Phenology of Five Shrub Communities along an Elevation Gradient in the Qilian Mountains, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
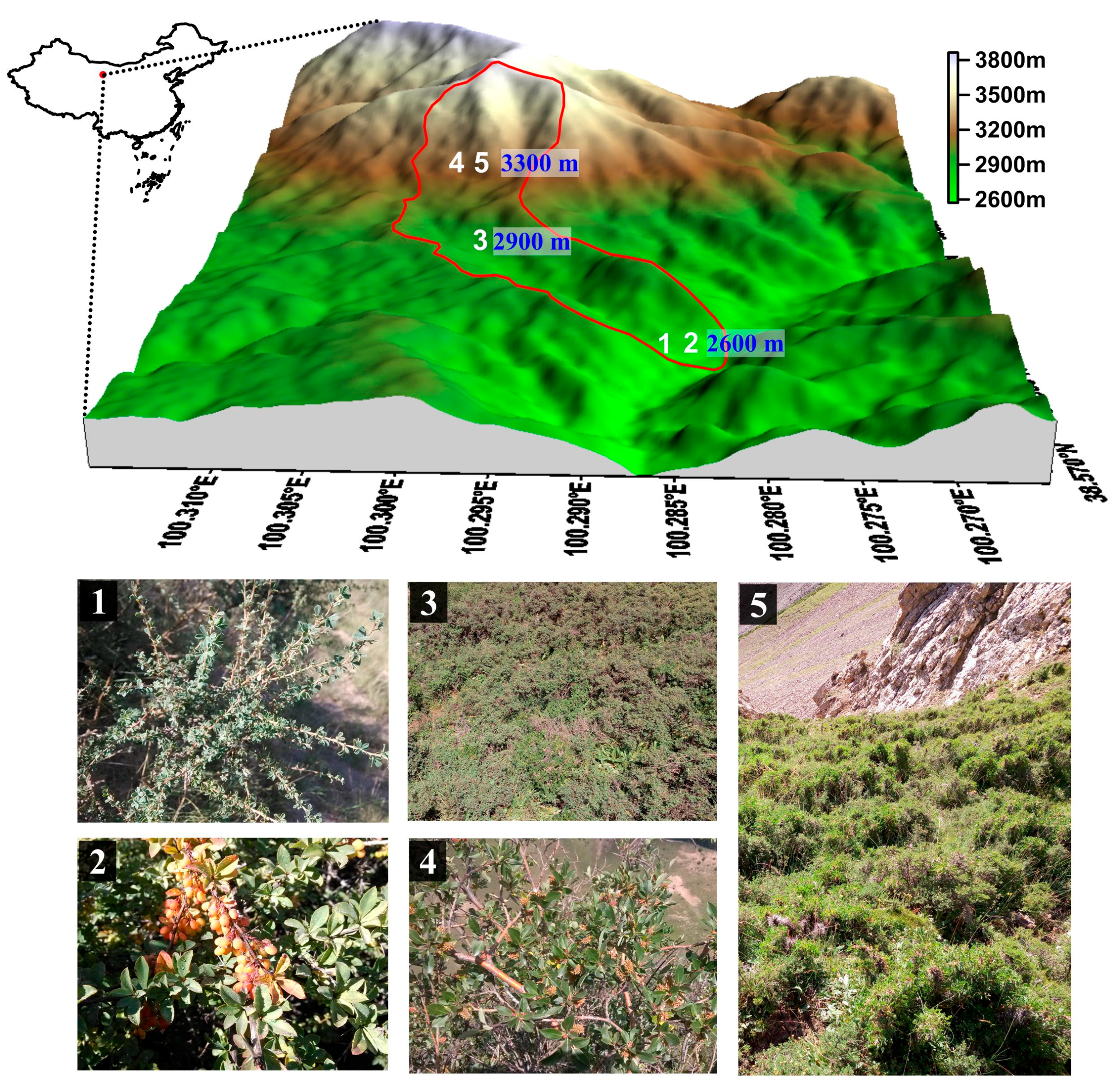
2.1. Study Site
2.2. Phenological Observations
2.3. Sample Plot Investigation
2.4. Data Processing
3. Results
3.1. Growth Parameters of Five Shrub Species
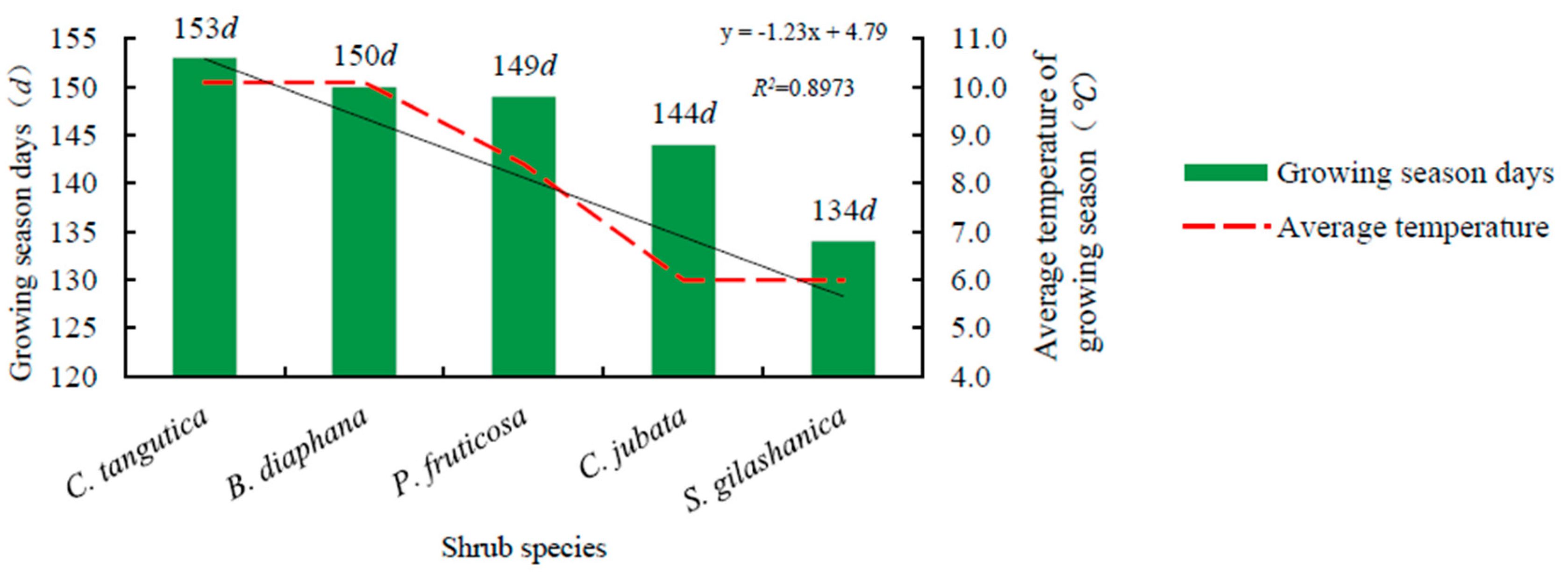
3.2. Duration of Phenological Stages of the Five Shrub Species
3.3. Variability in Temperature and Tacc
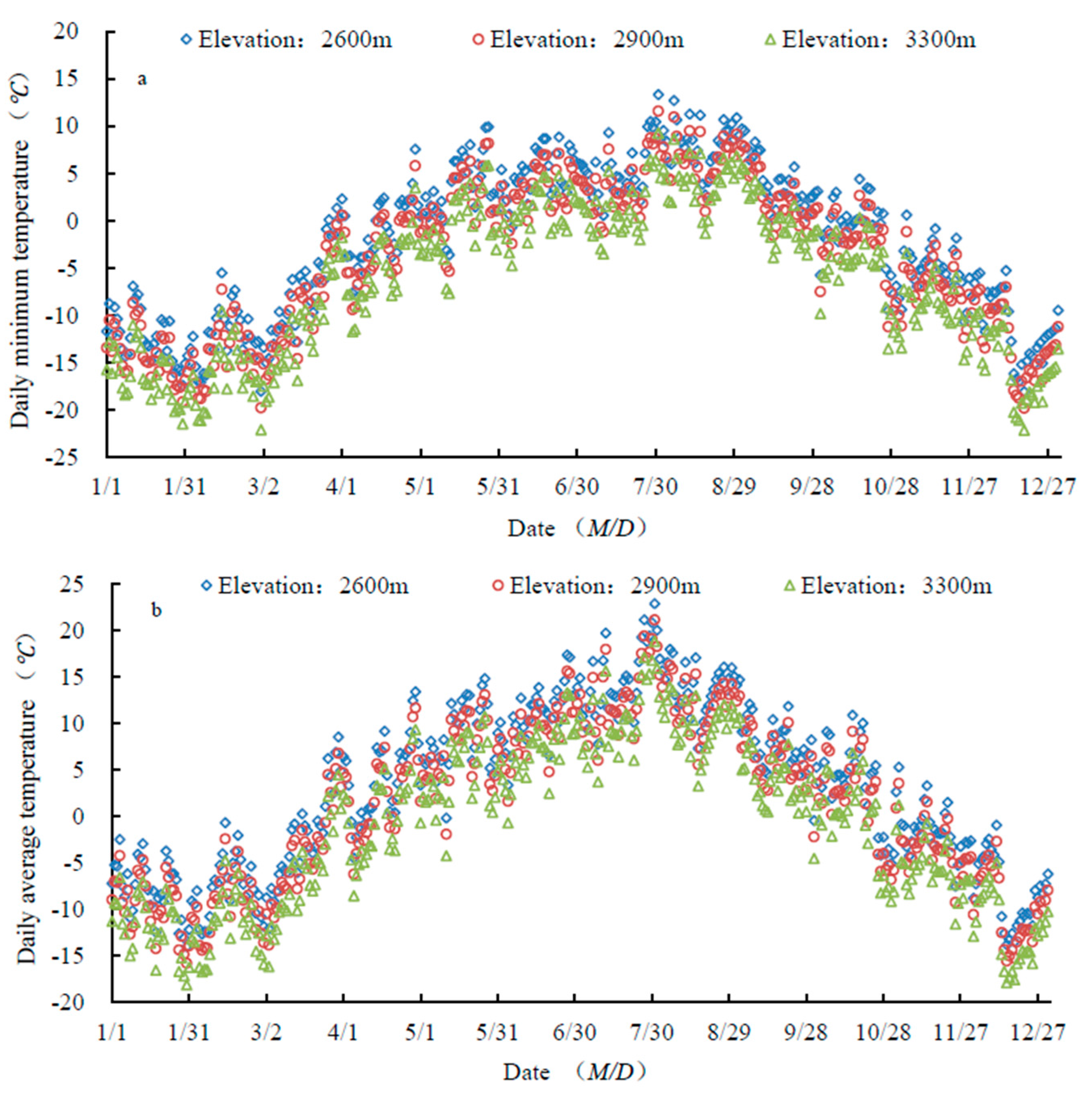
3.3.1. Changes in Temperature
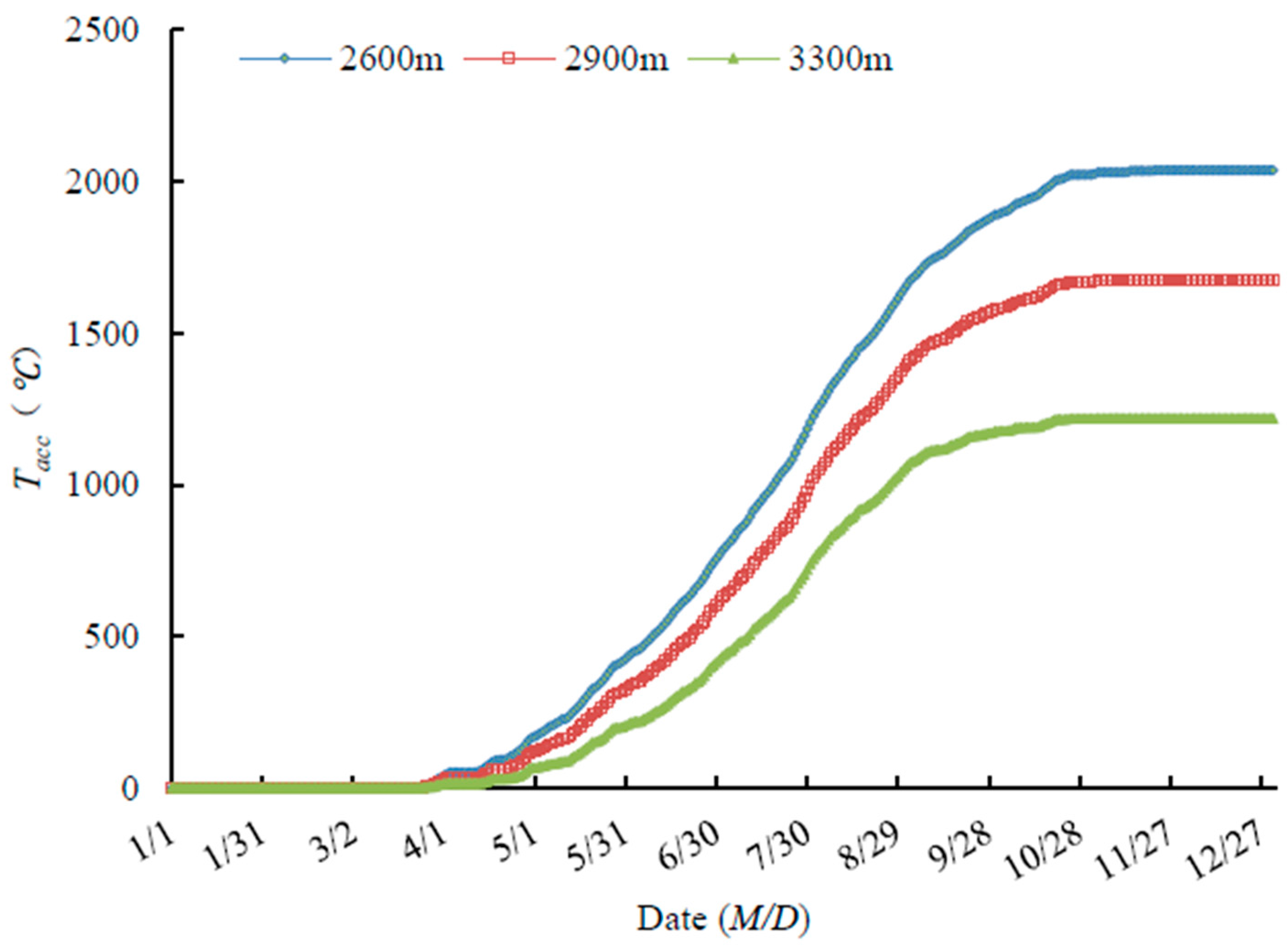
3.3.2. Variability in Tacc in 2015
3.3.3. Tacc of Different Phenophases
4. Discussion
4.1. Phenophases of Shrub Species
4.2. Effects of Temperature on Shrub Phenology
4.3. Effects of Topography on Shrub Phenology
4.3.1. Elevation
4.3.2. Slope Aspect
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vitasse, Y.; Delzon, S.; Dufrêne, E.; Pontaillerc, J.Y.; Louveta, J.M.; Kremera, A.; Michaleta, R. Leaf phenology sensitivity to temperature in European trees: Do within-species populations exhibit similar responses? Agric. For. Meteorol. 2009, 149, 735–744. [Google Scholar] [CrossRef]
- Luo, Z.K.; Osbert, J.S.; Ge, Q.S.; Xu, W.T.; Zheng, J.Y. Phenological responses of plants to climate change in an urban environment. Ecol. Res. 2007, 22, 507–514. [Google Scholar] [CrossRef]
- Josiane, S.; Aude, V.; Karine, P.; Remy, S.; Marc, A.; Nicolas, B.; Patricia, D.R.; Sylvie, G.; Marielle, G.; Abakar, H.M.; et al. Relationships between climate, soil moisture and phenology of the woody cover in two sites located along the West African latitudinal gradient. J. Hydrol. 2009, 375, 78–89. [Google Scholar]
- Frank, M.C.; Thomas, R. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar]
- Gough, C.M.; Flower, C.E.; Vogel, C.S.; Curtis, P.S. Phenological and Temperature Controls on the Temporal Non-Structural Carbohydrate Dynamics of Populus grandidentata and Quercus rubra. Forests 2010, 1, 65–81. [Google Scholar] [CrossRef]
- Zhou, L.; He, H.L.; Sun, X.M.; Zhang, L.; Yu, G.R.; Ren, X.L.; Wang, J.Y.; Zhang, J.H. Species- and Community-Scale Simulation of the Phenology of a Temperate Forest in Changbai Mountain Based on Digital Camera Images. J. Resour. Ecol. 2013, 4, 317–326. [Google Scholar]
- Michael, A.W.; Peter, E.T.; Steven, W.R. A continental phenology model for monitoring vegetation responses to interannual climatic variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar]
- Franz, W.B.; Alberte, B.; Kristin, B.; Daniel, D.; Wolfgang, L.; Jörg, S.; Stephen, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar]
- Valdez, H.M.; Andrade, J.L.; Jackson, P.C.; Rebolledo, V.M. Phenology of five tree species of a tropical dry forest in Yucatan, Mexico: effects of environmental and physiological factors. Plant Soil 2010, 329, 155–171. [Google Scholar] [CrossRef]
- Richardson, A.D.; Bailey, A.S.; Denny, E.G.; Martin, C.W.; Keefe, J.O. Phenology of a northern hardwood forest canopy. Glob. Chang. Biol. 2006, 12, 1174–1188. [Google Scholar] [CrossRef]
- De Medeiros, D.P.W.; Lopes, A.V.; Zickel, C.S. Phenology of woody species in tropical coastal vegetation, northeastern Brazil. Funct. Ecol. Plants 2007, 202, 513–520. [Google Scholar] [CrossRef]
- Yuan, W.P.; Zhou, G.S.; Wang, Y.H.; Han, X.; Wang, Y.S. Simulating phenological characteristics of two dominant grass species in a semi-arid steppe ecosystem. Ecol. Res. 2007, 22, 784–791. [Google Scholar] [CrossRef]
- Chang, Z.F.; Zhu, S.J.; Han, F.G.; Zhong, S.N. Differences in response of desert plants of different ecotypes to climate warming: a case study in Minqin, Northwest China. J. Arid Land 2012, 4, 140–150. [Google Scholar] [CrossRef]
- Wolfe, D.W.; Schwartz, M.D.; Lakso, A.N.; Otsuki, Y.; Pool, R.M.; Shaulis, N.J. Climate change and shifts in spring phenology of three horticultural woody perennials in northeastern USA. Int. J. Biometeorol. 2005, 49, 303–309. [Google Scholar] [CrossRef] [PubMed]
- McEwan, R.W.; Brecha, R.J.; Geiger, D.R.; John, G.P. Flowering phenology change and climate warming in southwestern Ohio. Plant Ecol. 2011, 212, 55–61. [Google Scholar] [CrossRef]
- Ma, T.; Zhou, C.H. Climate-associated changes in spring plant phenology in China. Int. J. Biometeorol. 2012, 56, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; He, Z.B.; Yang, J.J.; Chen, L.F.; Zhu, X. Detecting the effects of climate change on canopy phenology in coniferous forests in semi-arid mountain regions of China. Int. J. Remote Sens. 2014, 35, 6490–6507. [Google Scholar] [CrossRef]
- He, Z.B.; Du, J.; Zhao, W.Z.; Yang, J.J.; Chen, L.F.; Zhu, X.; Chang, X.X.; Liu, H. Assessing temperature sensitivity of subalpine shrub phenology in semi-arid mountain regions of China. Agric. For. Meteorol. 2015, 213, 42–52. [Google Scholar] [CrossRef]
- Li, C.H.; Pei, S.X.; Guo, Q.S.; Xin, X.B.; Hong, M. Phenological responses of three shrub species to climate change in Harbin, Heilongjiang province. J. Northeast For. Univ. 2011, 39, 58–61. (In Chinese) [Google Scholar]
- Pei, S.X.; Guo, Q.S.; Xin, X.B.; Hong, M.; Kang, Y. Research on plant phenological responses to climate change abroad. World For. Res. 2009, 22, 31–37. (In Chinese) [Google Scholar]
- Xie, B.N.; Qin, Z.F.; Wang, Y.; Chang, Q.R. Monitoring vegetation phenology and their response to climate change on Chinese Loess Plateau based on remote sensing. Trans. Chin. Soc. Agric. Eng. 2015, 31, 153–160. (In Chinese) [Google Scholar]
- Chen, X.Q.; Tan, Z.J.; Schwartz, M.D.; Xu, C.X. Determining the growing season of land vegetation on the basis of plant phenology and satellite data in Northern China. Int. J. Biometeorol. 2000, 44, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Oscar, G.; Juan, J.S. Long-term temporal changes of plant phenology in the Western Mediterranean. Glob. Chang. Biol. 2009, 15, 1930–1948. [Google Scholar]
- Gunderson, A.C.; Edwards, N.T.; Walker, A.V.; O’Hara, K.H.; Campion, C.M.; Hanson, P.J. Forest phenology and a warmer climate—Growing season extension in relation to climatic provenance. Glob. Chang. Biol. 2012, 18, 2008–2025. [Google Scholar] [CrossRef]
- Seyed, A.H.; Sasan, B.K.; Asadollah, M. Evaluating Beech Tree Phenology in a Deciduous Broadleaf Forest in Northern Iran Using Ground Observation and MODIS Images. J. Sustain. For. 2011, 30, 697–712. [Google Scholar]
- Menzel, A. Phenology: Its importance to the global change community. Clim. Chang. 2002, 54, 379–385. [Google Scholar] [CrossRef]
- Jolly, W.M.; Running, S.W. Effects of precipitation and soil water potential on drought deciduous phenology in the Kalahari. Glob. Chang. Biol. 2004, 10, 303–308. [Google Scholar] [CrossRef]
- Xie, S.C.; Sheng, C.Y.; Li, S.C. A phonological study on main tree species of montane humid evergreen broad-leaved forest in Ailao Mountains. Acta Ecol. Sin. 1997, 17, 53–55. (In Chinese) [Google Scholar]
- Gough, C.M.; Flower, C.E.; Vogel, C.S.; Dragoni, D.O.; Curtis, P.S. Whole ecosystem labile carbon production in a north temperate deciduous forest. Agric. For. Meteorol. 2009, 149, 1531–1540. [Google Scholar] [CrossRef]
- Zhao, Z.; Jia, W.X.; Zhang, Y.S.; Liu, Y.R.; Chen, J.H. Monitoring and change trends of phonological phase in the Qilian Mountains based on remote sensing. J. Desert Res. 2015, 35, 1388–1395. (In Chinese) [Google Scholar]
- Deng, S.F. Impacts of Climate Change on Vegetation in Qilian Mountains from 2000 to 2011; Lanzhou University: Lanzhou, China, 2013. [Google Scholar]
- Jiang, L. Research on Soil Genetic Characteristics and Taxonomic Classification of Typical Soil Types in the Xi-shui Forest Zone of the Qilian Mountains; Northwest Agricultural and Forestry University: Yangling, China, 2012. [Google Scholar]
- Chang, Z.F.; Zhao, M.; Han, F.G.; Zhong, S.N.; Li, F.M. Phenological characteristics of desert plant in Minqin Desert area. Sci. Silvae Sin. 2008, 44, 58–64. (In Chinese) [Google Scholar]
- Annie, D.A.; Sergio, R.; Tommaso, A.; Antonio, S. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar]
- Han, F.; Zhang, Q.; Buyantuev, A.; Niu, J.M.; Liu, P.T.; Li, X.H.; Kang, S.; Zhang, J.; Chang, C.M.; Li, Y.P. Effects of climate change on phenology and primary productivity in the desert steppe of Inner Mongolia. J. Arid Land 2015, 7, 251–263. [Google Scholar] [CrossRef]
- Peng, H.H.; Li, C.K.; Tang, Z.G.; Liang, J. Spatial patterns in the terrestrial vegetation phenology of Danjiangkou reservoir area and its relation with elevation. Resour. Environ. Yangtze Basin 2016, 25, 1626–1634. (In Chinese) [Google Scholar]
- Yin, Z.Y.; Li, M.; Zhang, Y.; Shao, X. Growth–climate relationships along an elevation gradient on a southeast-facing mountain slope in the semi-arid eastern Qaidam Basin, northeastern Tibetan Plateau. Trees 2016, 30, 1095–1109. [Google Scholar] [CrossRef]
- Crimmins, T.M.; Crimmins, M.A.; Bertelsen, C.D. Spring and summer patterns in flowering onset, duration, and constancy across a water-limited gradient. Am. J. Bot. 2013, 100, 1137–1147. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shrub Species | Soil Depth (cm) | Soil Types | Elevation (m) | Slope Degree (°) | Slope Aspect | Canopy Cover (%) |
|---|---|---|---|---|---|---|
| Caragana. tangutica | 60 | Chestnut soil | 2600 | 26 | SW | 50 |
| Berberisdiaphana | 50 | Chestnut soil | 2600 | 30 | W | 65 |
| Potentilla Fruticosa L. | 60 | Meadow soil | 2900 | 31 | E | 80 |
| CaraganaJubata (Pall.) Poir. | 50 | Meadow soil | 3300 | 38 | NE | 65 |
| Salix Gilashanica C. Wang et P. Y. Fu | 50 | Meadow soil | 3300 | 36 | NE | 75 |
| Shrub Species | Elevation (m) | Aspect | Shrub Height (cm) | Basal Diameter (cm) | West-East Crown Diameter (cm) | North-South Crown Diameter (cm) |
|---|---|---|---|---|---|---|
| C. tangutica | 2600 | Sunny | 180.2 ± 5.8 | 1.3 ± 0.0 | 178.7 ± 9.4 | 183.9 ± 9.2 |
| B. diaphana | 2600 | Semi-sunny | 154.8 ± 4.8 | 1.2 ± 0.1 | 144.0 ± 5.5 | 156.9 ± 5.4 |
| P. fruticosa | 2900 | Semi-sunny | 45.5 ± 1.8 | 0.5 ± 0.0 | 44.5 ± 3.3 | 42.3 ± 3.0 |
| C. jubata | 3300 | Semi-sunny | 92.3 ± 2.2 | 1.2 ± 0.3 | 75.4 ± 3.7 | 89.2 ± 3.3 |
| S. gilashanica | 3300 | Sunny | 119.4 ± 3.5 | 1.1 ± 0.3 | 92.3 ± 5.0 | 90.5 ± 4.3 |
| Shrub Species | Budding | Leaf Opening | Flowering | Fruit Ripening | Leaf Discoloration | Leaf Fall | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | Start | End | Start | End | Start | End | Start | End | Start | End | ||
| Date (Month-Day) | C. tangutica | 4–26 | 5–12 | 5–13 | 6–11 | 6–12 | 7–15 | 7–16 | 9–1 | 9–2 | 9–15 | 9–16 | 9–25 |
| B.diaphana | 4–28 | 5–5 | 5–6 | 5–18 | 5–19 | 6–28 | 6–29 | 8–25 | 8–26 | 9–15 | 9–16 | 9–24 | |
| P. fruticosa | 5–1 | 5–15 | 5–16 | 6–30 | 7–1 | 7–31 | 8–1 | 8–14 | 8–15 | 9–15 | 9–16 | 9–26 | |
| C. jubata | 5–5 | 5–21 | 5–22 | 7–1 | 7–2 | 8–16 | 8–17 | 8–25 | 8–26 | 9–1 | 9–2 | 9–15 | |
| S. gilashanica | 5–5 | 5–21 | 5–22 | 6–24 | 6–25 | 8–14 | 8–15 | 8–25 | 8–26 | 9–14 | 9–15 | 9–25 | |
| Duration (days) | C. tangutica | 17 | 30 | 34 | 48 | 14 | 10 | ||||||
| B. diaphana | 8 | 13 | 41 | 58 | 21 | 9 | |||||||
| P. fruticosa | 15 | 46 | 31 | 14 | 32 | 11 | |||||||
| C. jubata | 17 | 41 | 46 | 9 | 7 | 14 | |||||||
| S. gilashanica | 17 | 34 | 51 | 11 | 20 | 11 | |||||||
| Shrub Species | Tacc Phenophase | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Budding | Leaf Opening | Flowering | Fruit Ripening | Leaf Color Change | Leaf Fall | Growing Season | ||||||||
| Start | End | Start | End | Start | End | Start | End | Start | End | Start | End | |||
| Tacc at start and end (°C) | C. tangutica | 123.2 | 232.1 | 244.2 | 520.1 | 530.5 | 950.9 | 963.8 | 1659.8 | 1674.5 | 1781.1 | 1789.9 | 1858.6 | 1735.4 |
| B. diaphana | 144.7 | 195.0 | 202.3 | 299.2 | 312.3 | 729.7 | 742.6 | 1555.5 | 1570.3 | 1781.1 | 1789.9 | 1854.2 | 1709.5 | |
| P. fruticosa | 119.6 | 193.0 | 201.2 | 603.3 | 615.4 | 1000.5 | 1018.8 | 1187.0 | 1199.7 | 1494.9 | 1501.9 | 1559.3 | 1439.7 | |
| C. jubata | 75.5 | 151.3 | 154.6 | 423.7 | 434.5 | 916.5 | 919.8 | 986.4 | 997.2 | 1062.4 | 1073.0 | 1126.8 | 1051.3 | |
| S. gilashanica | 75.5 | 151.3 | 154.6 | 351.0 | 359.3 | 893.2 | 903.6 | 986.4 | 997.2 | 1120.5 | 1126.8 | 1163.7 | 1088.2 | |
| Tacc for the duration of phenophase (°C) | C. tangutica | 108.9 | 275.9 | 420.4 | 696.0 | 106.6 | 68.7 | - | ||||||
| B. diaphana | 50.3 | 96.9 | 417.4 | 812.9 | 210.8 | 64.3 | - | |||||||
| P. fruticosa | 73.4 | 402.1 | 385.1 | 168.2 | 295.2 | 57.4 | - | |||||||
| C. jubata | 75.8 | 269.1 | 482.0 | 66.6 | 65.2 | 53.8 | - | |||||||
| S. gilashanica | 75.8 | 196.4 | 533.9 | 82.8 | 123.3 | 36.9 | - | |||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Liu, X.; Li, G.; Wang, S.; Zhao, W.; Ma, J. Phenology of Five Shrub Communities along an Elevation Gradient in the Qilian Mountains, China. Forests 2018, 9, 58. https://doi.org/10.3390/f9020058
Zhao Y, Liu X, Li G, Wang S, Zhao W, Ma J. Phenology of Five Shrub Communities along an Elevation Gradient in the Qilian Mountains, China. Forests. 2018; 9(2):58. https://doi.org/10.3390/f9020058
Chicago/Turabian StyleZhao, Yonghong, Xiande Liu, Guang Li, Shunli Wang, Weijun Zhao, and Jian Ma. 2018. "Phenology of Five Shrub Communities along an Elevation Gradient in the Qilian Mountains, China" Forests 9, no. 2: 58. https://doi.org/10.3390/f9020058
APA StyleZhao, Y., Liu, X., Li, G., Wang, S., Zhao, W., & Ma, J. (2018). Phenology of Five Shrub Communities along an Elevation Gradient in the Qilian Mountains, China. Forests, 9(2), 58. https://doi.org/10.3390/f9020058