour. Manag. 2018, 10–15. (In Chinese) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Code | Latin Name | Family | Abundance (0 ≤ y ≤ 1000 m) | Abundance (0 ≤ y ≤ 400 m) | Abundance (600 ≤ y ≤ 1000 m) | DBH (cm) | Height (m) |
|---|---|---|---|---|---|---|---|
| 1 | Acer mono Maxim. | Aceraceae | 482 | 280 | 195 | 7.38 ± 4 | 5.9 ± 2.29 |
| 2 | Acer truncatum Bunge | Aceraceae | 228 | 124 | 92 | 7.66 ± 5.63 | 5.53 ± 3.08 |
| 3 | Berberis beijingensis Ying | Berberidaceae | 1 | 1 | 0 | 2.5 ± NA | 1.75 ± NA |
| 4 | Betula dahurica Pall. | Betulaceae | 205 | 138 | 62 | 13.92 ± 7.07 | 7.64 ± 3.12 |
| 5 | Betula platyphylla Suk. | Betulaceae | 89 | 42 | 46 | 17.43 ± 4.91 | 10.33 ± 1.7 |
| 6 | Carpinus turczaninowii Hance | Betulaceae | 8 | 1 | 4 | 7.65 ± 2.98 | 7.04 ± 1.05 |
| 7 | Chimonanthus praecox (L.) Link | Calycanthaceae | 3 | 3 | 0 | 5.47 ± 3.41 | 3.01 ± 0.37 |
| 8 | Sambucus williamsii Hance | Caprifoliaceae | 6 | 4 | 0 | 4.17 ± 0.94 | 2.56 ± 0.46 |
| 9 | Diospyros lotus L. | Ebenaceae | 2 | 0 | 0 | 20.83 ± 2.32 | 8.72 ± 0.49 |
| 10 | Lithocarpus cleistocarpus (Seemen) Rehd. & E. H. Wils. | Fagaceae | 19 | 15 | 4 | 5.38 ± 0.53 | 5.78 ± 0.6 |
| 11 | Quercus mongolica Fisch. ex Ledeb. | Fagaceae | 8466 | 4016 | 3064 | 11.07 ± 5.74 | 6.33 ± 2.73 |
| 12 | Juglans cathayensis Dode | Juglandaceae | 29 | 13 | 12 | 10.54 ± 10.66 | 8.1 ± 5.64 |
| 13 | Juglans mandshurica Maxim. | Juglandaceae | 824 | 376 | 211 | 11.84 ± 8.23 | 8.03 ± 3.9 |
| 14 | Albizia kalkora (Roxb.) Prain | Leguminosae | 156 | 93 | 59 | 6.98 ± 5.53 | 5.65 ± 2.94 |
| 15 | Hibiscus mutabilis L. | Malvaceae | 7 | 1 | 5 | 6.16 ± 0.73 | 6.16 ± 0.13 |
| 16 | Myrtus communis L. | Myrtaceae | 24 | 17 | 7 | 11.6 ± 0.6 | 7.52 ± 0.57 |
| 17 | Fraxinus chinensis Roxb. | Oleaceae | 1903 | 994 | 688 | 7.04 ± 4.26 | 5.27 ± 2.49 |
| 18 | Fraxinus mandshurica Rupr. | Oleaceae | 30 | 12 | 8 | 9.87 ± 6.81 | 7.85 ± 3.78 |
| 19 | Fraxinus rhynchophylla Hance | Oleaceae | 743 | 277 | 208 | 7.63 ± 4.87 | 5.6 ± 2.48 |
| 20 | Syringa pekinensis Rupr. | Oleaceae | 1 | 1 | 0 | 12.1 ± NA | 7.35 ± NA |
| 21 | Syringa reticulata (Blume) H. Hara var. amurensis (Rupr.) J. S. Pringle | Oleaceae | 4618 | 1935 | 1668 | 7.26 ± 4.35 | 5.27 ± 2.39 |
| 22 | Pinus tabuliformis Carrière | Pinaceae Lindl. | 24,579 | 7539 | 11,534 | 12.85 ± 6.02 | 8.55 ± 2.83 |
| 23 | Rhamnus davurica Pall. | Rhamnaceae | 2 | 0 | 1 | 7 ± 0.71 | 6.7 ± 0.14 |
| 24 | Ziziphus jujuba Mill. | Rhamnaceae | 62 | 18 | 37 | 11.1 ± 7.52 | 5.49 ± 2.97 |
| 25 | Ziziphus jujuba Mill. var. spinosa (Bunge) Hu ex H. F. Chow | Rhamnaceae | 3 | 0 | 2 | 14.63 ± 6.01 | 5.9 ± 2.72 |
| 26 | Amygdalus davidiana (Carrière) de Vos ex Henry | Rosaceae | 9 | 5 | 4 | 8.62 ± 4.26 | 6.65 ± 1.35 |
| 27 | Armeniaca mume Sieb. | Rosaceae | 4 | 4 | 0 | 4.98 ± 0.43 | 6.88 ± 0.16 |
| 28 | Armeniaca sibirica (L.) Lam. | Rosaceae | 6227 | 1789 | 2602 | 8.35 ± 4.55 | 5.04 ± 2.56 |
| 29 | Crataegus pinnatifida Bunge | Rosaceae | 1 | 0 | 0 | 10.5 ± NA | 5.17 ± NA |
| 30 | Malus baccata (L.) Borkh. | Rosaceae | 21 | 6 | 12 | 10.84 ± 12.52 | 4.96 ± 3.73 |
| 31 | Prunus salicina Lindl. | Rosaceae | 1 | 1 | 0 | 3.7 ± NA | 2.8 ± NA |
| 32 | Pyrus bretschneideri Rehd. | Rosaceae | 17 | 14 | 0 | 6.64 ± 0.75 | 4.32 ± 0.61 |
| 33 | Sorbus alnifolia (Siebold & Zucc.) C. Koch | Rosaceae | 121 | 18 | 42 | 8.73 ± 5.04 | 6.02 ± 2.66 |
| 34 | Sorbus discolor (Maxim.) Maxim. | Rosaceae | 492 | 71 | 168 | 7.71 ± 4.33 | 6.43 ± 2.46 |
| 35 | Leptodermis potanini Batal. | Rubiaceae | 220 | 19 | 62 | 4.82 ± 2.88 | 4.61 ± 1.84 |
| 36 | Populus davidiana Dode | Salicaceae | 37 | 23 | 11 | 9.97 ± 8.44 | 7.24 ± 2.94 |
| 37 | Ailanthus altissima (Mill.) Swingle | Simaroubaceae | 36 | 29 | 7 | 9.46 ± 7.68 | 7.71 ± 5.43 |
| 38 | Tilia amurensis Rupr. | Tiliaceae | 687 | 434 | 202 | 9.27 ± 4.87 | 6.76 ± 2.68 |
| 39 | Tilia mandshurica Rupr. & Maxim. | Tiliaceae | 2 | 0 | 2 | 12.35 ± 2.76 | 8.07 ± 2.22 |
| 40 | Celtis bungeana Blume | Ulmaceae | 655 | 403 | 181 | 5.61 ± 4.24 | 4.77 ± 2.39 |
| 41 | Hemiptelea davidii (Hance) Planch. | Ulmaceae | 12 | 0 | 1 | 6.71 ± 5.43 | 4.7 ± 2.53 |
| 42 | Ulmus davidiana Planch. var. japonica (Rehder) Nakai | Ulmaceae | 230 | 133 | 61 | 6.9 ± 4.42 | 5.78 ± 3.24 |
| 43 | Ulmus macrocarpa Hance | Ulmaceae | 133 | 5 | 31 | 7.71 ± 4.56 | 6.21 ± 2.33 |
| 44 | Ulmus pumila L. | Ulmaceae | 2124 | 826 | 729 | 12.79 ± 7.89 | 8.34 ± 3.66 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, P.; Gao, J.; Song, Z.; Liu, Y.; Hui, C. Spatial Segregation Facilitates the Coexistence of Tree Species in Temperate Forests. Forests 2018, 9, 768. https://doi.org/10.3390/f9120768
Shi P, Gao J, Song Z, Liu Y, Hui C. Spatial Segregation Facilitates the Coexistence of Tree Species in Temperate Forests. Forests. 2018; 9(12):768. https://doi.org/10.3390/f9120768
Chicago/Turabian StyleShi, Peijian, Jie Gao, Zhaopeng Song, Yanhong Liu, and Cang Hui. 2018. "Spatial Segregation Facilitates the Coexistence of Tree Species in Temperate Forests" Forests 9, no. 12: 768. https://doi.org/10.3390/f9120768
APA StyleShi, P., Gao, J., Song, Z., Liu, Y., & Hui, C. (2018). Spatial Segregation Facilitates the Coexistence of Tree Species in Temperate Forests. Forests, 9(12), 768. https://doi.org/10.3390/f9120768