Phytophthora cinnamomi Colonized Reclaimed Surface Mined Sites in Eastern Kentucky: Implications for the Restoration of Susceptible Species




Abstract
:1. Introduction
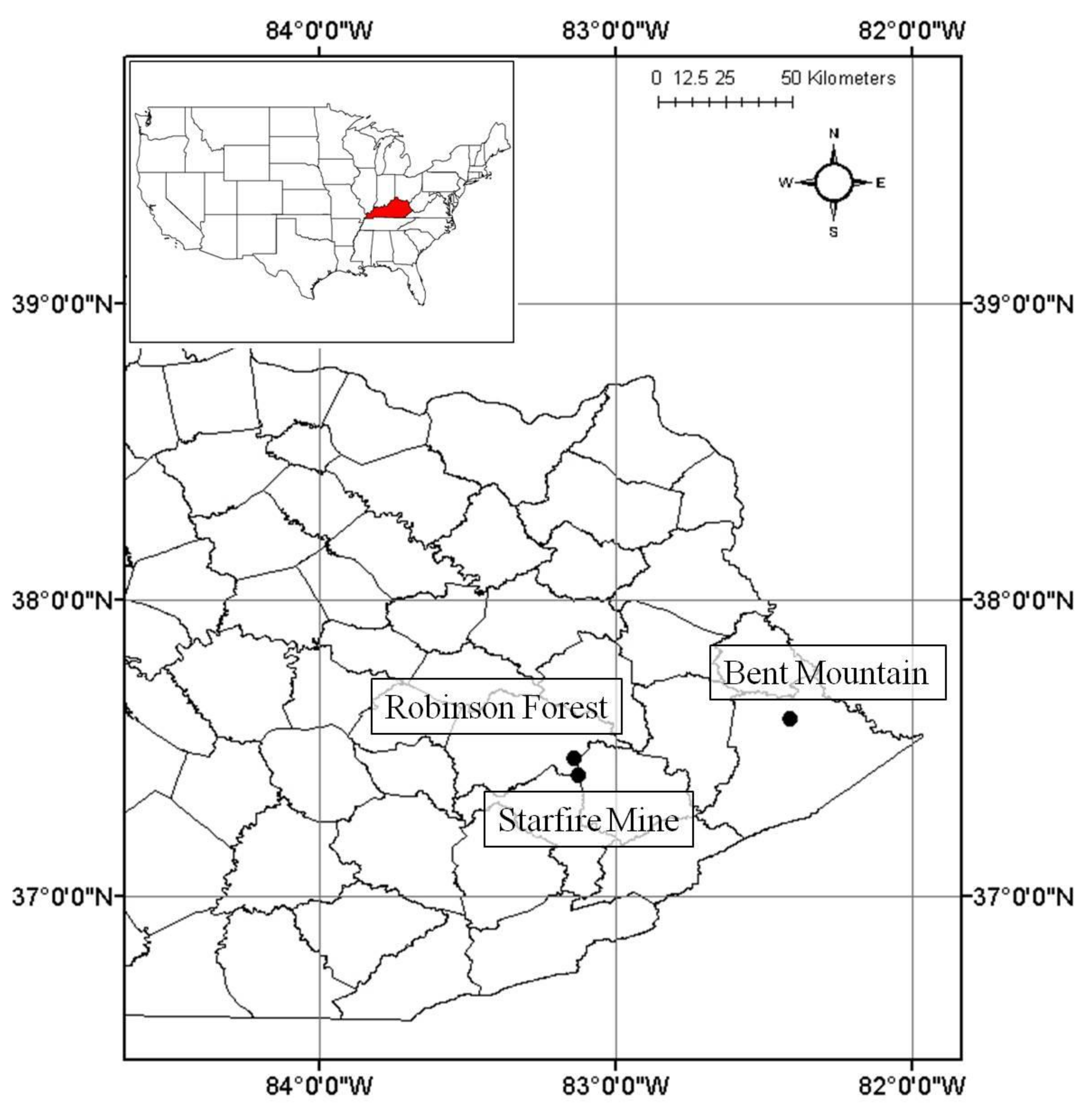
2. Methods and Materials
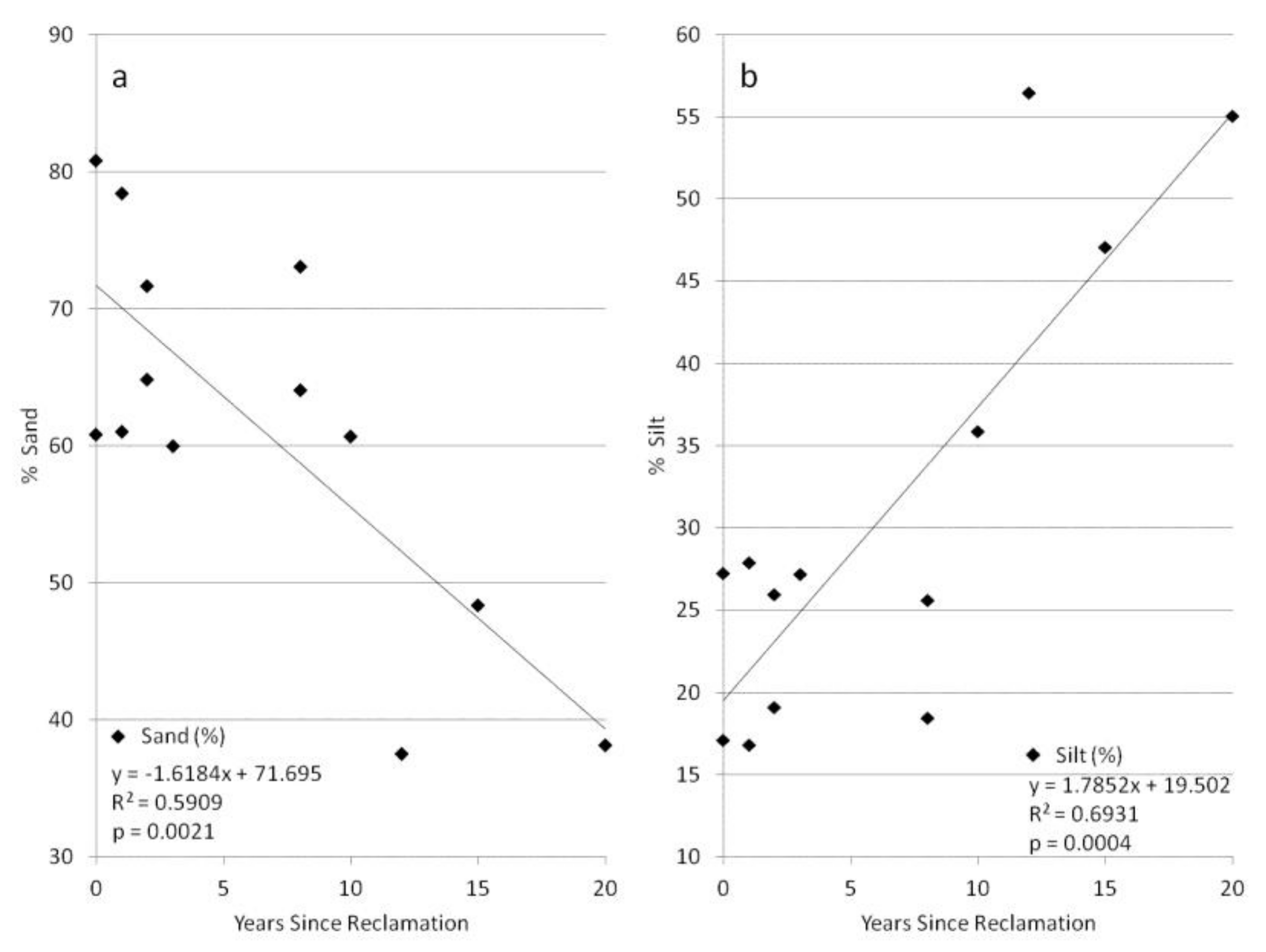
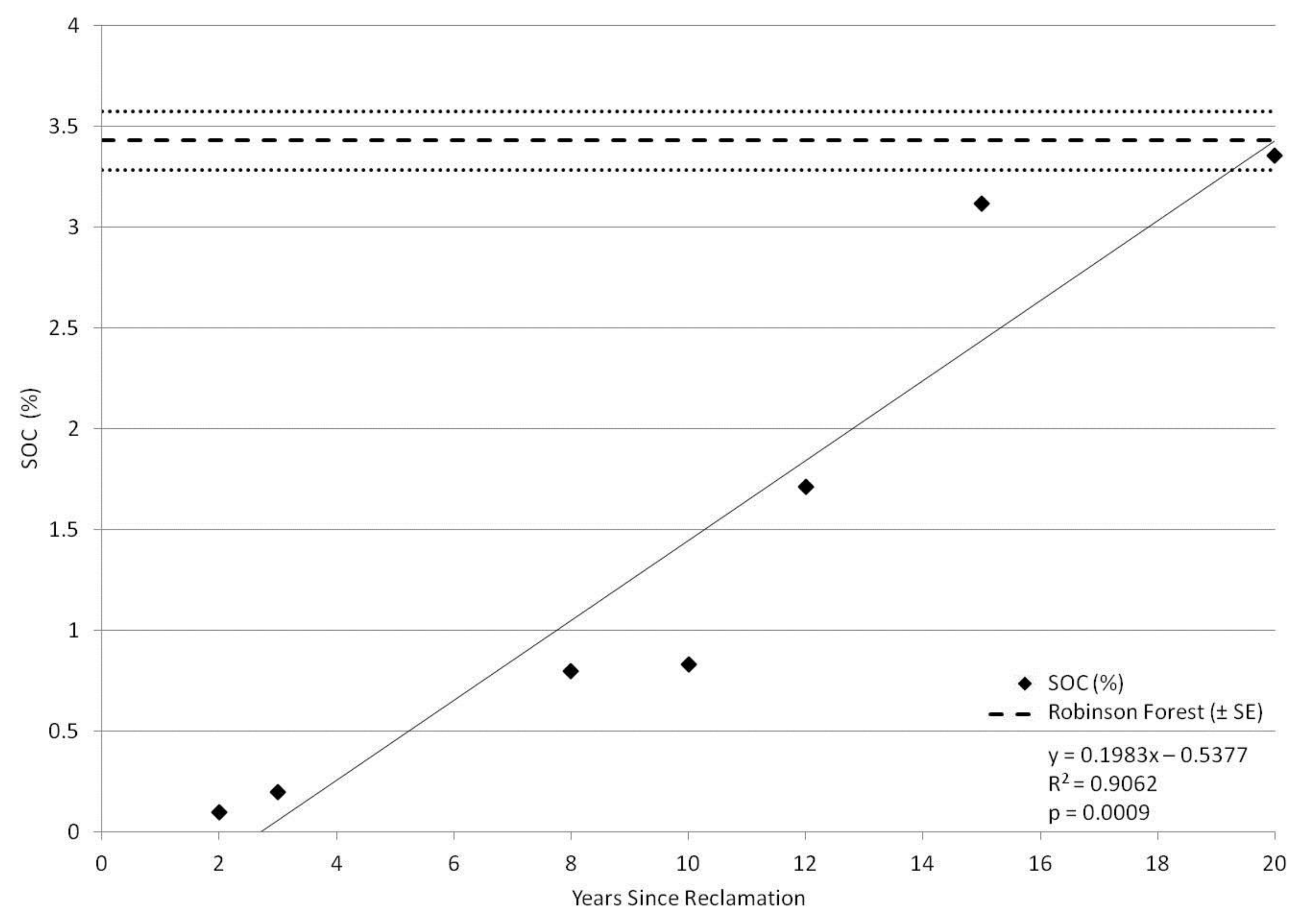
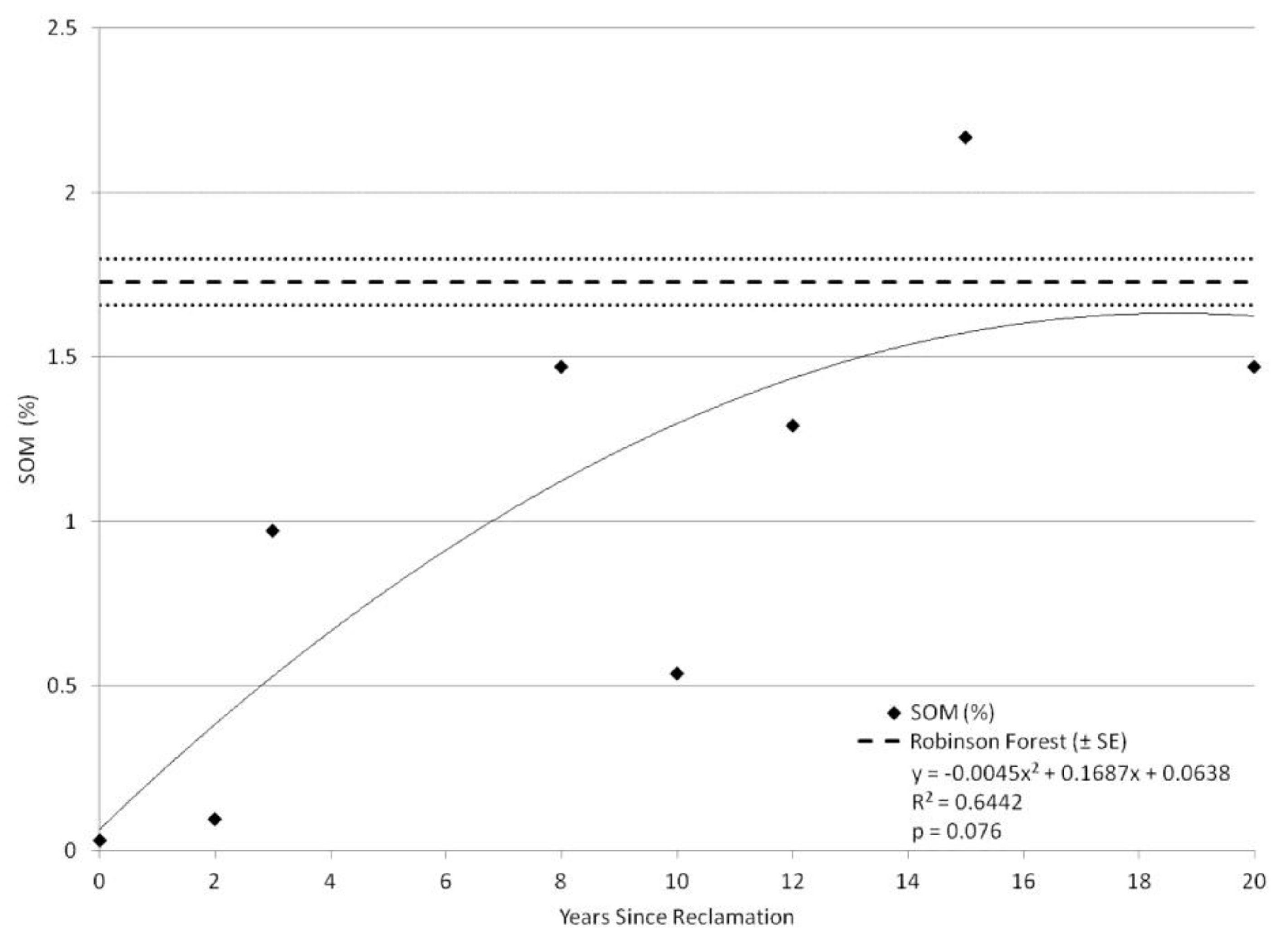
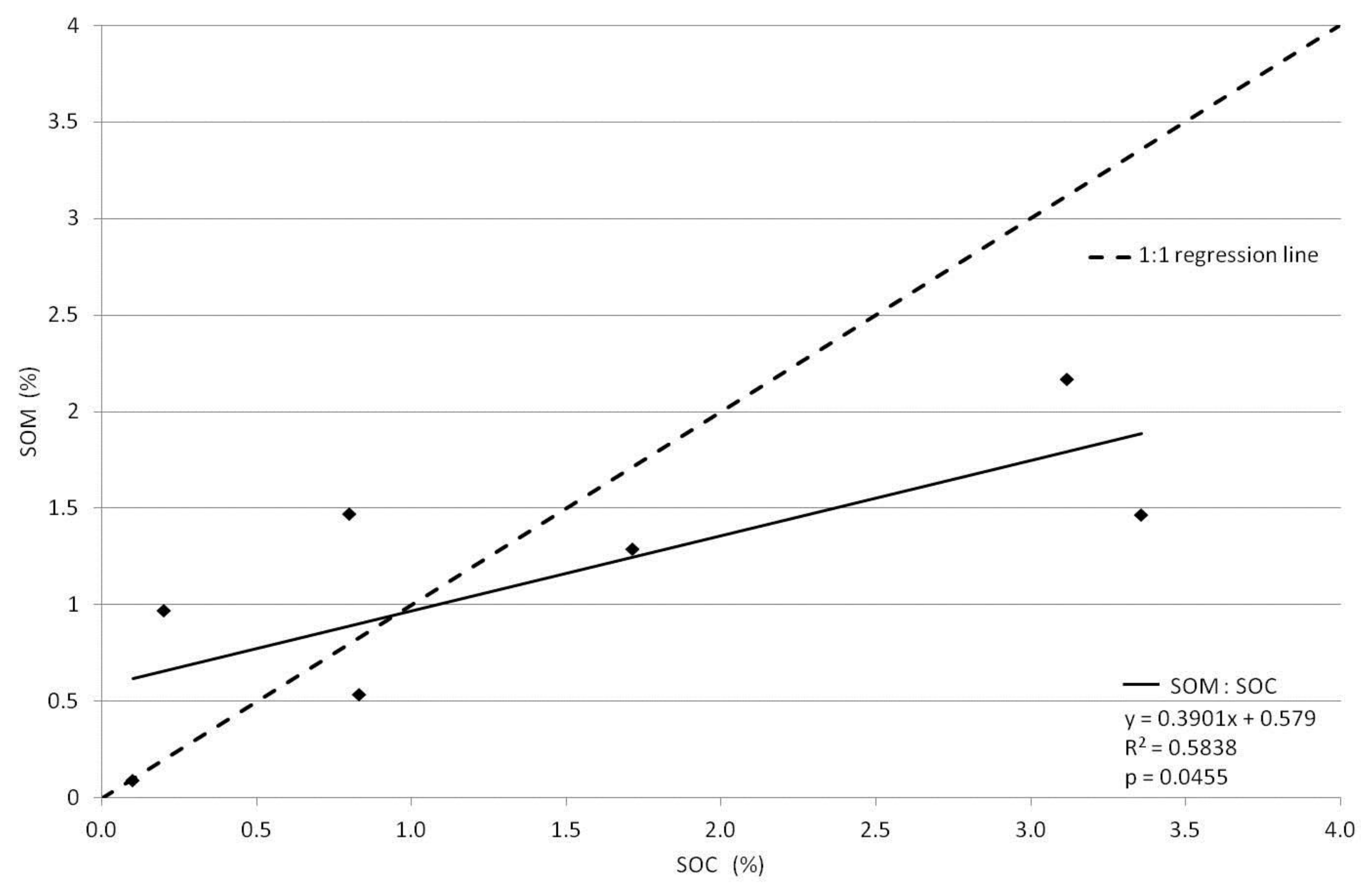
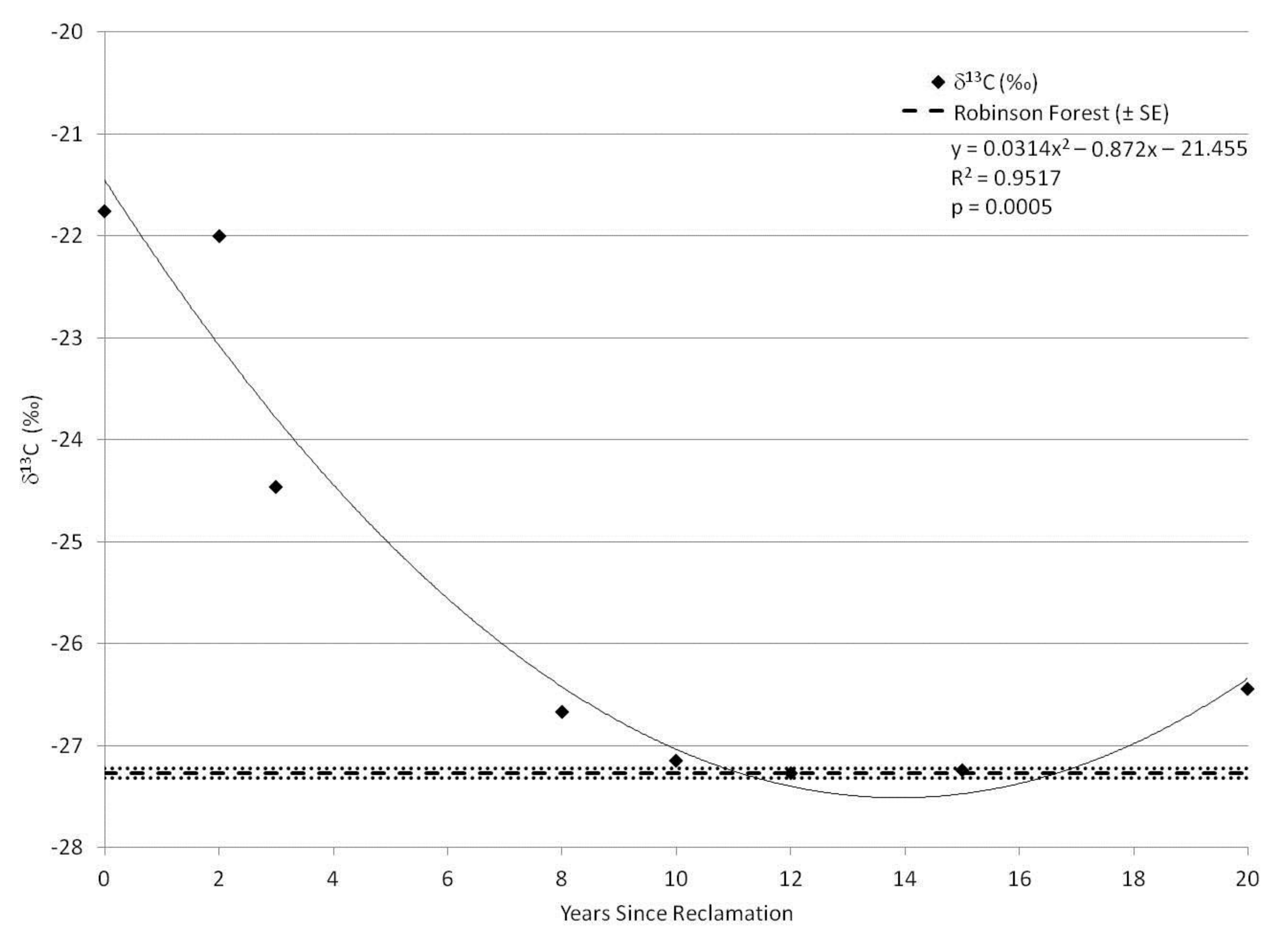
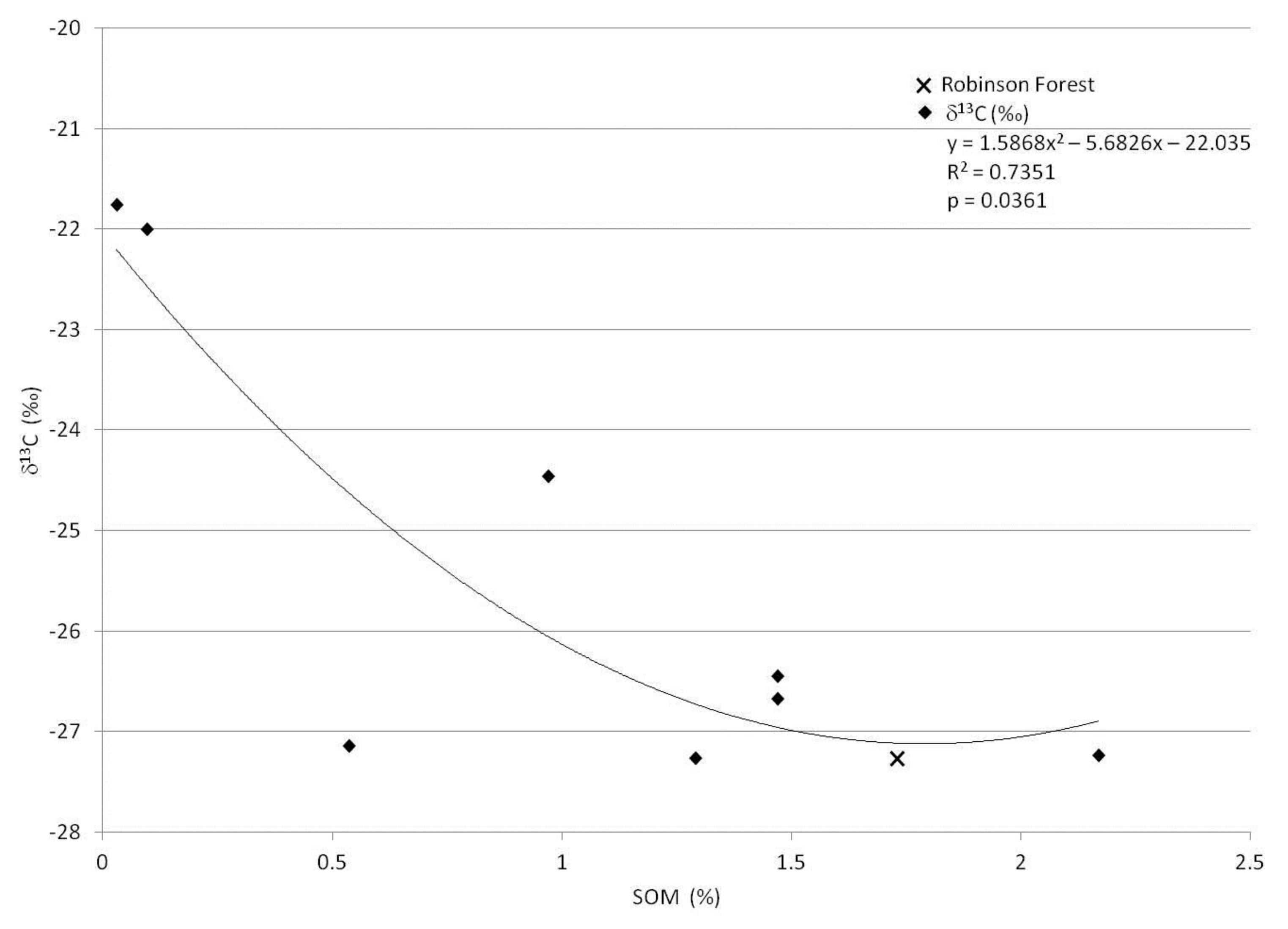
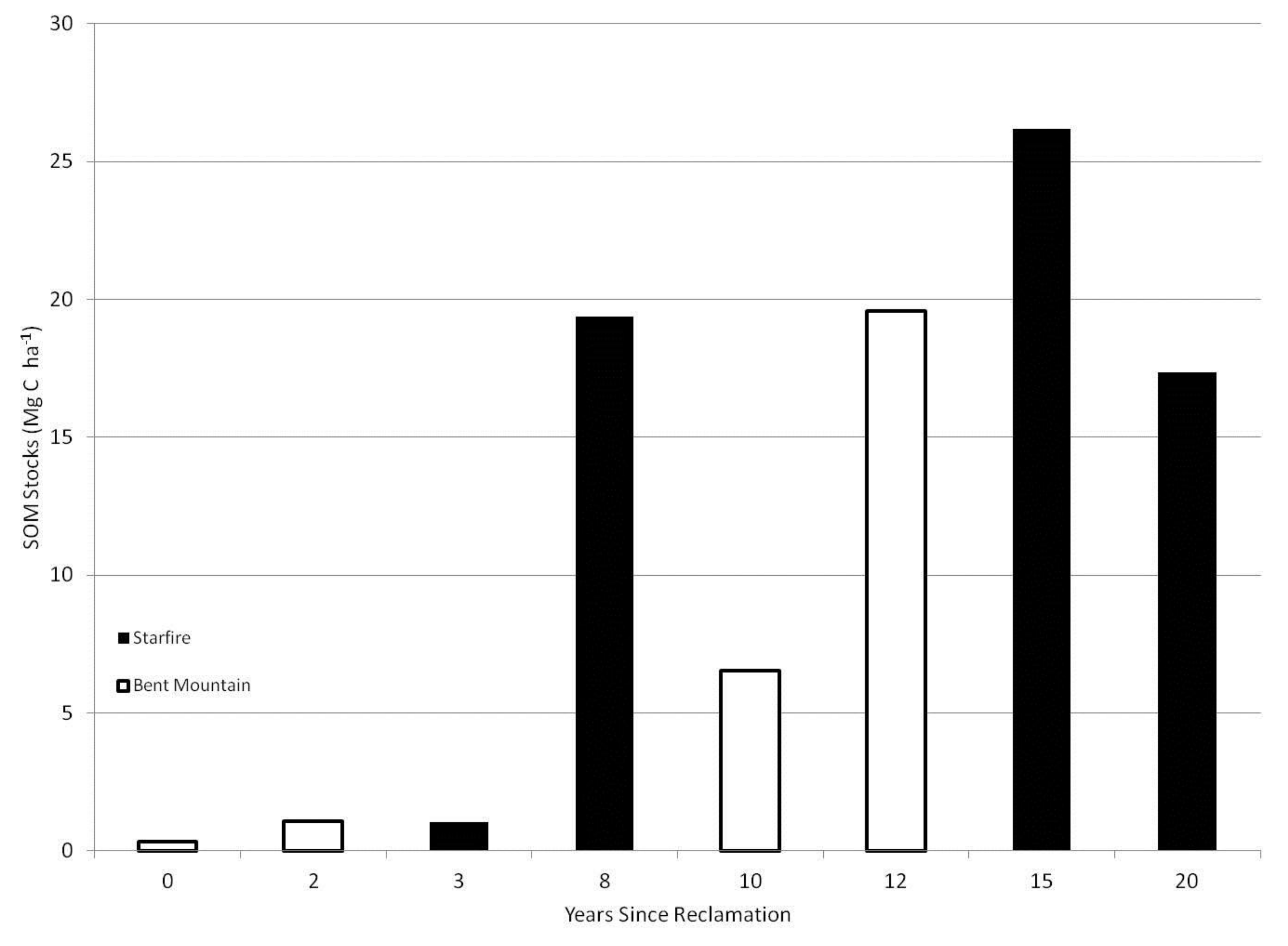
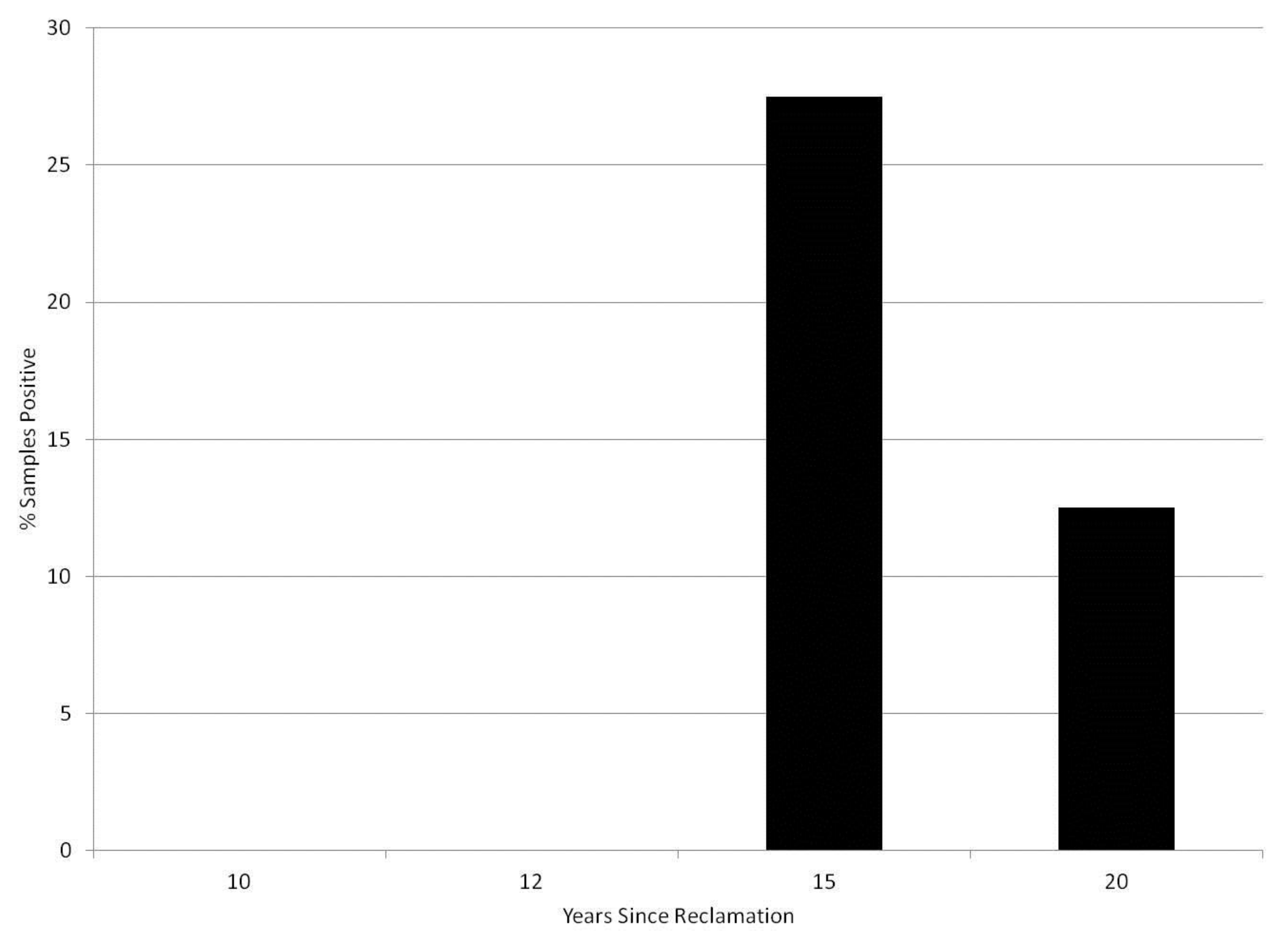
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Butler, P.R.; Iverson, L.; Thompson, F.R.; Brandt, L.; Handler, S.; Janowiak, M.; Shannon, P.D.; Swanston, C.; Karriker, K.; Bartig, J. Central Appalachians Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Central Appalachians Climate Change Response Framework Project; USDA Forest Service Northern Research Station, NRS-GTR-146; US Department of Agriculture: Washington, DC, USA, 2015.
- McLaughlin, S.B.; Wullschleger, S.D.; Sun, G.; Nosal, M. Interactive effects of ozone and climate on water use, soil moisture content and streamflow in a southern Appalachian forest in the USA. New Phytol. 2007, 174, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Wickham, J.; Wood, P.B.; Nicholson, M.C.; Jenkins, W.; Druckenbrod, D.; Suter, G.W.; Strager, M.P.; Mazzarella, C.; Galloway, W.; Amos, J. The overlooked terrestrial impacts of mountaintop mining. Bioscience 2013, 63, 335–348. [Google Scholar] [CrossRef]
- Gragson, T.L.; Bolstad, P.V. Land use legacies and the future of southern Appalachia. Soc. Nat. Resour. 2006, 19, 175–190. [Google Scholar] [CrossRef]
- Cormier, S.M.; Wilkes, S.P.; Zheng, L. Relationship of land use and elevated ionic strength in Appalachian watersheds. Environ. Toxicol. Chem. 2013, 32, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Flower, C.E.; Knight, K.S.; Rebbeck, J.; Gonzalez-Meler, M.A. The relationship between the emerald ash borer (Agrilus planipennis) and ash (Fraxinus spp.) tree decline: Using visual canopy condition assessments and leaf isotope measurements to assess pest damage. For. Ecol. Manag. 2013, 303, 143–147. [Google Scholar] [CrossRef]
- Anagnostakis, S. The effect of multiple importations of pests and pathogens on a native tree. Biol. Invasions 2001, 3, 245–254. [Google Scholar] [CrossRef]
- Nelson, C.D.; Powell, W.; Maynard, C.; Baier, K.; Newhouse, A.; Merkle, S.; Nairn, C.; Kong, L.; Carlson, J.; Addo-Quaye, C. The forest health initiative, American chestnut (Castanea dentata) as a model for forest tree restoration: Biological research program. In Fifth International Chestnut Symposium; Double, M.L., MacDonald, W.L., Eds.; International Society for Horticultural Science: Shepherdstown, WV, USA, 2012; pp. 179–189. [Google Scholar]
- Paillet, F.L. Chestnut: History and ecology of a transformed species. J. Biogeogr. 2002, 29, 1517–1530. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Diskin, M.; Steiner, K.C.; Hebard, F.V. Recovery of American chestnut characteristics following hybridization and backcross breeding to restore blight-ravaged Castanea dentata. For. Ecol. Manag. 2006, 223, 439–447. [Google Scholar] [CrossRef]
- Corsa, W.P. Nut Culture in the United States: Embracing Native and Introduced Species; USDA Division of Pomology: Washington, DC, USA, 1896.
- Milburn, M.; Gravatt, G. Preliminary note on a Phytophthora root disease of chestnut. Phytopathology 1932, 22, 977–978. [Google Scholar]
- Arentz, F. Phytophthora cinnamomi A1: An ancient resident of New Guinea and Australia of Gondwanan origin? For. Pathol. 2017, 47, e12342. [Google Scholar] [CrossRef]
- Shearer, B.L.; Crane, C.E.; Cochrane, A. Quantification of the susceptibility of the native flora of the south-west botanical province, Western Australia, to Phytophthora cinnamomi. Aust. J. Bot. 2004, 52, 435–443. [Google Scholar] [CrossRef]
- Jeffers, S.N.; James, J.B.; Sisco, P.H. Screening for resistance to Phytophthora cinnamomi in hybrid seedlings of American chestnut. In Phytophthoras in Forests and Natural Ecosystems: Proceedings of the Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party; Goheen, E.M., Frankel, S.J., Eds.; USDA Forest Service Pacific Southwest Research Station: Monterey, CA, USA, 2009; pp. 188–194. [Google Scholar]
- Rhoades, C.C.; Brosi, S.L.; Dattilo, A.J.; Vincelli, P. Effect of soil compaction and moisture on incidence of Phytophthora root rot on American chestnut (Castanea dentata) seedlings. For. Ecol. Manag. 2003, 184, 47–54. [Google Scholar] [CrossRef]
- Olukolu, B.A.; Nelson, C.D.; Abott, A. Mapping resistance to Phytophthora cinnamomi in chestnut (Castanea sp.). In Proceedings of the Fourth International Workshop on the Genetics of Host–Parasite Interactions in Forestry: Disease and Insect Resistance in Forest Trees, Eugene, OR, USA, 31 July–5 August 2012; Sniezko, R.A., Yanchuk, A.D., Kliejunas, J.T., Palmieri, K.M., Alexander, J.M., Frankel, S.J., Eds.; Pacific Southwest Research Station, Forest Service, US Department of Agriculture: Albany, CA, USA, 2012; p. 177. [Google Scholar]
- Zhebentyayeva, T.; Chandra, A.; Abbott, A.; Staton, M.; Olukolu, B.; Hebard, F.; Georgi, L.; Jeffers, S.; Sisco, P.; James, J. Genetic and genomic resources for mapping resistance to Phytophthora cinnamomi in chestnut. In Proceedings of the Fifth International Chestnut Symposium, Shepherdstown, WV, USA, 4–8 September 2013; Double, M.L., MacDonald, W.L., Eds.; International Society of Horticultural Science: Leuven, Belgium, 2013; pp. 263–270. [Google Scholar]
- Sena, K.; Crocker, E.; Vincelli, P.; Barton, C. Phytophthora cinnamomi as a driver of forest change: Implications for conservation and management. For. Ecol. Manag. 2018, 409, 799–807. [Google Scholar] [CrossRef]
- Meadows, I.M.; Jeffers, S.N. Distribution and recovery of Phytophthora cinnamomi in soils of mixed hardwood-pine forests of the south-eastern USA. N. Z. J. For. Sci. 2011, 41S, S39–S47. [Google Scholar]
- Wilson, B.; Lewis, A.; Aberton, J. Spatial model for predicting the presence of cinnamon fungus (Phytophthora cinnamomi) in sclerophyll vegetation communities in south-eastern Australia. Austral Ecol. 2003, 28, 108–115. [Google Scholar] [CrossRef]
- Pryce, J.; Edwards, W.; Gadek, P.A. Distribution of Phytophthora cinnamomi at different spatial scales: When can a negative result be considered positively? Austral Ecol. 2002, 27, 459–462. [Google Scholar] [CrossRef]
- McLaughlin, I.M.; Jeffers, S.N.; Waldrop, T.A. Survival of Phytophthora cinnamomi in soil after prescribed fire in a southern Appalachian mountain forest. In Phytophthoras in Forests and Natural Ecosystems: Proceedings of the Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party; Goheen, E.M., Frankel, S., Eds.; USDA Forest Service Pacific Southwest Research Station: Monterey, CA, USA, 2009; p. 151. [Google Scholar]
- Dawson, P.; Weste, G. Changes in the distribution of Phytophthora cinnamomi in the Brisbane Ranges National Park between 1970 and 1980–81. Aust. J. Bot. 1985, 33, 309–315. [Google Scholar] [CrossRef]
- Rodríguez-Molina, M.; Blanco-Santos, A.; Palo-Núñez, E.; Torres-Vila, L.; Torres-Álvarez, E.; Suárez-de-la-Cámara, M. Seasonal and spatial mortality patterns of holm oak seedlings in a reforested soil infected with Phytophthora cinnamomi. For. Pathol. 2005, 35, 411–422. [Google Scholar] [CrossRef]
- Shea, S.; Dell, B. Structure of the surface root system of Eucalyptus marginata Sm. and its infection by Phytophthora cinnamomi Rands. Aust. J. Bot. 1981, 29, 49–58. [Google Scholar] [CrossRef]
- Sena, K.; Dreaden, T.D.; Crocker, E.; Barton, C. Detection of Phytophthora cinnamomi in forest soils by PCR on DNA extracted from leaf disc baits. Plant Health Prog. 2018. accepted. [Google Scholar]
- Pinchot, C.C.; Schlarbaum, S.E.; Clark, S.L.; Saxton, A.M.; Sharp, A.M.; Schweitzer, C.J.; Hebard, F.V. Growth, survival, and competitive ability of chestnut (Castanea Mill.) seedlings planted across a gradient of light levels. New For. 2017, 48, 491–512. [Google Scholar] [CrossRef]
- Sharpe, S.R. Phytophthora Species Associated with American, Chinese, and Backcross Hybrid Chestnut Seedlings in Field Sites in the Southeastern United States. Master’s Thesis, Clemson University, Clemson, SC, USA, 2017. [Google Scholar]
- Zipper, C.E.; Burger, J.A.; Barton, C.D.; Skousen, J.G. Rebuilding soils on mined land for native forests in Appalachia. Soil Sci. Soc. Am. J. 2013, 77, 337–349. [Google Scholar] [CrossRef]
- Zipper, C.E.; Burger, J.A.; McGrath, J.M.; Rodrigue, J.A.; Holtzman, G.I. Forest restoration potentials of coal-mined lands in the eastern United States. J. Environ. Qual. 2011, 40, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Zipper, C.E.; Burger, J.A.; Skousen, J.G.; Angel, P.N.; Barton, C.D.; Davis, V.; Franklin, J.A. Restoring forests and associated ecosystem services on Appalachian coal surface mines. Environ. Manag. 2011, 47, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Amichev, B.Y.; Burger, J.A.; Rodrigue, J.A. Carbon sequestration by forests and soils on mined land in the midwestern and Appalachian coalfields of the U.S. For. Ecol. Manag. 2008, 256, 1949–1959. [Google Scholar] [CrossRef]
- Wickham, J.D.; Riitters, K.H.; Wade, T.G.; Coan, M.; Homer, C. The effect of Appalachian mountaintop mining on interior forest. Landsc. Ecol. 2006, 22, 179–187. [Google Scholar] [CrossRef]
- Adams, M.B. The Frestry Reclamation Approach: Guide to Successful Reforestation of Mined Lands; USDA Forest Service, N.R.S.: Newtown Square, PA, USA, 2017; Volume 169, p. 128.
- Wilson-Kokes, L.; Emerson, P.; DeLong, C.; Thomas, C.; Skousen, J. Hardwood tree growth after eight years on brown and gray mine soils in West Virginia. J. Environ. Qual. 2013, 42, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Kokes, L.; DeLong, C.; Thomas, C.; Emerson, P.; O’Dell, K.; Skousen, J. Hardwood tree growth on amended mine soils in West Virginia. J. Environ. Qual. 2013, 42, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Sena, K.; Barton, C.; Hall, S.; Angel, P.; Agouridis, C.; Warner, R. Influence of spoil type on afforestation success and natural vegetative recolonization on a surface coal mine in Appalachia, united states. Restor. Ecol. 2015, 23, 131–138. [Google Scholar] [CrossRef]
- Cotton, C.; Barton, C.; Lhotka, J.; Angel, P.N.; Graves, D. Evaluating reforestation success on a surface mine in eastern Kentucky. In National Proceedings: Forest and Conservation Nursery Associations; Haase, D.L., Pinto, J.R., Riley, L.E., Eds.; USDA Forest Service, Rocky Mountain Research Station, RMRS-P-68; Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; pp. 16–23. [Google Scholar]
- Chaudhuri, S.; Pena-Yewtukhiw, E.M.; McDonald, L.M.; Skousen, J.; Sperow, M. Early C sequestration rate changes for reclaimed minesoils. Soil Sci. 2012, 177, 443–450. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lal, R. Carbon and nitrogen pools in reclaimed land under forest and pasture ecosystems in Ohio, USA. Geoderma 2010, 157, 196–205. [Google Scholar] [CrossRef]
- French, M.E.; Barton, C.; Graves, D.; Angel, P.N.; Hebard, F.V. Evaluation of mine spoil suitability for the introduction of American chestnut hybrids in the Cumberland plateau. In Proceedings of the American Society of Mining and Reclamation, Gillette, WY, USA, 2–7 June 2007. [Google Scholar]
- Fields-Johnson, C.W.; Burger, J.A.; Evans, D.M.; Zipper, C.E. American chestnut establishment techniques on reclaimed Appalachian surface mined lands. Ecol. Restor. 2012, 30, 99–101. [Google Scholar] [CrossRef]
- McCarthy, B.C.; Bauman, J.M.; Keiffer, C.H. Mine land reclamation strategies for the restoration of American chestnut. Ecol. Restor. 2008, 26, 292–294. [Google Scholar] [CrossRef]
- Hiremath, S.; Lehtoma, K.; Bauman, J.M. Survey for the presence of Phytophthora cinnamomi on reclaimed mined lands in Ohio chosen for restoration of the American chestnut. J. Am. Soc. Min. Reclam. 2013, 2, 68–79. [Google Scholar] [CrossRef]
- Taylor, T.J.; Agouridis, C.T.; Warner, R.C.; Barton, C.D. Runoff curve numbers for loose-dumped spoil in the Cumberland plateau of eastern Kentucky. Int. J. Min. Reclam. Environ. 2009, 23, 103–120. [Google Scholar] [CrossRef]
- Taylor, T.J.; Agouridis, C.T.; Warner, R.C.; Barton, C.D.; Angel, P.N. Hydrologic characteristics of Appalachian loose-dumped spoil in the Cumberland plateau of eastern Kentucky. Hydrol. Process. 2009, 23, 3372–3381. [Google Scholar] [CrossRef]
- Sena, K.; Barton, C.; Angel, P.; Agouridis, C.; Warner, R. Influence of spoil type on chemistry and hydrology of interflow on a surface coal mine in the eastern US coalfield. Water Air Soil Pollut. 2014, 225, 1–14. [Google Scholar] [CrossRef]
- Adank, K.M.; Barton, C.D.; French, M.E.; de Sá, P.B. Occurrence of Phytophthora on reforested loose-graded spoils in eastern Kentucky. In Proceedings of the American Society of Mining and Reclamation, Richmond, VA, USA, 14–19 June 2008; pp. 1–13. [Google Scholar]
- Littlefield, T.; Barton, C.; Arthur, M.; Coyne, M. Factors controlling carbon distribution on reforested minelands and regenerating clearcuts in Appalachia, USA. Sci. Total Environ. 2013, 465, 240–247. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture (USDA). Forest Inventory and Analysis Factsheet, Kentucky; USDA: Washington, DC, USA, 2004; 4p.
- Cherry, M.A. Hydrochemical Characterization of Ten Headwater Catchments in Eastern Kentucky. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2006; 166p. [Google Scholar]
- Daniels, W.L.; Zipper, C.E.; Orndorff, Z.W.; Skousen, J.; Barton, C.D.; McDonald, L.M.; Beck, M.A. Predicting total dissolved solids release from central Appalachian coal mine spoils. Environ. Pollut. 2016, 216, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Agouridis, C.T.; Angel, P.N.; Taylor, T.J.; Barton, C.D.; Warner, R.C.; Yu, X.; Wood, C. Water quality characteristics of discharge from reforested loose-dumped mine spoil in eastern Kentucky. J. Environ. Qual. 2012, 41, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Barton, C.; Agouridis, C.; Fogel, A.; Dowdy, T.; Angel, P. Evaluating soil genesis and reforestation success on a surface coal mine in Appalachia. Soil Sci. Soc. Am. J. 2012, 76, 950–960. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal rbosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Schena, L.; Duncan, J.; Cooke, D. Development and application of a PCR-based ‘molecular tool box’ for the identification of Phytophthora species damaging forests and natural ecosystems. Plant Pathol. 2008, 57, 64–75. [Google Scholar] [CrossRef]
- Kunadiya, M.; White, D.; Dunstan, W.A.; Hardy, G.E.S.J.; Andjic, V.; Burgess, T.I. Pathways to false-positive diagnoses using molecular genetic detection methods; Phytophthora cinnamomi a case study. FEMS Microbiol. Lett. 2017, 364, fnx009. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, S.; Barton, C.D.; Karathanasis, T.A.; Rowe, H.D.; Rimmer, S.M. Distinguishing “new” from “old” organic carbon in reclaimed coal mine sites using thermogravimetry: II. Field validation. Soil Sci. 2007, 172, 302–312. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Hillier, S. Particulate composition and origin of suspended sediment in the R. Don, Aberdeeshire, UK. Sci. Total Environ. 2001, 265, 281–293. [Google Scholar] [CrossRef]
- Yeager, K.M.; Santschi, P.H.; Phillips, J.D.; Herbert, B.E. Suspended sediment sources and tributary effects in the lower reaches of a coastal plain stream as indicated by radionuclides, Loco Bayou, Texas. Environ. Geol. 2005, 3, 382–395. [Google Scholar] [CrossRef]
- Grossman, R.B.; Reinsch, T.G. The solid phase. In Methods of Soil Analysis, Part 4: Physical Methods; Dane, J.H., Topp, G.C., Eds.; Soil Science Society of America: Madison, WI, USA, 2002. [Google Scholar]
- Maharaj, S.; Barton, C.D.; Karathanasis, T.A.; Rowe, H.D.; Rimmer, S.M. Distinguishing “new” from “old” organic carbon on reclaimed coal mine sites using thermogravimetry: I. Method development. Soil Sci. 2007, 172, 292–301. [Google Scholar] [CrossRef]
- Essington, M. Soil and Water Chemistry: An Integrative Approach; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Sposito, G. The Chemistry of Soils; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- French, M.E. Establishment of American Chestnuts on Surface Mined Lands in the Appalachian Coalfields Region. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2017. [Google Scholar]
- Swiecki, T.J.; Bernhardt, E.A. Testing and implementing methods for managing Phytophthora root diseases in California native habitats and restoration sites. In Proceedings of the Sudden Oak Death Sixth Science Symposium, San Francisco, CA, USA, 20–23 June 2017. [Google Scholar]
- Balci, Y.; Long, R.; Mansfield, M.; Balser, D.; MacDonald, W. Involvement of Phytophthora species in white oak (Quercus alba) decline in southern Ohio. For. Pathol. 2010, 40, 430–442. [Google Scholar] [CrossRef]
- Hwang, S.; Ko, W. Biology of chlamydospores, sporangia, and zoospores of Phytophthora cinnamomi in soil. Phytopathology 1978, 68, 726–731. [Google Scholar] [CrossRef]
- Hardy, G.S.J.; Colquhoun, I.; Nielsen, P. The early development of disease caused by Phytophthora cinnamomi in Eucalyptus marginata and Eucalyptus calophylla growing in rehabilitated bauxite mined areas. Plant Pathol. 1996, 45, 944–954. [Google Scholar] [CrossRef]
- Koch, J.M.; Samsa, G.P. Restoring jarrah forest trees after bauxite mining in Western Australia. Restor. Ecol. 2007, 15, S17–S25. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years Since Reclamation | Robinson Forest | ||||
|---|---|---|---|---|---|
| 10 | 12 | 15 | 20 | ||
| Al (ppm) | 36,800 ± 640 | 36,800 ± 1200 | 46,600 ± 7200 | 48,100 ± 9800 | 50,800 ± 5000 |
| Mn (ppm) | 184 ± 8.5 | 316 ± 25 | 321 ± 38 | 259 ± 39 | 997 ± 150 |
| Fe (ppm) | 9770 ± 140 | 17,800 ± 960 | 17,400 ± 2500 | 19,800 ± 3400 | 25,600 ± 2000 |
| Mg (ppm) | 1420 ± 37 | 2280 ± 63 | 2700 ± 550 | 3300 ± 680 | 1980 ± 230 |
| K (ppm) | 13,500 ± 400 | 12,400 ± 800 | 15,400 ± 2500 | 14,600 ± 2700 | 12,900 ± 1400 |
| Ca (ppm) | 489 ± 36 | 1040 ± 130 | 1540 ± 440 | 1220 ± 62 | 455 ± 110 |
| Na (ppm) | 3880 ± 31 | 2740 ± 120 | 2650 ± 220 | 1800 ± 590 | 833 ± 57 |
| % Sand | 60.6 ± 3.3 | 37.5 ± 1.9 | 48.4 ± 5.9 | 38.1 ± 2.1 | 27.3 ± 3.7 |
| % Silt | 35.9 ± 3.0 | 56.4 ± 2.2 | 47.1 ± 5.8 | 55.1 ± 2.0 | 62.6 ± 3.6 |
| % Clay | 3.45 ± 0.38 | 6.07 ± 0.48 | 4.53 ± 0.33 | 6.80 ± 1.6 | 10.1 ± 0.28 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sena, K.L.; Yeager, K.M.; Dreaden, T.J.; Barton, C.D. Phytophthora cinnamomi Colonized Reclaimed Surface Mined Sites in Eastern Kentucky: Implications for the Restoration of Susceptible Species. Forests 2018, 9, 203. https://doi.org/10.3390/f9040203
Sena KL, Yeager KM, Dreaden TJ, Barton CD. Phytophthora cinnamomi Colonized Reclaimed Surface Mined Sites in Eastern Kentucky: Implications for the Restoration of Susceptible Species. Forests. 2018; 9(4):203. https://doi.org/10.3390/f9040203
Chicago/Turabian StyleSena, Kenton L., Kevin M. Yeager, Tyler J. Dreaden, and Christopher D. Barton. 2018. "Phytophthora cinnamomi Colonized Reclaimed Surface Mined Sites in Eastern Kentucky: Implications for the Restoration of Susceptible Species" Forests 9, no. 4: 203. https://doi.org/10.3390/f9040203
APA StyleSena, K. L., Yeager, K. M., Dreaden, T. J., & Barton, C. D. (2018). Phytophthora cinnamomi Colonized Reclaimed Surface Mined Sites in Eastern Kentucky: Implications for the Restoration of Susceptible Species. Forests, 9(4), 203. https://doi.org/10.3390/f9040203