Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests


Abstract
:1. Introduction
2. Materials and Methods
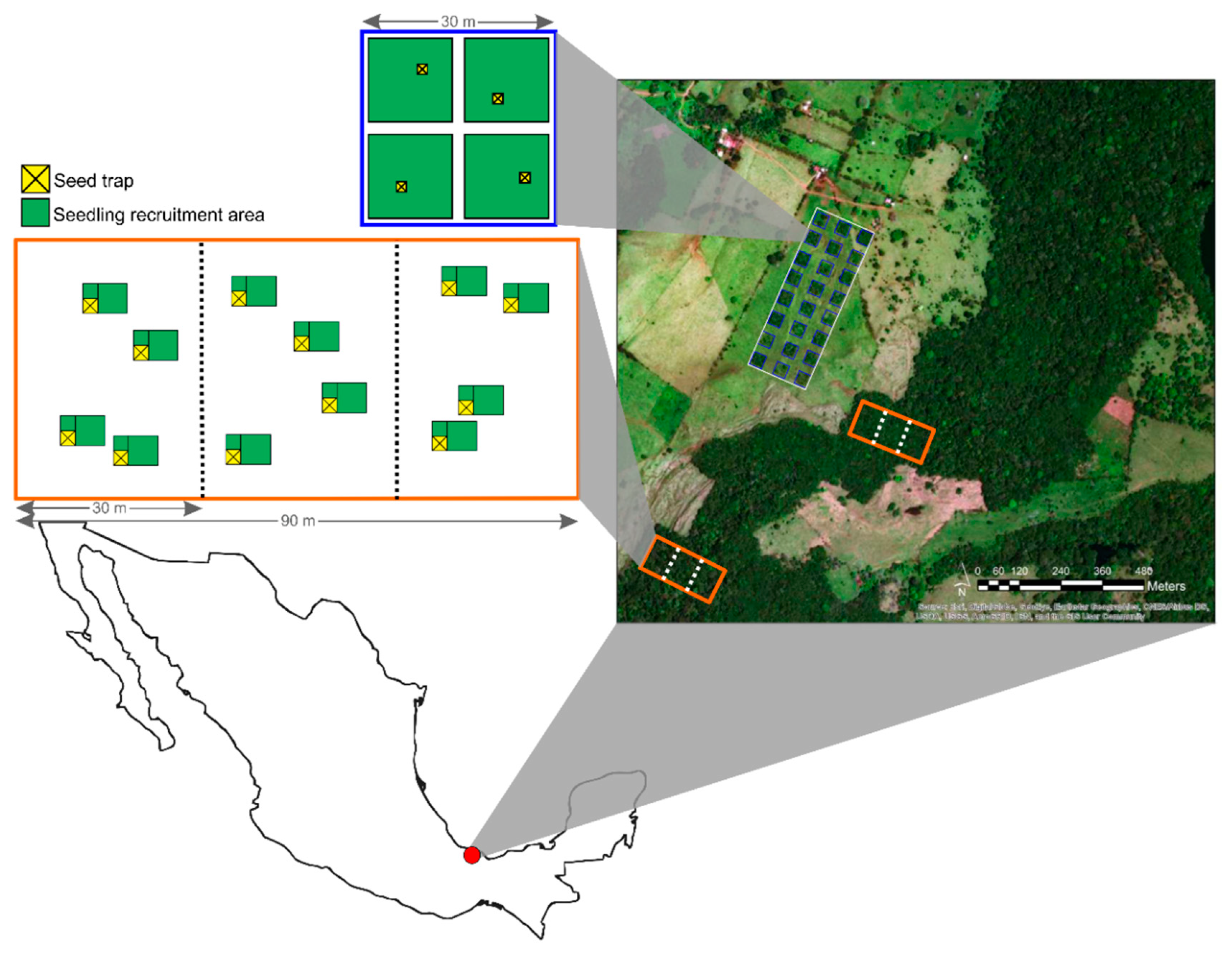
2.1. Study Site and Sampling
2.2. Calculations and Analysis
Indices
3. Results
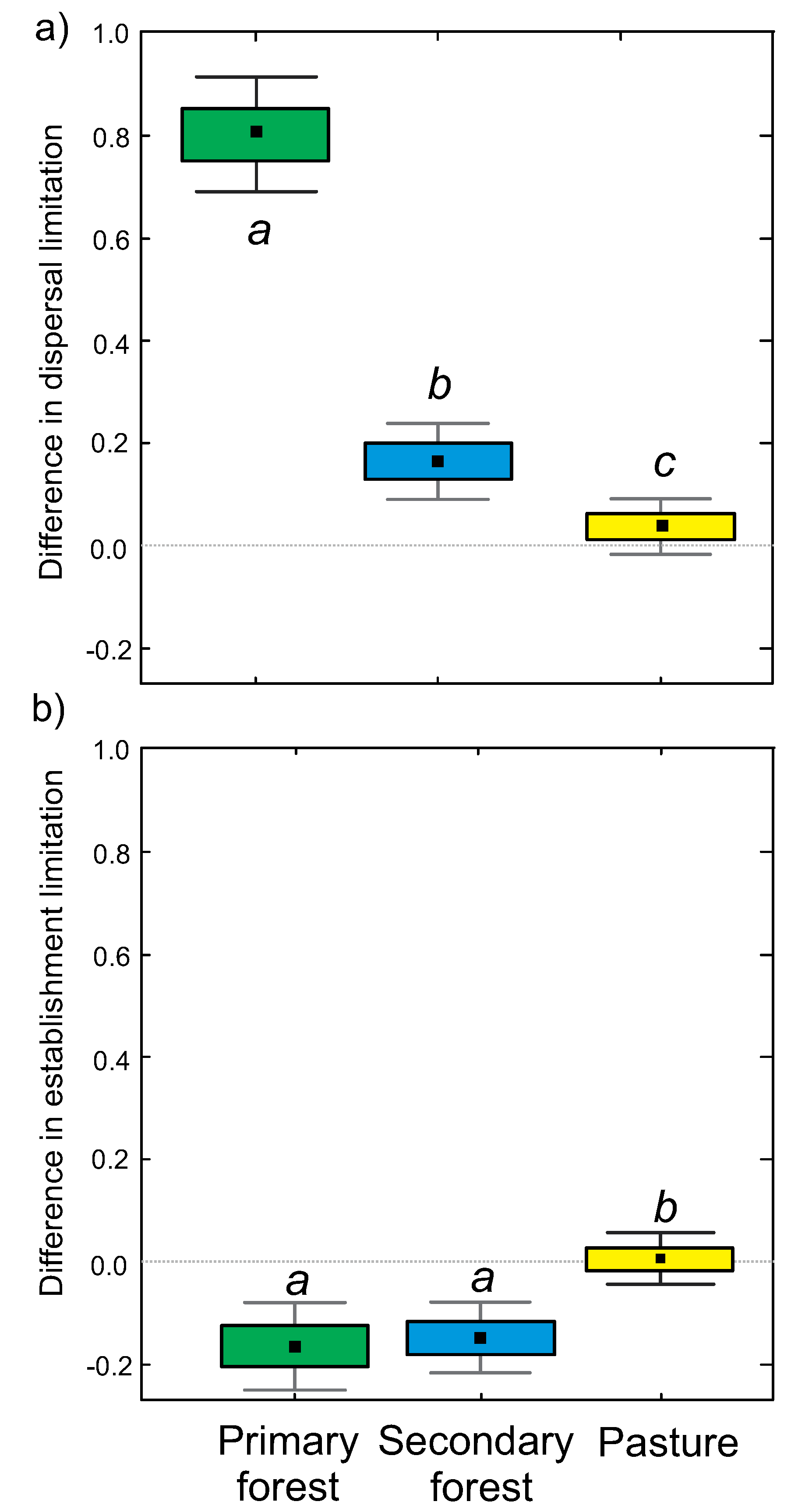
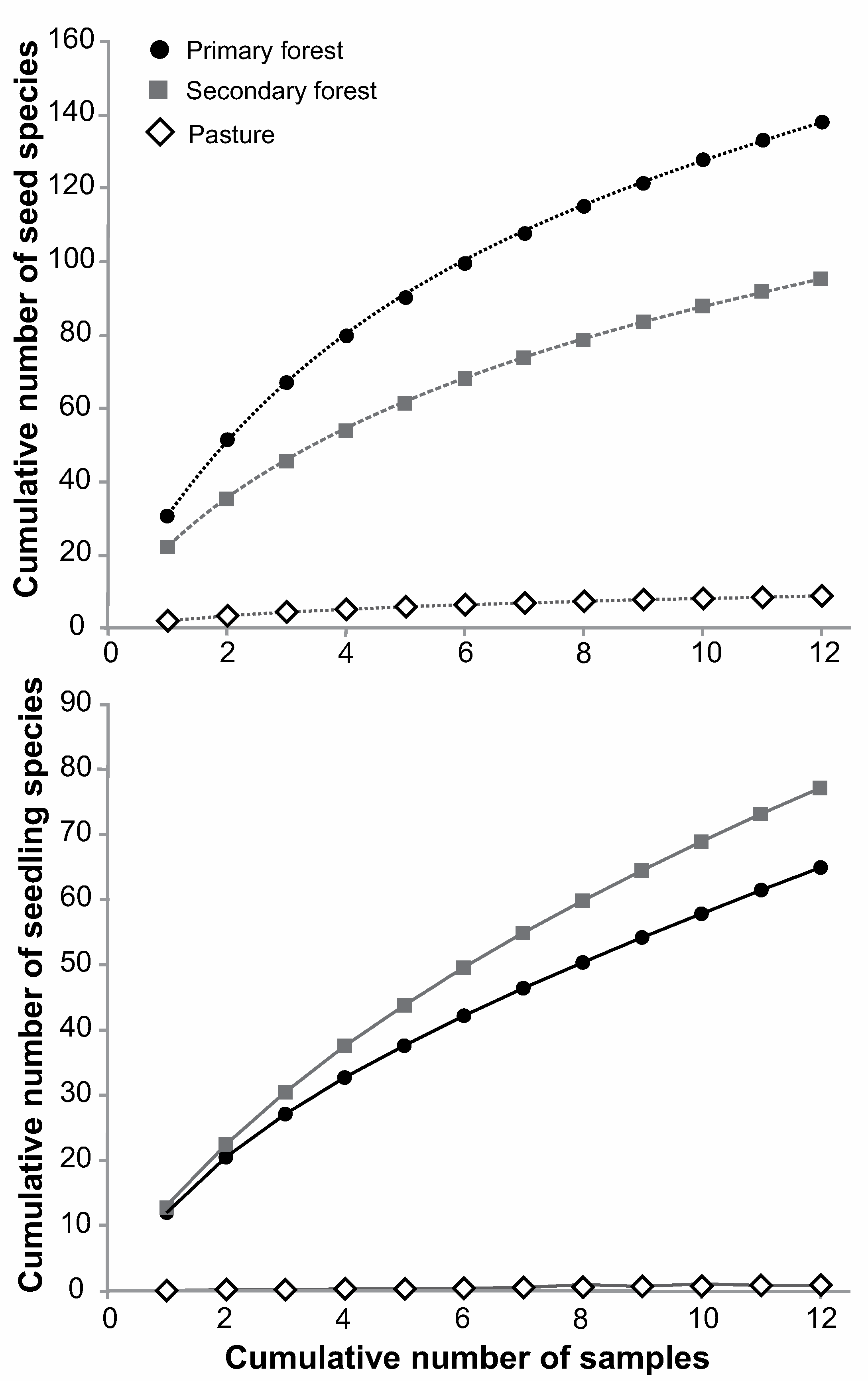
3.1. Overall Dispersal and Establishment Limitation in Different Habitats
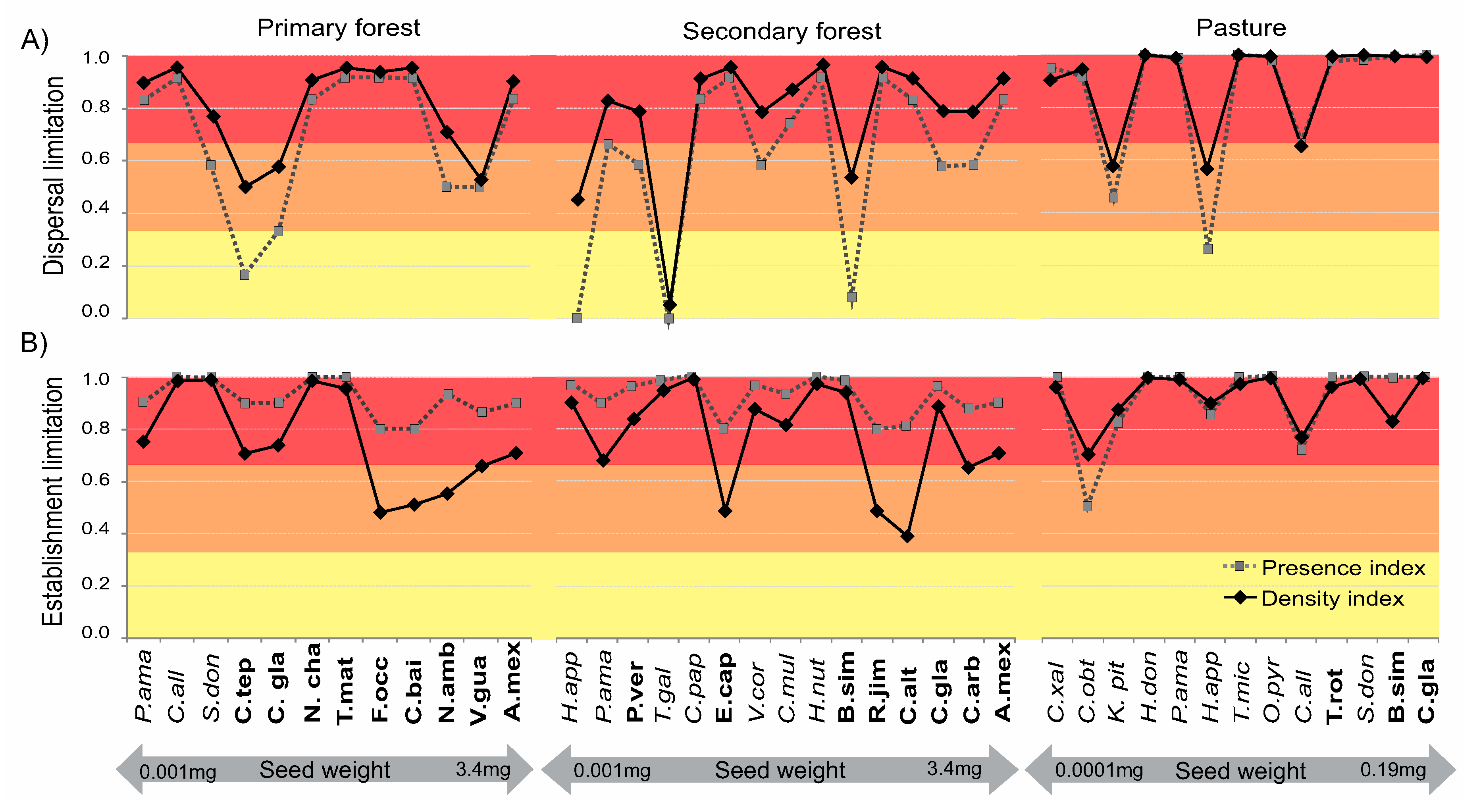
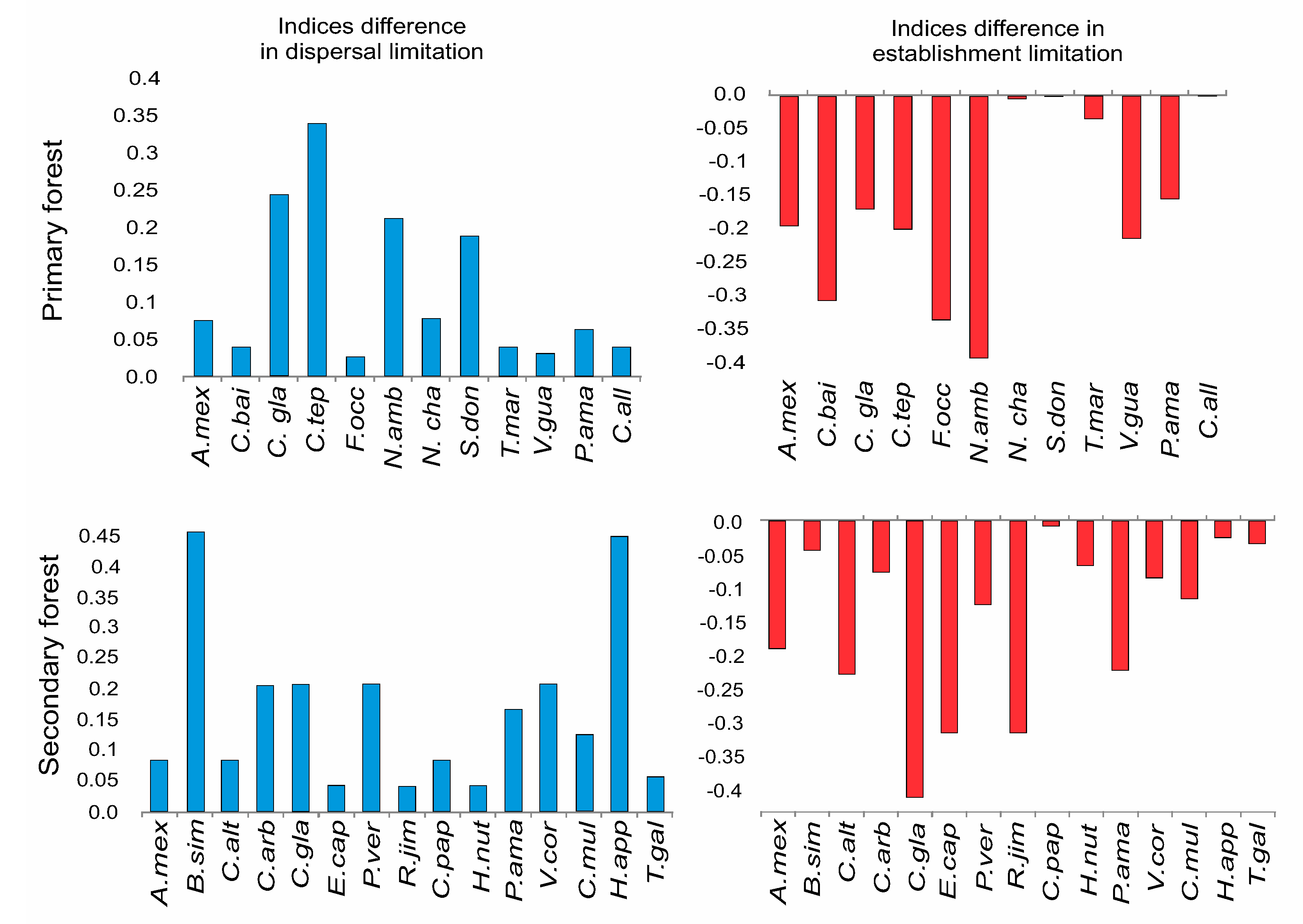
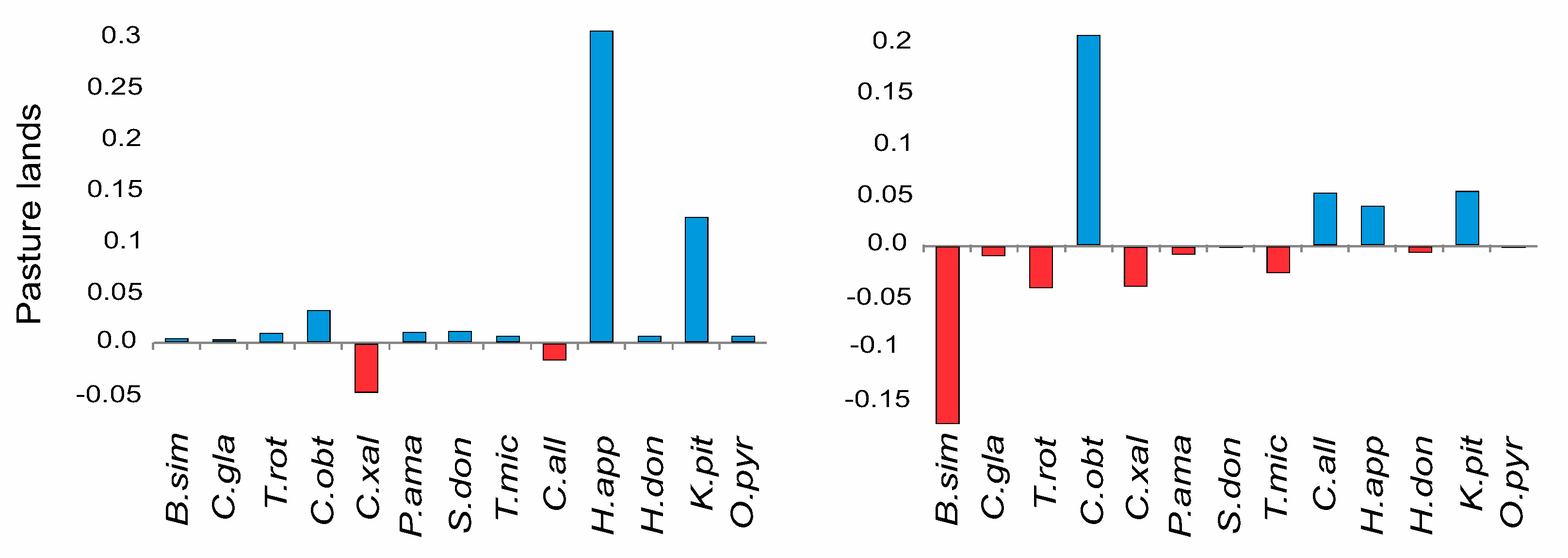
3.2. Species-Specific Dispersal and Establishment Limitation in Three Habitats
4. Discussion
4.1. Difference Between Indices
4.2. Differences between Habitats
4.3. Differences within Species
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Dispersal Limitation | Establishment Limitation | ||
|---|---|---|---|---|
| Presence Index | Density Weighted Index | Presence Index | Density Weighted Index | |
| PRIMARY FOREST | ||||
| Astrocaryum mexicanum Liebm. ex Mart. | 0.07 | 0.83 | 0.90 | 0.71 |
| Chamaedorea tepejilote Liebm. | 0.34 | 0.17 | 0.90 | 0.70 |
| Cordia alliodora (Ruiz & Pav.) Oken | 0.04 | 0.92 | 1.00 | 1.00 |
| Cupania glabra Sw. | 0.24 | 0.33 | 0.90 | 0.73 |
| Cymbopetalum baillonii R.E. Fr. | 0.04 | 0.92 | 0.80 | 0.49 |
| Faramea occidentalis (L.) A. Rich. | 0.03 | 0.92 | 0.80 | 0.47 |
| Nectandra ambigens (S.F. Blake) C.K. Allen | 0.21 | 0.50 | 0.93 | 0.54 |
| Piper amalago L. | 0.06 | 0.83 | 0.90 | 0.75 |
| Notopleura chapensis (Steyerm.) C.M. Taylor | 0.08 | 0.83 | 1.00 | 1.00 |
| Stemmadenia donnell-smithii (Rose) Woodson | 0.19 | 0.58 | 1.00 | 1.00 |
| Trichilia martiana C. DC. | 0.04 | 0.92 | 1.00 | 0.97 |
| Virola guatemalensis (Hemsl.) Warb. | 0.03 | 0.50 | 0.87 | 0.65 |
| SECONDARY FOREST | ||||
| Astrocaryum mexicanum Liebm. ex Mart. | 0.83 | 0.92 | 0.90 | 0.71 |
| Bursera simaruba (L.) Sarg. | 0.08 | 0.54 | 0.98 | 0.94 |
| Carica papaya L. | 0.83 | 0.92 | 1.00 | 0.99 |
| Chamaedorea alternans H. Wendl. | 0.83 | 0.92 | 0.80 | 0.39 |
| Cnidoscolus multilobus (Pax) I.M. Johnst. | 0.75 | 0.87 | 0.93 | 0.82 |
| Cojoba arborea (L.) Britton & Rose | 0.58 | 0.79 | 0.88 | 0.65 |
| Cupania glabra Sw. | 0.58 | 0.79 | 0.96 | 0.88 |
| Eugenia capuli (Schltdl. & Cham.) Hook. & Arn. | 0.92 | 0.96 | 0.80 | 0.48 |
| Hampea nutricia Fryxell | 0.92 | 0.96 | 1.00 | 0.98 |
| Heliocarpus appendiculatus Turcz. | 0.00 | 0.45 | 0.97 | 0.90 |
| Piper amalago L. | 0.67 | 0.83 | 0.90 | 0.68 |
| Psychotria veracruzensis Lorence & Dwyer | 0.58 | 0.79 | 0.96 | 0.84 |
| Rollinia jimenezii Saff. | 0.92 | 0.96 | 0.80 | 0.48 |
| Trichospermum galeottii (Turcz.) Kosterm. | 0.00 | 0.06 | 0.98 | 0.95 |
| Vachellia cornigera (L.) Seigler & Ebinger | 0.58 | 0.79 | 0.96 | 0.88 |
| PASTURE | ||||
| Bursera simaruba (L.) Sarg. | 0.99 | 0.99 | 1.00 | 0.83 |
| Cecropia obtusifolia Bertol. | 0.92 | 0.95 | 0.50 | 0.70 |
| Conostegia xalapensis (Bonpl.) D. Don ex DC. | 0.95 | 0.90 | 1.00 | 0.96 |
| Cordia alliodora (Ruiz & Pav.) Oken | 0.67 | 0.65 | 0.72 | 0.77 |
| Cupania glabra Sw. | 0.99 | 0.99 | 1.00 | 1.00 |
| Heliocarpus appendiculatus Turcz. | 0.26 | 0.56 | 0.86 | 0.90 |
| Heliocarpus donnellsmithii Rose | 0.99 | 0.99 | 1.00 | 0.99 |
| Koanophyllon pittieri (Klatt) R.M. King & H. Robinson | 0.46 | 0.58 | 0.83 | 0.88 |
| Ochroma pyramidale (Cav. ex Lam.) Urb. | 0.99 | 0.99 | 1.00 | 1.00 |
| Piper amalago L. | 0.98 | 0.99 | 1.00 | 0.99 |
| Stemmadenia donnell-smithii (Rose) Woodson | 0.98 | 0.99 | 1.00 | 1.00 |
| Tetrorchidium rotundatum Standl. | 0.98 | 0.99 | 1.00 | 0.96 |
| Trema micrantha (L.) Blume | 0.99 | 0.99 | 1.00 | 0.98 |
References
- Clark, J.S. Why trees migrate so fast: Confronting theory with dispersal biology and the paleorecord. Am. Nat. 1998, 152, 204–224. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; p. 375. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; Volume 1. [Google Scholar]
- Ridley, H.N. The Dispersal of Plants throughout the World; L. Reeve & Company, Limited: Ashford, UK, 1930. [Google Scholar]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–527. [Google Scholar] [CrossRef]
- Connell, J.H. On the Role of Natural Enemies in Preventing Competitive Exclusion in Some Marine Animals and in Rain Forest Trees. In Dynamics of Populations; Center for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1971. [Google Scholar]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Comita, L.S.; Muller-Landau, H.C.; Aguilar, S.; Hubbell, S.P. Asymmetric density dependence shapes species abundances in a tropical tree community. Science 2010, 329, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; HilleRisLambers, J. Seed dispersal near and far: Patterns across temperate and tropical forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Harms, K.E.; Wright, S.J.; Calderon, O.; Hernandez, A.; Herre, E.A. Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest. Nature 2000, 404, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.J.; Poulsen, J.R.; Levey, D.J. Roles of seed and establishment limitation in determining patterns of afrotropical tree recruitment. PLoS ONE 2013, 8, e63330. [Google Scholar] [CrossRef] [PubMed]
- Slocum, M.G. How tree species differ as recruitment foci in a tropical pasture. Ecology 2001, 82, 2547–2559. [Google Scholar] [CrossRef]
- Norden, N.; Chave, J.; Belbenoit, P.; Caubere, A.; Chatelet, P.; Forget, P.M.; Riera, B.; Viers, J.; Thebaud, C. Interspecific variation in seedling responses to seed limitation and habitat conditions for 14 neotropical woody species. J. Ecol. 2009, 97, 186–197. [Google Scholar] [CrossRef]
- Tilman, D. Competition and biodiversity in spatially structured habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Pacala, S.W. The consequences of recruitment limitation—Reconciling chance, history and competitive differences between plants. J. Theor. Biol. 1995, 176, 1–12. [Google Scholar] [CrossRef]
- Platt, W.J.; Weis, I.M. Resource partitioning and competition within a guild of fugitive prairie plants. Am. Nat. 1977, 111, 479–513. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342. [Google Scholar] [CrossRef] [PubMed]
- Terborgh, J.; Alvarez-Loayza, P.; Dexter, K.; Cornejo, F.; Carrasco, C. Decomposing dispersal limitation: Limits on fecundity or seed distribution? J. Ecol. 2011, 99, 935–944. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; González-Montagut, R. Seed rain from forest fragments into tropical pastures in Los Tuxtlas, Mexico. Plant Ecol. 1999, 145, 255–265. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Howe, H.F. Restoring tropical diversity: Beating the time tax on species loss. J. Appl. Ecol. 2003, 40, 423–429. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Flores-Palacios, A.; De La Peña-Domene, M.; Howe, H.F. Seed rain in a tropical agricultural landscape. J. Trop. Ecol. 2009, 25, 541–550. [Google Scholar] [CrossRef]
- Howe, H.F.; Urincho-Pantaleon, Y.; de la Peña-Domene, M.; Martinez-Garza, C. Early seed fall and seedling emergence: Precursors to tropical restoration. Oecologia 2010, 164, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Ingle, N.R. Seed dispersal by wind, birds, and bats between Philippine montane rainforest and successional vegetation. Oecologia 2003, 134, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L.; Letcher, S.G.; van Breugel, M.; Martinez-Ramos, M.; Bongers, F.; Finegan, B. Rates of change in tree communities of secondary neotropical forests following major disturbances. Philos. Trans. R. Soc. B-Biol. Sci. 2007, 362, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Boucher, D.H.; Vandermeer, J.H.; de la Cerda, I.G.; Mallona, M.A.; Perfecto, I.; Zamora, N. Post-agriculture versus post-hurricane succession in southeastern Nicaraguan rain forest. Plant Ecol. 2001, 156, 131–137. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Guariguata, M.R. Natural regeneration as a tool for large-scale forest restoration in the tropics: Prospects and challenges. Biotropica 2016, 48, 716–730. [Google Scholar] [CrossRef]
- Martínez-Garza, C.; Osorio-Beristain, M.; Alcalá-Martínez, R.; Valenzuela-Galván, D.; Mariano, N.; Ceccon, E. Ocho años de restauración experimental en las selvas estacionales de México. Experiencias Mexicanas en la Restauración de los Ecosistemas 2016, 1, 385–405. [Google Scholar]
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Hubbell, S.P.; Foster, R.B. Assessing recruitment limitation: Concepts, methods and case-studies from a tropical forest. In Seed Dispersal and Frugivory; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CAB International: Wallingford, UK, 2002; pp. 35–53. [Google Scholar]
- De La Peña-Domene, M.; Martínez-Garza, C.; Howe, H.F. Early recruitment dynamics in tropical restoration. Ecol. Appl. 2013, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- De la Peña-Domene, M.; Howe, H.F.; Cruz-León, E.; Jiménez-Rolland, R.; Lozano-Huerta, C.; Martínez-Garza, C. Seed to seedling transitions in successional habitats across a tropical landscape. Oikos 2017, 126, 410–419. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples. Version 8.0 User’s Guide and Application. 2006. Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 16 July 2012).
- Williams, G.C. Sex and Evolution; Princeton University Press: Princeton, NJ, USA, 1975. [Google Scholar]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; de Lao, S.L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Vázquez-Yanes, C.; Orozco-Segovia, A. Effects of litter from a tropical rainforest on tree seed germination and establishment under controlled conditions. Tree Physiol. 1992, 11, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Muller-Landau, H.C. The tolerance–fecundity trade-off and the maintenance of diversity in seed size. Proc. Natl. Acad. Sci. USA 2010, 107, 4242–4247. [Google Scholar] [CrossRef] [PubMed]
- De La Peña-Domene, M.; Martínez-Garza, C.; Ayestarán-Hernández, L.M.; Howe, H.F. Plant attributes that drive dispersal and establishment limitation in tropical agricultural landscapes. Forests 2018. submitted. [Google Scholar]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Peña-Domene, M.; Martínez-Garza, C. Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests. Forests 2018, 9, 570. https://doi.org/10.3390/f9090570
De la Peña-Domene M, Martínez-Garza C. Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests. Forests. 2018; 9(9):570. https://doi.org/10.3390/f9090570
Chicago/Turabian StyleDe la Peña-Domene, Marinés, and Cristina Martínez-Garza. 2018. "Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests" Forests 9, no. 9: 570. https://doi.org/10.3390/f9090570
APA StyleDe la Peña-Domene, M., & Martínez-Garza, C. (2018). Integrating Density into Dispersal and Establishment Limitation Equations in Tropical Forests. Forests, 9(9), 570. https://doi.org/10.3390/f9090570