Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Data Collection
2.2. Area Occupied by Whitebark Pine
2.3. Number of Trees by Size Class and Forest Type
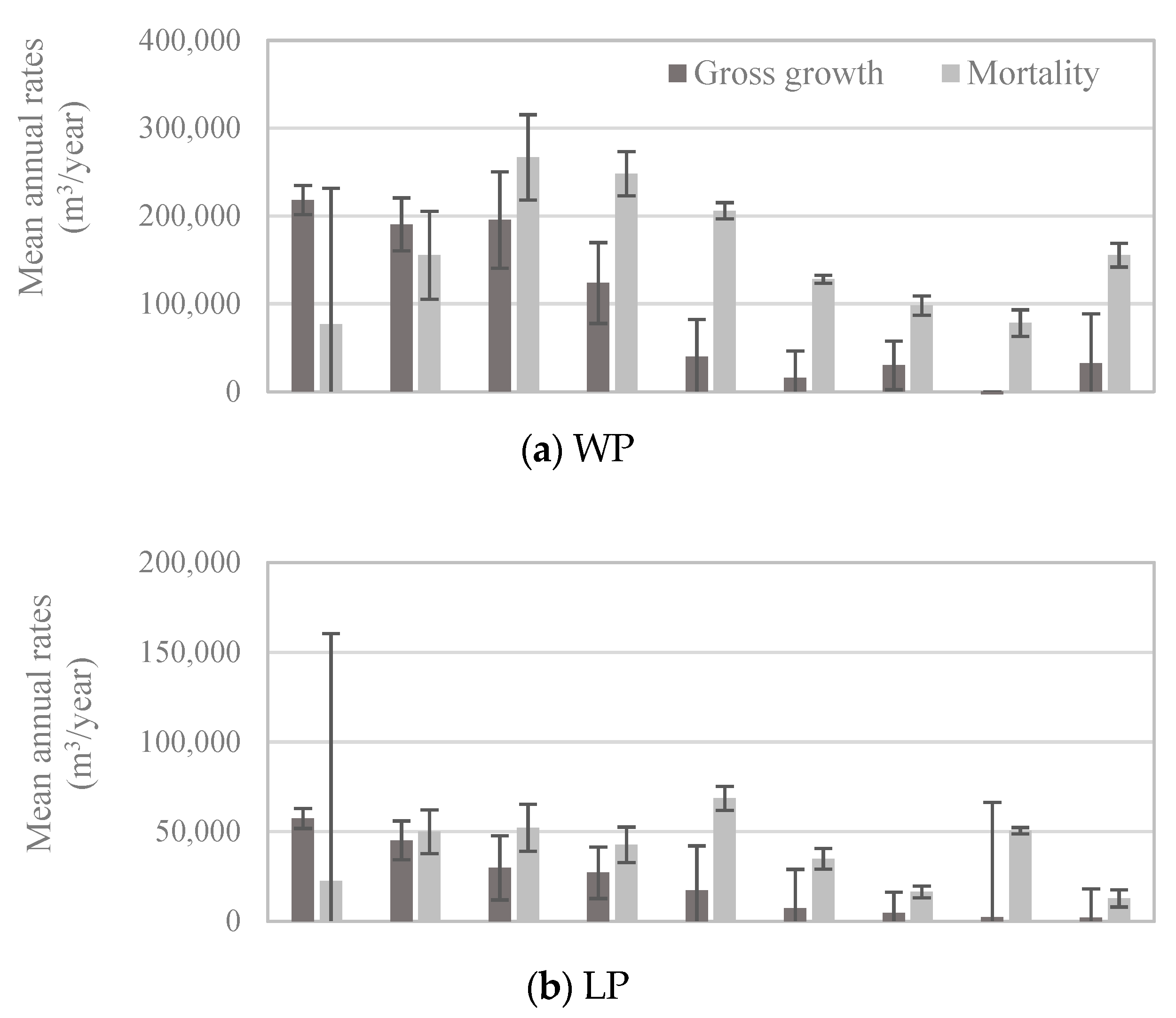
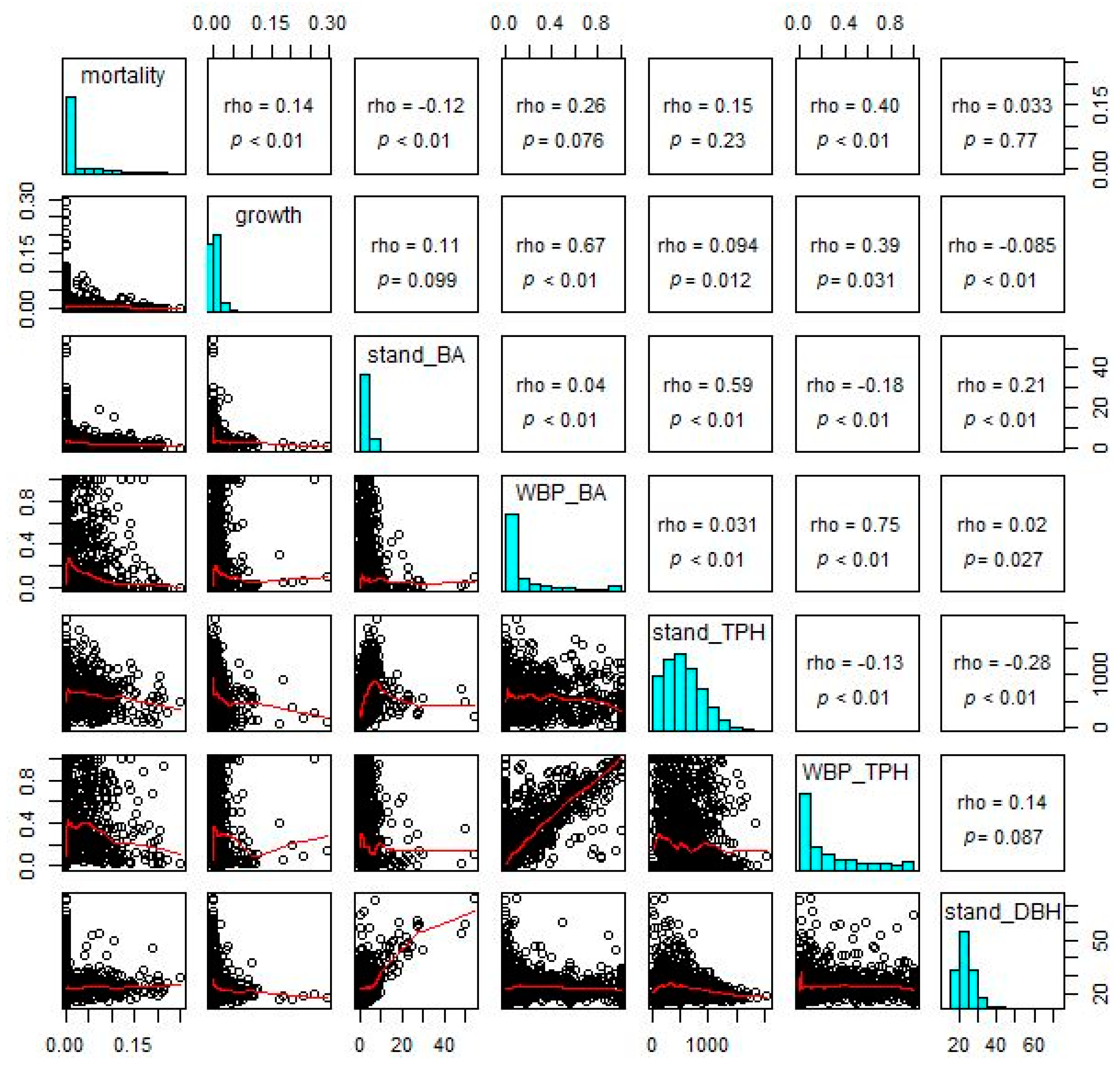
2.4. Whitebark Pine Growth and Mortality
3. Results
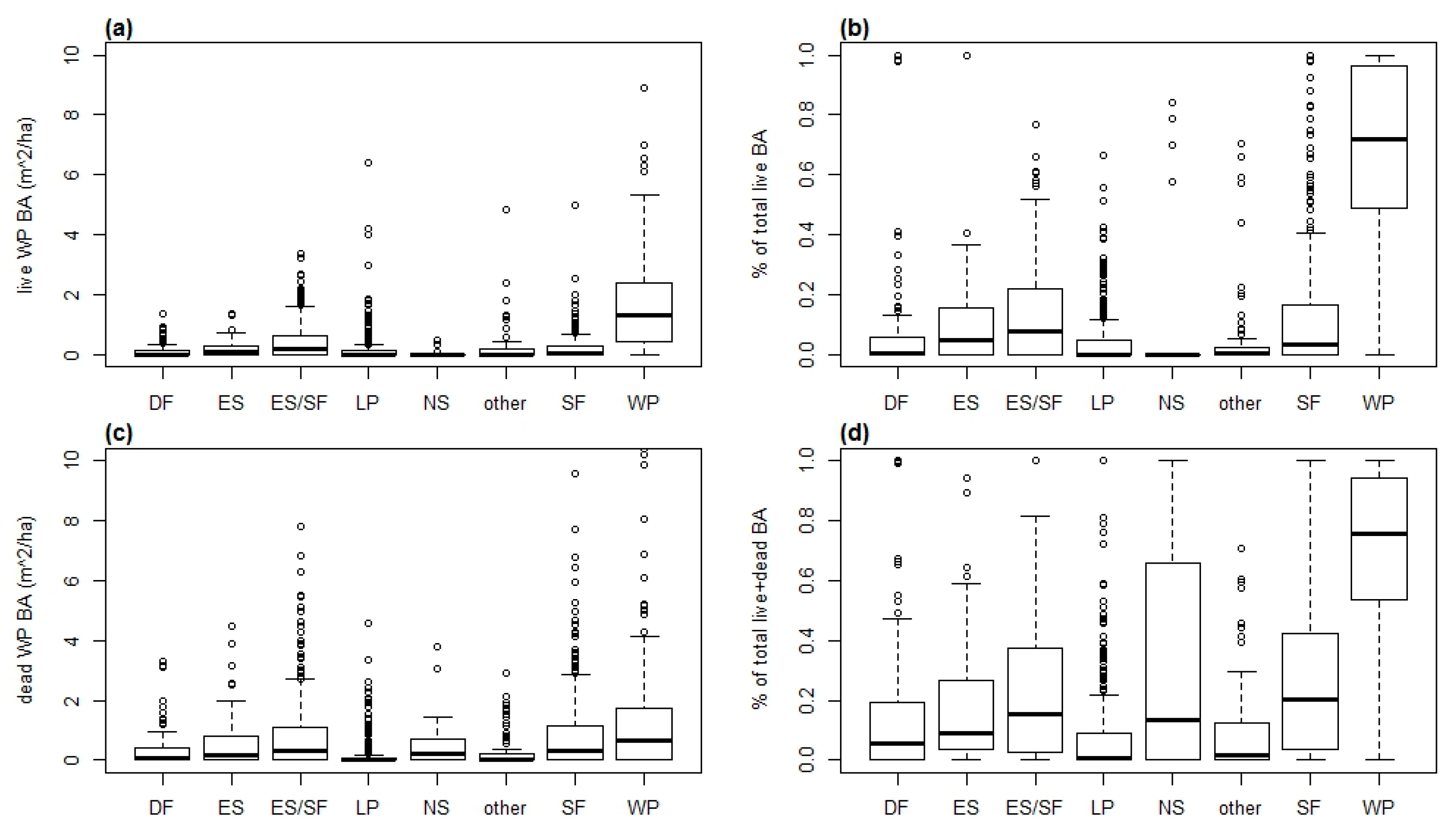
3.1. Area Occupied by Whitebark Pine
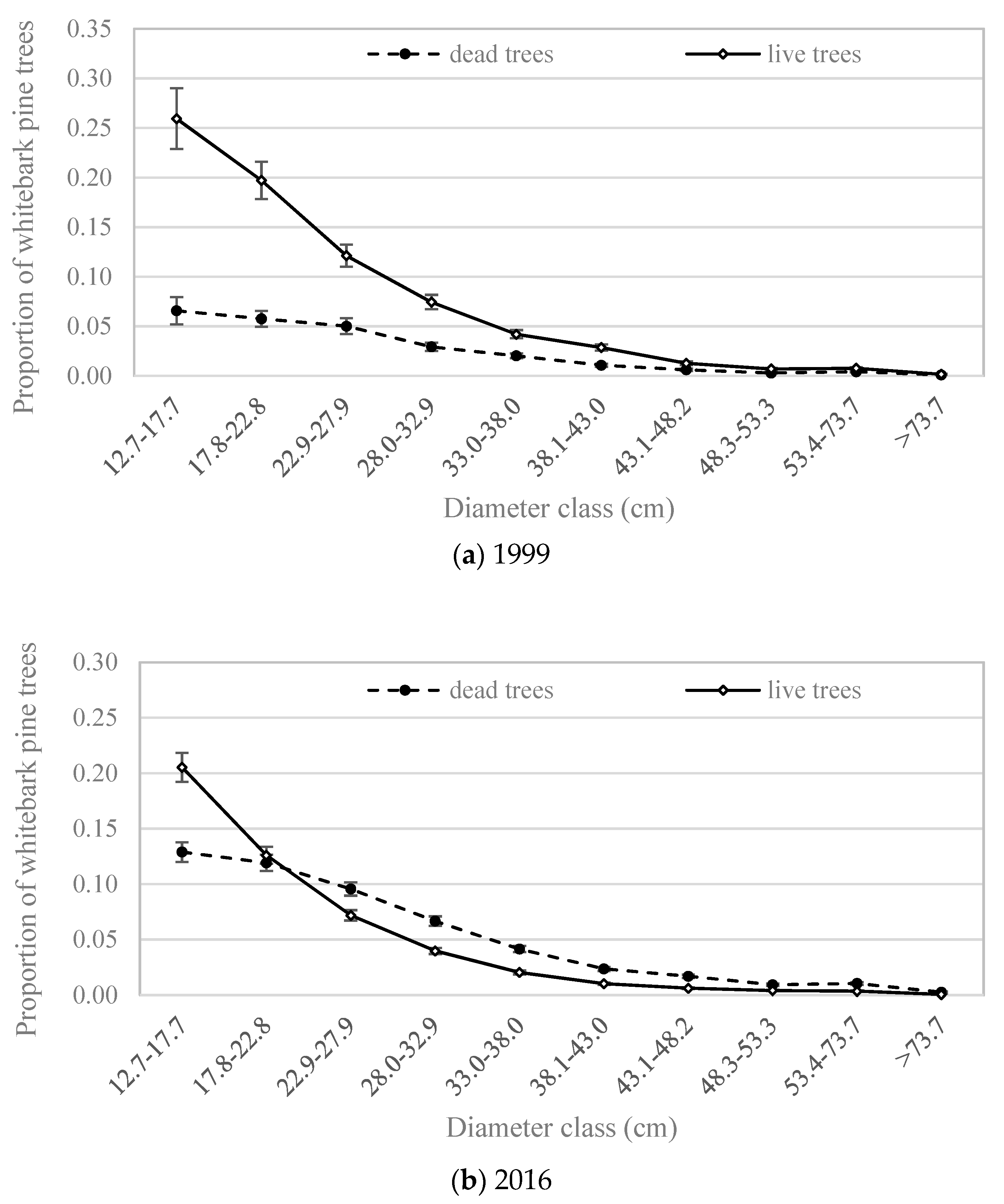
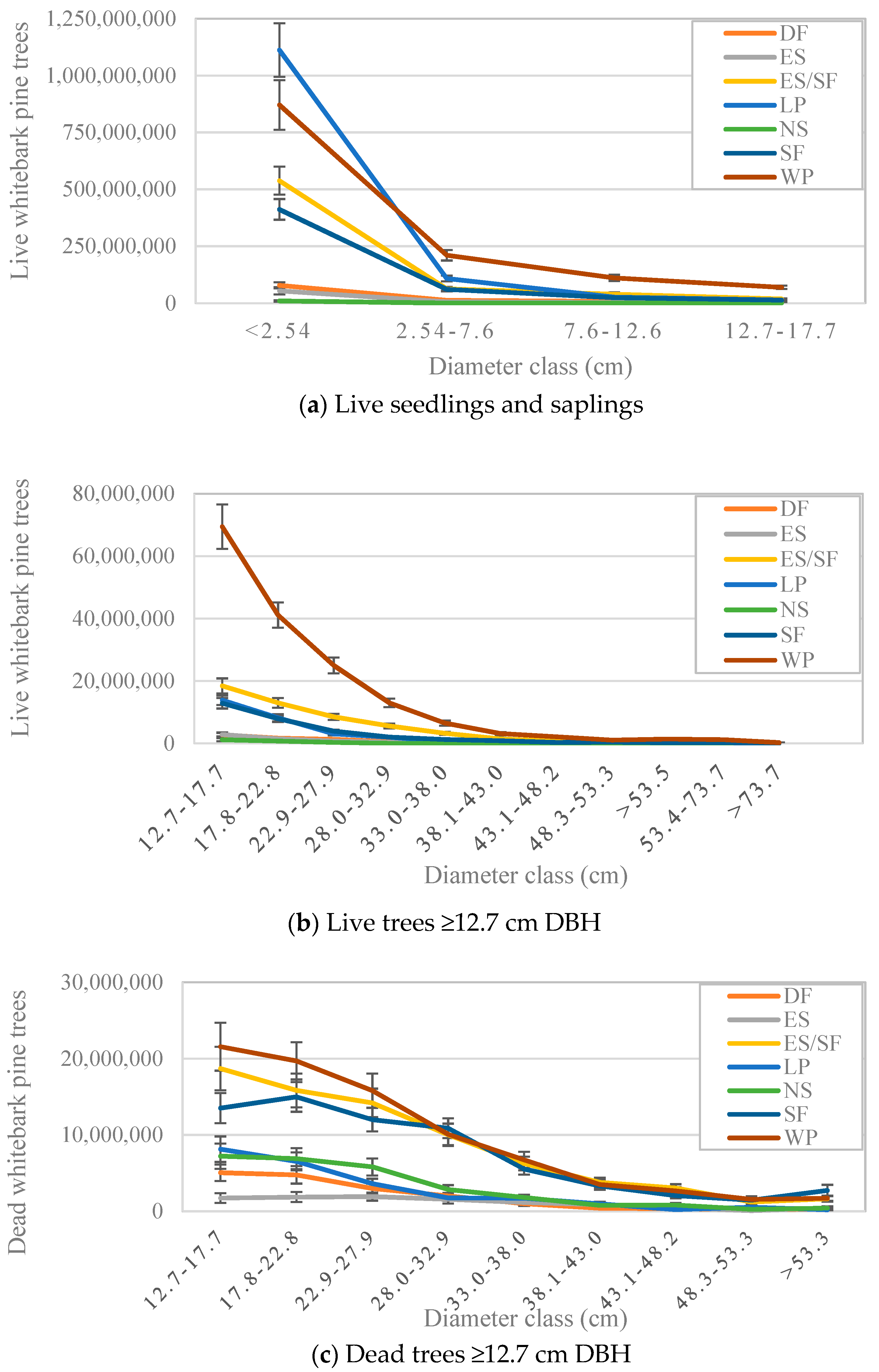
3.2. Number of Trees by Size Class and Forest Type
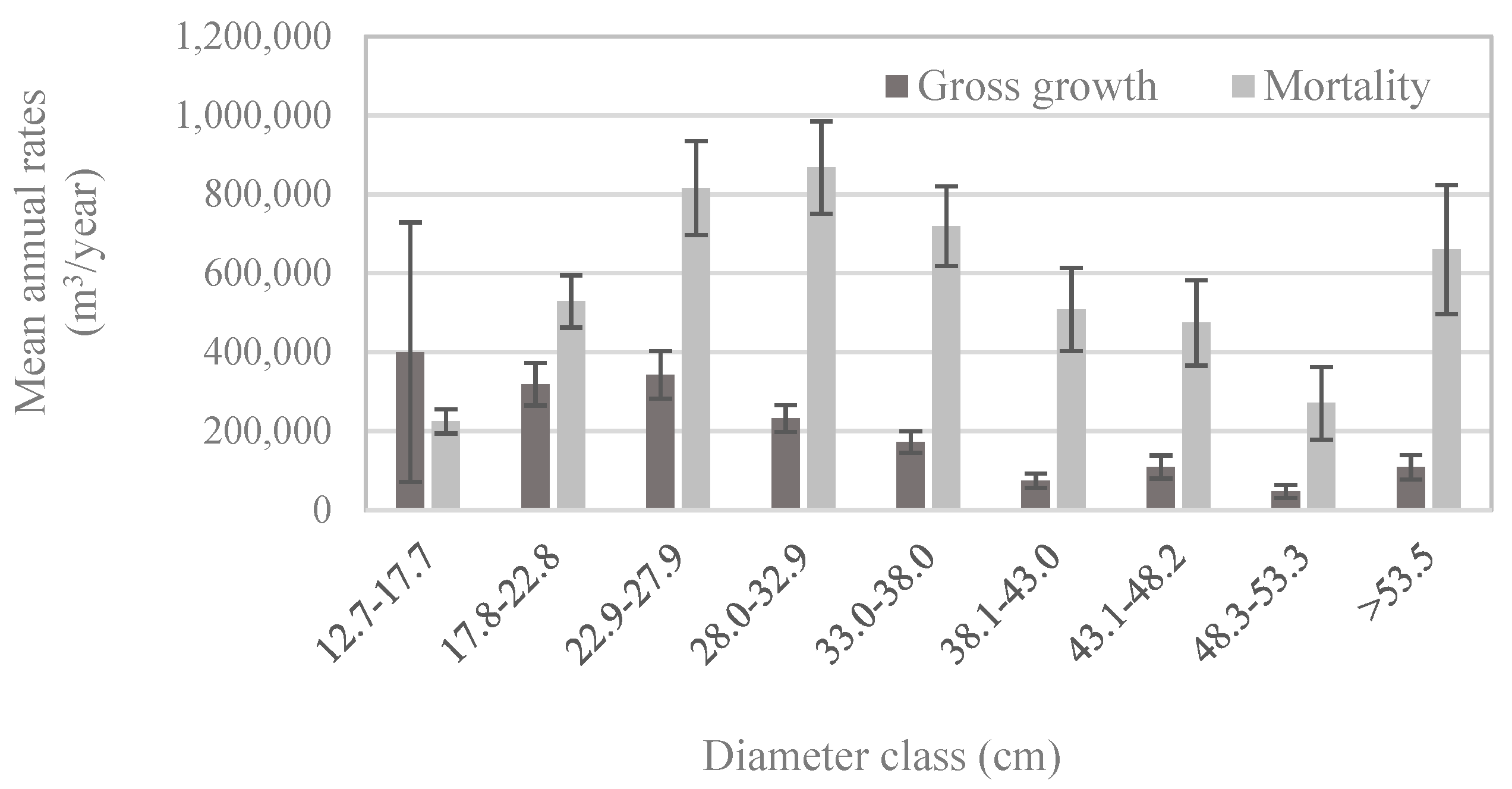
3.3. Whitebark Pine Growth and Mortality
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Forest, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of Foundation Species: Consequences for the Structure and Dynamics of Forested Ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Buotte, P.C.; Hicke, J.A.; Preisler, H.K.; Abatzoglou, J.T.; Raffa, K.F.; Logan, J.A. Climate influences on whitebark pine mortality from mountain pine beetle in the Greater Yellowstone Ecosystem. Ecol. Appl. 2016, 26, 2507–2524. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, E.; Irvine, K.M.; Thoma, D.; Wilmoth, S.; Ray, A.; Legg, K.; Shovic, H. Whitebark pine mortality related to white pine blister rust, mountain pine beetle outbreak, and water availability. Ecosphere 2016, 7, e01610. [Google Scholar] [CrossRef]
- Millar, C.I.; Westfall, R.D.; Delany, D.L.; Bokach, M.J.; Flint, A.L.; Flint, L.E. Forest mortality in high-elevation whitebark pine (Pinus albicaulis) forests of eastern California, USA; influence of environmental context, bark beetles, climatic water deficit, and warming. Can. J. For. Res. 2012, 42, 749–765. [Google Scholar] [CrossRef]
- Keane, R.E.; Tomback, D.F.; Aubry, C.A.; Bower, A.D.; Campbell, E.M.; Cripps, C.L.; Jenkins, M.B.; Mahalovich, M.F.; Manning, M.; McKinney, S.T.; et al. A Range-Wide Restoration Strategy for Whitebark Pine (Pinus albicaulis); RMRS-GTR-279; USDA Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; p. 108.
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Smith, C.M.; Wilson, B.; Rasheed, S.; Walker, R.C.; Carolin, T.; Shepherd, B. Whitebark pine and white pine blister rust in the Rocky Mountains of Canada and northern Montana. Can. J. For. Res. 2008, 38, 982–995. [Google Scholar] [CrossRef]
- Keane, R.E.; Arno, S.F. Rapid decline of whitebark pine in western Montana: Evidence from 20-year remeasurements. West. J. Appl. For. 1993, 8, 44–47. [Google Scholar]
- Tomback, D.F.; Resler, L.M.; Keane, R.E.; Pansing, E.R.; Andrade, A.J.; Wagner, A.C. Community structure, biodiversity, and ecosystem services in treeline whitebark pine communities: Potential impacts from a non-native pathogen. Forests 2016, 7, 21. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Endangered and threatened wildlife and plants: 12-month finding on a petition to list Pinus albicaulis as endangered or threatened with critical habitat. Fed. Regist. 2011, 76, 42631–42654. [Google Scholar]
- Retzlaff, M.L.; Leirfallom, S.B.; Keane, R.E. A 20-Year Reassessment of the Health and Status of Whitebark Pine Forests in the Bob Marshall Wilderness Complex, Montana; RMRS-RN-73; USDA Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2016; p. 10.
- Macfarlane, W.W.; Logan, J.A.; Kern, W.R. An innovative aerial assessment of Greater Yellowstone Ecosystem mountain pine beetle-caused whitebark pine mortality. Ecol. Appl. 2012, 23, 421–437. [Google Scholar] [CrossRef]
- Wong, C.M.; Daniels, L.D. Novel forest decline triggered by multiple interactions among climate, an introduced pathogen and bark beetles. Glob. Chang. Biol. 2017, 23, 1926–1941. [Google Scholar] [CrossRef] [PubMed]
- Larson, E.R.; Kipfmueller, K.F. Patterns in whitebark pine regeneration and their relationships to biophysical site characteristics in southwest Montana, central Idaho, and Oregon, USA. Can. J. For. Res. 2010, 40, 476–487. [Google Scholar] [CrossRef]
- Manley, P.N.; Zielinski, W.J.; Schlesinger, M.D.; Mori, S.R. Evaluation of a multiple-species approach to monitoring species at the ecoregional scale. Ecol. Appl. 2004, 14, 296–310. [Google Scholar] [CrossRef]
- Rogers, P.C.; O’Connell, B.; Mwang’ombe, J.; Madoffe, S.; Hertel, G. sForest health monitoring in the Ngangao forest, Taita Hills, Kenya: A five year assessment of change. J. East Afr. Nat. Hist. 2008, 97, 3–17. [Google Scholar] [CrossRef]
- O’Connell, B.M.; Conkling, B.M.; Wilson, A.M.; Burrill, E.A.; Turner, J.A.; Pugh, S.A.; Christiansen, G.; Ridley, T.; Menlove, J. The Forest Inventory and Analysis Database: Database Description and User Guide Version 7.0 for Phase 2. Available online: https://www.fia.fs.fed.us/library/database-documentation/ (accessed on 3 January 2018).
- Forest Inventory and Analysis National Core Field Guide, Volume I: Field Data Collection Procedures for Phase 2 Plots, Version 6.0. Available online: https://www.fia.fs.fed.us/library/field-guides-methods-proc/docs/2013/Core%20FIA%20P2%20field%20guide_6-0_6_27_2013.pdf (accessed on 3 January 2018).
- Cleland, D.T.; Freeouf, J.A.; Keys, J.E.; Nowacki, G.J.; Carpenter, C.A.; McNab, W.H. Ecological Subregions: Sections and Subsections for the Conterminous United States; WO-GTR-76D; USDA Forest Service: Washington, DC, USA, 2007.
- Reams, G.A.; Smith, W.D.; Hansen, M.H.; Bechtold, W.A.; Roesch, F.A.; Moisen, G.G. The forest inventory and analysis sampling frame. In The Enhanced Forest Inventory and Analysis Program—National Sampling Design and Estimation Procedures; Bechtold, W.A., Patterson, P.L., Eds.; FS-SRS-80; USDA Forest Service Southern Research Station: Asheville, NC, USA, 2005; pp. 11–26. [Google Scholar]
- FIA DataMart. Available online: https://apps.fs.usda.gov/fia/datamart/ (accessed on 3 January 2018).
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1996; p. 662. [Google Scholar]
- Miles, P.D. Forest Inventory EVALIDator Web-Application Version 1.6.0.03. Available online: http://apps.fs.fed.us/Evalidator/evalidator.jsp (accessed on 23 March 2017).
- Goeking, S.A. Disentangling forest change from forest inventory change: A case study from the US Interior West. J. Forest. 2015, 113, 475–483. [Google Scholar] [CrossRef]
- Zar, J.H. Significance Testing of the Spearman Rank Correlation Coefficient. J. Am. Stat. Assoc. 1972, 67, 578–580. [Google Scholar] [CrossRef]
- Cleveland, W.S.; Devlin, S.J. Locally weighted regression: An approach to regression analysis by local fitting. J. Am. Stat. Assoc. 1988, 83, 596–610. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 20 February 2018).
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: New York, NY, USA, 2013; 426p. [Google Scholar]
- Beardmore, T.; Loo, J.; McAfee, B.; Malouin, C.; Simpson, D. A survey of tree species of concern in Canada: The role for genetic conservation. For. Chron. 2006, 82, 351–363. [Google Scholar] [CrossRef]
- Hutchins, H.E.; Lanner, R.M. The central role of Clark’s nutcracker in the dispersal and establishment of Whitebark pine. Oecologia 1982, 55, 192–201. [Google Scholar] [CrossRef] [PubMed]
- McKinney, S.T.; Fiedler, C.E.; Tomback, D.F. Invasive pathogen threatens bird–pine mutualism: Implications for sustaining a high-elevation ecosystem. Ecol. Appl. 2009, 19, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, T.J.; Sullivan, K.A.; Bakian, A.V.; Aubry, C.A. Cache-site selection in Clark’s nutcracker (Nucifraga columbiana). Auk 2011, 128, 237–247. [Google Scholar] [CrossRef]
- Leirfallom, S.B.; Keane, R.E.; Tomback, D.F.; Dobrowski, S.Z. The effects of seed source health on whitebark pine (Pinus albicaulis) regeneration density after wildfire. Can. J. For. Res. 2015, 45, 1597–1606. [Google Scholar] [CrossRef]
- Tomback, D.F.; Sund, S.K.; Hoffman, L.A. Postfire regeneration of Pinus albicaulis: Height–age relationships, age structure, and microsite characteristics. Can. J. For. Res. 1993, 23, 113–119. [Google Scholar] [CrossRef]
- Perkins, J.L. Fire enhances whitebark pine seedling establishment, survival, and growth. Fire Ecol. 2015, 11, 84–99. [Google Scholar] [CrossRef]
- Mahalovich, M.F.; Hipkins, V.D. Molecular genetic variation in whitebark pine (Pinus albicaulis Engelm.) in the Inland West. In The Future of High-Elevation, Five-Needle white Pines in Western North America: Proceedings of the High Five Symposium; Keane, R.E., Tomback, D.F., Murray, M.P., Smith, C.M., Eds.; Proceedings RMRS-P-63; USDA Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; pp. 118–132. [Google Scholar]
- Kinloch, B.B., Jr. White pine blister rust in North America: Past and prognosis. Phytopathology 2003, 93, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.M.; Antos, J.A. Postfire succession in Pinus albicualis—Abies lasiocarpa forests of southern British Columbia. Can. J. Bot. 2003, 81, 383–397. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Scale | Units | Source |
|---|---|---|---|
| Tree species | Tree | -- | SPCD in table TREE |
| Tree diameter | Tree | cm | DIA in table TREE |
| Tree live/dead status | Tree | -- | STATUS_CD in table TREE |
| Forest type | Stand | -- | FORTYPCD in table COND |
| Mean tree diameter | Stand | cm | Calc: f (tree diameter) |
| Total basal area of all species | Stand | m2 ha−1 | Calc: f (tree diameter) |
| Whitebark pine basal area/total basal area | Stand | % | Calc: f (tree species, tree diameter) |
| Total stem density of all species | Stand | trees∙ha−1 | Calc: f (TPA_UNADJ in table TREE) |
| Whitebark pine stem density/total stem density | Stand | % | Calc: f (tree species, TPA_UNADJ in table TREE) |
| Mean annual mortality | Stand | m3 ha−1 year−1 | Calc: f (FMORTCFAL, TPAMORT_UNADJ in table TREE) |
| Mean annual annual net growth | Stand | m3 ha−1 year−1 | Calc: f (FGROWCFAL, TPAGROW_UNADJ in table TREE) |
| Mean annual gross growth | Stand | m3 ha−1 year−1 | Calc: f (mean annual net growth, mean annual mortality) |
| Standardized mortality | Stand | % year−1 | Calc: f (mean annual mortality, VOLCFNET in table TREE) |
| Standarized growth | Stand | % year−1 | Calc: f (mean annual gross growth, VOLCFNET in table TREE) |
| Eco-province | Plot | -- | Calc: f (ECOSUBCD in table PLOT) |
| Forest Type | Area (ha) | CA | ID | MT | NV | OR | WA | WY |
|---|---|---|---|---|---|---|---|---|
| Douglas-fir | 323,790 | 0.00 | 0.52 | 0.31 | 0.00 | 0.00 | 0.07 | 0.09 |
| Engelmann spruce | 140,083 | 0.00 | 0.08 | 0.25 | 0.00 | 0.01 | 0.04 | 0.62 |
| Engelmann spruce/subalpine fir | 840,285 | 0.00 | 0.16 | 0.51 | 0.00 | 0.02 | 0.03 | 0.28 |
| Lodgepole pine | 952,007 | 0.06 | 0.17 | 0.42 | 0.00 | 0.05 | 0.02 | 0.29 |
| Nonstocked | 232,906 | 0.00 | 0.41 | 0.42 | 0.00 | 0.01 | 0.04 | 0.12 |
| Subalpine fir | 804,461 | 0.00 | 0.37 | 0.38 | 0.00 | 0.03 | 0.07 | 0.15 |
| Whitebark pine | 629,401 | 0.16 | 0.13 | 0.38 | 0.01 | 0.03 | 0.04 | 0.26 |
| All types covering < 100,000 ha | 205,645 | 0.10 | 0.06 | 0.18 | 0.02 | 0.26 | 0.27 | 0.10 |
| Total | 4,128,578 | 0.04 | 0.23 | 0.40 | 0.00 | 0.04 | 0.05 | 0.23 |
| Forest Type 1 | Seedlings 2 | Saplings 3 | Live Trees 4 | Dead Trees 4 |
|---|---|---|---|---|
| DF | 78,083,238 | 26,874,601 | 6,688,873 | 17,058,244 |
| ES | 54,130,800 | 15,069,681 | 5,766,980 | 10,112,510 |
| ES/SF | 537,944,279 | 153,556,743 | 51,739,482 | 74,710,866 |
| LP | 1,111,981,547 | 162,479,371 | 28,096,009 | 23,707,011 |
| NS | 9,489,166 | 2,188,641 | 2,188,642 | 26,884,162 |
| SF | 411,773,363 | 113,610,271 | 29,361,035 | 66,516,853 |
| WP | 870,792,088 | 483,766,807 | 162,446,784 | 83,322,974 |
| All types | 3,134,662,520 | 975,173,510 | 291,634,115 | 307,623,262 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goeking, S.A.; Izlar, D.K. Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline. Forests 2018, 9, 131. https://doi.org/10.3390/f9030131
Goeking SA, Izlar DK. Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline. Forests. 2018; 9(3):131. https://doi.org/10.3390/f9030131
Chicago/Turabian StyleGoeking, Sara A., and Deborah Kay Izlar. 2018. "Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline" Forests 9, no. 3: 131. https://doi.org/10.3390/f9030131
APA StyleGoeking, S. A., & Izlar, D. K. (2018). Pinus albicaulis Engelm. (Whitebark Pine) in Mixed-Species Stands throughout Its US Range: Broad-Scale Indicators of Extent and Recent Decline. Forests, 9(3), 131. https://doi.org/10.3390/f9030131