Red deer (Cervus elaphus) Did Not Play the Role of Maintenance Host for Bluetongue Virus in France: The Burden of Proof by Long-Term Wildlife Monitoring and Culicoides Snapshots

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Wildlife Investigations
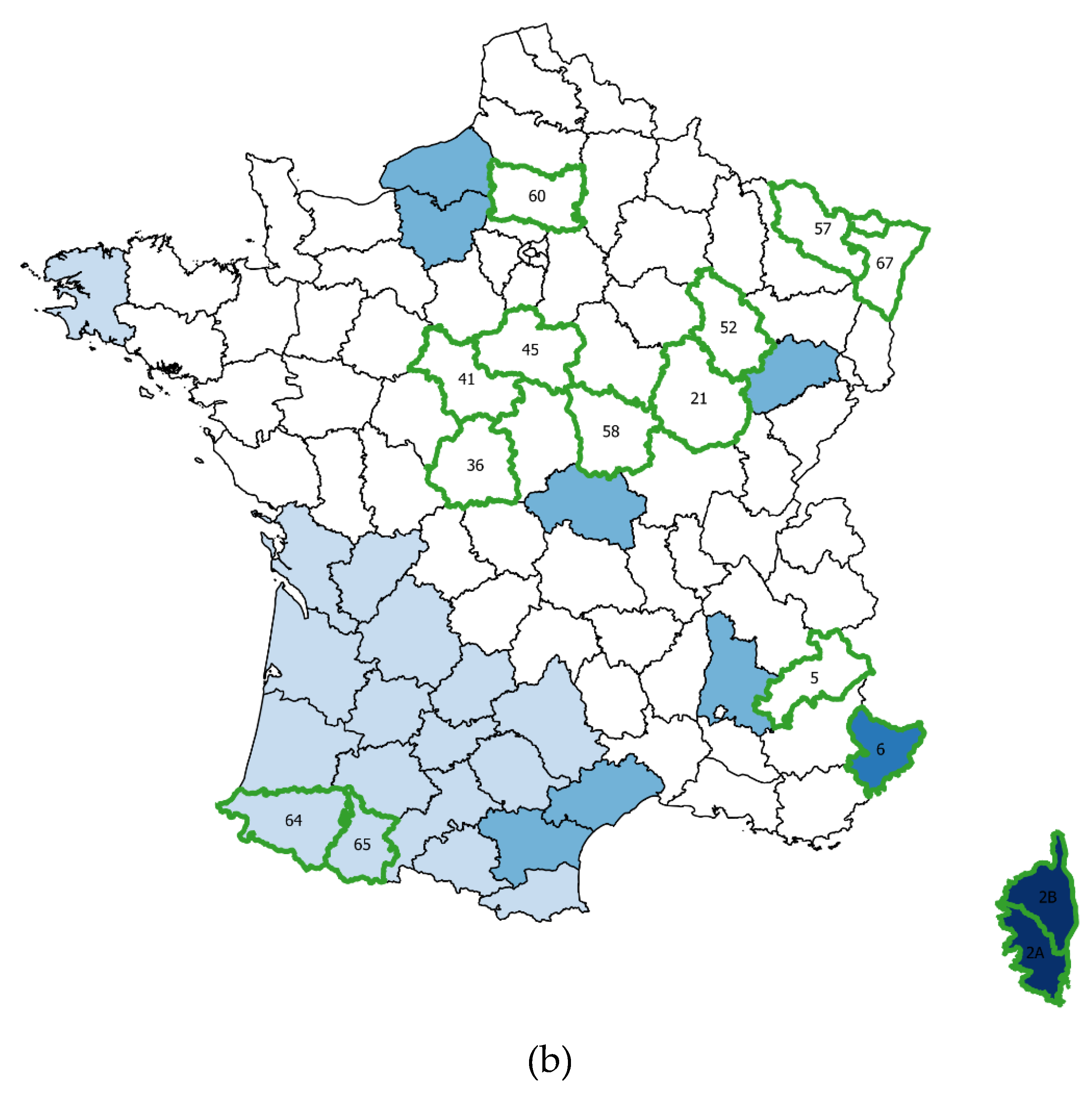
2.1.1. Wildlife Sampling
2.1.2. Diagnostic Tests for BTV
2.1.3. Risk Factors
2.1.4. Statistical Analyses of the Red Deer Serological Data
- The individual serological status based on the c-ELISA result; and
- Among the seropositive samples, the mean VNT titer regarding the BTV8 strain.
2.2. Entomological Analyses
2.2.1. Entomological Collections and Identification
2.2.2. Description of Culicoides Species Communities in the Different Environments
3. Results
3.1. Red Deer Study
3.1.1. Samples and Average Seroprevalence
3.1.2. Evolution of Seroprevalence (c-ELISA) and BTV8 NA Titers in the Continental France
3.2. Culicoides Species Communities
3.2.1. Culicoides Collections
3.2.2. Description of Culicoides Species Communities in the Different Environments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Age | Dep | Season | Sex | Age*season | Dep*season | df | logLik | QAICc | Delta |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | + | + | −0.373 | NA | + | NA | 18 | −799.397 | 1540.870 | 0 |
| 2 | + | + | −0.373 | + | + | NA | 19 | −799.397 | 1542.907 | 2.037 |
| Model | Age | Dep | Season | Sex | Age*dep | Age*season | Dep*season | df | logLik | QAICc | Delta |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | NA | NA | −0.243 | NA | NA | NA | NA | 3 | −390.775 | 267.471 | 0 |
| 2 | NA | NA | −0.244 | + | NA | NA | NA | 4 | −388.289 | 267.895 | 0.424 |
| 3 | + | NA | −0.262 | NA | NA | NA | NA | 5 | −386.921 | 269.077 | 1.605 |
| 4 | + | NA | −0.197 | NA | NA | + | NA | 7 | −382.236 | 270.193 | 2.722 |
References
- Mertens, P.; Baylis, M.; Mellor, P. Bluetongue; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Zientara, S.; Sánchez-Vizcaíno, J.M. Control of bluetongue in Europe. Vet. Microbiol. 2013, 165, 33–37. [Google Scholar] [CrossRef]
- Stallknecht, D.E.; Howerth, E.W. Epidemiology of bluetongue and epizootic haemorrhagic disease in wildlife: Surveillance methods. Vet. Ital. 2004, 40, 203–207. [Google Scholar]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef]
- De La Puente, J.M.; Merino, S.; Tomas, G.; Moreno, J.; Morales, J.; Lobato, E.; Talavera, S.; Monteys, V.S.I. Factors affecting Culicoides species composition and abundance in avian nests. Parasitology 2009, 136, 1033–1041. [Google Scholar] [CrossRef]
- Talavera, S.; Muñoz-Muñoz, F.; Durán, M.; Verdún, M.; Soler-Membrives, A.; Oleaga, Á.; Arenas, A.; Ruiz-Fons, F.; Estrada, R.; Pagès, N. Culicoides Species Communities Associated with Wild Ruminant Ecosystems in Spain: Tracking the Way to Determine Potential Bridge Vectors for Arboviruses. PLoS ONE 2015, 10, e0141667. [Google Scholar]
- Rádrová, J.; Mračková, M.; Galková, Z.; Lamka, J.; Račka, K.; Barták, P.; Votýpka, J. Seasonal Dynamics, Parity Rate, and Composition of Culicoides (Diptera: Ceratopogonidae) Occurring in the Vicinity of Wild and Domestic Ruminants in the Czech Republic. J. Med. Entomol. 2016, 53, 416–424. [Google Scholar] [CrossRef]
- Talavera, S.; Muñoz-Muñoz, F.; Verdún, M.; Pujol, N.; Pagès, N. Revealing potential bridge vectors for BTV and SBV: A study on Culicoides blood feeding preferences in natural ecosystems in Spain. Med. Vet. Èntomol. 2017, 32, 35–40. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; Sánchez-Matamoros, A.; Gortázar, C.; Sánchez-Vizcaíno, J.M. The role of wildlife in bluetongue virus maintenance in Europe: Lessons learned after the natural infection in Spain. Virus Res. 2014, 182, 50–58. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; Reyes-García, Á.R.; Alcaide, V.; Gortázar, C. Spatial and Temporal Evolution of Bluetongue Virus in Wild Ruminants, Spain. Emerg. Infect. Dis. 2008, 14, 951–953. [Google Scholar] [CrossRef]
- Linden, A.; Gregoire, F.; Nahayo, A.; Hanrez, D.; Mousset, B.; Massart, L.; De Leeuw, I.; Vandemeulebroucke, E.; Vandenbussche, F.; De Clercq, K. Bluetongue virus in wild deer, Belgium, 2005–2008. Emerg. Infect. Dis. 2010, 16, 833–836. [Google Scholar] [CrossRef]
- Falconi, C.; López-Olvera, J.R.; Gortazar, C. BTV infection in wild ruminants, with emphasis on red deer: A review. Vet. Microbiol. 2011, 151, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Pioz, M.; Beard, E.; Durand, B.; Gibert, P.; Gauthier, D.; Klein, F.; Maillard, D.; Saint-Andrieux, C.; Saubusse, T.; et al. Bluetongue dynamics in French wildlife: Exploring the driving forces. Transbound. Emerg. Dis. 2014, 61, e12–e24. [Google Scholar] [CrossRef]
- López-Olvera, J.R.; Falconi, C.; Fernández-Pacheco, P.; Fernández-Pinero, J.; Sánchez, M.A.; Palma, A.; Herruzo, I.; Vicente, J.; Jiménez-Clavero, M.A.; Arias, M.; et al. Experimental infection of European red deer (Cervus elaphus) with bluetongue virus serotypes 1 and 8. Vet. Microbiol. 2010, 145, 148–152. [Google Scholar] [CrossRef]
- Carpenter, S.; Wilson, A.; Mellor, P.S. Culicoides and the emergence of bluetongue virus in northern Europe. Trends Microbiol. 2009, 17, 172–178. [Google Scholar] [CrossRef]
- Pioz, M.; Guis, H.; Calavas, D.; Durand, B.; Abrial, D.; Ducrot, C. Estimating front-wave velocity of infectious diseases: A simple, efficient method applied to bluetongue. Vet. Res. 2011, 42, 60. [Google Scholar] [CrossRef]
- Sailleau, C.; Breard, E.; Viarouge, C.; Vitour, D.; Romey, A.; Garnier, A.; Fablet, A.; Lowenski, S.; Gorna, K.; Caignard, G.; et al. Re-Emergence of Bluetongue Virus Serotype 8 in France, 2015. Transbound. Emerg. Dis. 2017, 64, 998–1000. [Google Scholar] [CrossRef]
- Bréard, E.; Sailleau, C.; Quenault, H.; Lucas, P.; Viarouge, C.; Touzain, F.; Fablet, A.; Vitour, D.; Attoui, H.; Zientara, S.; et al. Complete Genome Sequence of Bluetongue Virus Serotype 8, Which Reemerged in France in August 2015. Genome Announc. 2016, 4, e00163-16. [Google Scholar] [CrossRef] [Green Version]
- Courtejoie, N.; Durand, B.; Bournez, L.; Gorlier, A.; Breard, E.; Sailleau, C.; Vitour, D.; Zientara, S.; Baurier, F.; Gourmelen, C.; et al. Circulation of bluetongue virus 8 in French cattle, before and after the re-emergence in 2015. Transbound. Emerg. Dis. 2018, 65, 281–284. [Google Scholar] [CrossRef]
- Zientara, S.; Sailleau, C.; Viarouge, C.; Höper, D.; Beer, M.; Jenckel, M.; Hoffmann, B.; Romey, A.; Bakkali-Kassimi, L.; Fablet, A.; et al. Novel bluetongue virus in goats, Corsica, France, 2014. Emerg. Infect. Dis. 2014, 20, 2123–2125. [Google Scholar] [CrossRef]
- Sailleau, C.; Viarouge, C.; Breard, E.; Perrin, J.B.; Doceul, V.; Vitour, D.; Zientara, S. Emergence of Bluetongue Virus Serotype 1 in French Corsica Island in September 2013. Transbound. Emerg. Dis. 2015, 62, 89–91. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Arenas-Montes, A.; Lorca-Oró, C.; Pujols, J.; González, M.Á.; Napp, S.; Gómez-Guillamón, F.; Zorrilla, I.; Miguel, E.S.; Arenas, A. Role of wild ruminants in the epidemiology of bluetongue virus serotypes 1, 4 and 8 in Spain. Vet. Res. 2011, 42, 88. [Google Scholar] [CrossRef]
- Lorca-Oró, C.; López-Olvera, J.R.; Ruiz-Fons, F.; Acevedo, P.; García-Bocanegra, I.; Oleaga, Á.; Gortázar, C.; Pujols, J. Long-Term Dynamics of Bluetongue Virus in Wild Ruminants: Relationship with Outbreaks in Livestock in Spain, 2006–2011. PLoS ONE 2014, 9, e100027. [Google Scholar] [CrossRef]
- Arenas-Montes, A.; Paniagua, J.; Arenas, A.; Lorca-Oró, C.; Carbonero, A.; Cano-Terriza, D.; García-Bocanegra, I. Spatial–temporal trends and factors associated with the bluetongue virus seropositivity in large game hunting areas from southern Spain. Transbound. Emerg. Dis. 2016, 63, e339–e346. [Google Scholar] [CrossRef]
- Rossi, S. Bluetongue in red deer: An emerging wild reservoir? Oral communication. In Proceedings of the EWDA Conference 2014, Edinburg, Scotland, 25–29 August 2014. [Google Scholar]
- Kidjo, N.; Feracci, G.; Bideau, E.; Gonzalez, G.; Mattéi, C.; Marchand, B.; Aulagnier, S. Extirpation and reintroduction of the Corsican red deer Cervus elaphus corsicanus in Corsica. Oryx 2007, 41, 488–494. [Google Scholar] [CrossRef]
- Kreeger, T.J.; Arnemo, J.M. Handbook of Wildlife Chemical Immobilization; International Wildlife Veterinary Services: Laramie, WY, USA, 1996. [Google Scholar]
- Boadella, M.; Gortázar, C. Effect of haemolysis and repeated freeze-thawing cycles on wild boar serum antibody testing by ELISA. BMC Res. Notes 2011, 4, 498. [Google Scholar] [CrossRef]
- Breard, E.; Belbis, G.; Hamers, C.; Moulin, V.; Lilin, T.; Moreau, F.; Millemann, Y.; Montange, C.; Sailleau, C.; Durand, B. Evaluation of humoral response and protective efficacy of two inactivated vaccines against bluetongue virus after vaccination of goats. Vaccine 2011, 29, 2495–2502. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. A practical information-theoretic approach. In Model Selection and Multimodel Inference, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org/ (accessed on 4 September 2019).
- Barton, K. MuMIn: Multi-Model Inference; R Package Version 1.43.6; 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 4 September 2019).
- Venail, R.; Balenghien, T.; Guis, H.; Tran, A.; Setier-Rio, M.-L.; Delecolle, J.-C.; Mathieu, B.; Cetre-Sossah, C.; Martinez, D.; Languille, J.; et al. Assessing Diversity and Abundance of Vector Populations at a National Scale: Example of Culicoides Surveillance in France after Bluetongue Virus Emergence. In Arthropods as Vectors of Emerging Diseases; Mehlhorn, H., Ed.; Parasitology Research Monographs; Springer: Berlin, Germany, 2002; Volume 3, pp. 77–102. [Google Scholar]
- Delécolle, J.C. Nouvelle Contribution a L’etude Systématique et Iconographique des Espèces du Genre Culicoides (Diptera: Ceratopogonidae) du Nord-Est de la France. Ph.D. Thesis, Université Louis Pasteur de Strasbourg, Strasbourg, Alsace, France, 9 May 1985. [Google Scholar]
- Mathieu, B.; Cêtre-Sossah, C.; Garros, C.; Chavernac, D.; Balenghien, T.; Carpenter, S.; Setier-Rio, M.-L.; Vignes-Lebbe, R.; Ung, V.; Candolfi, E.; et al. Development and validation of IIKC: An interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasites Vectors 2012, 5, 137. [Google Scholar] [CrossRef]
- Harrup, L.E.; Bellis, G.A.; Balenghien, T.; Garros, C. Culicoides Latreille (Diptera: Ceratopogonidae) taxonomy: Current challenges and future directions. Infect. Genet. Evol. 2015, 30, 249–266. [Google Scholar] [CrossRef]
- Garros, C.; Gardes, L.; Allène, X.; Rakotoarivony, I.; Viennet, E.; Rossi, S.; Balenghien, T. Adaptation of a species-specific multiplex PCR assay for the identification of blood meal source in Culicoides (Ceratopogonidae: Diptera): Applications on Palaearctic biting midge species, vectors of Orbiviruses. Infect. Genet. Evol. 2011, 11, 1103–1110. [Google Scholar] [CrossRef]
- Nolan, D.V.; Carpenter, S.; Barber, J.; Mellor, P.S.; Dallas, J.F.; Mordue, A.J.; Piertney, S.B. Rapid diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet. Microbiol. 2007, 124, 82–94. [Google Scholar] [CrossRef]
- Lebart, L.M.; Piron, A. Morineau. In Statistique Exploratoire Multidimensionnelle, 3rd ed.; Dunod: Paris, France, 2000. [Google Scholar]
- Dolédec, S.; Chessel, D. Rythmes saisonniers et composantes stationnelles en milieu aquatique I- Description d’un plan d’observations complet par projection de variables. Acta Oecol. 1987, 3, 403–426. [Google Scholar]
- Oksanen, J.; Guillaume, F.; Blanchet, M.; Friendly, R.; Kindt, P.; Legendre, D.; McGlinn, P.R.; Minchin, R.B.; O’Hara, G.L.; Simpson, P.; et al. vegan: Community Ecology Package, 2019, R Package Version 2.5-5. Available online: https://CRAN.R-project.org/package=vegan (accessed on 4 September 2019).
- Dray, S.; Dufour, A.-B. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Lesnoff, M.; Messad, S.; Lancelot, R. tdisplay: Tool Box for Data Importation and Display. R package Version 0.7-0/r11. Available online: https://R-Forge.R-project.org/projects/tdisplay/ (accessed on 4 September 2019).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Balenghien, T. Cirad, Montpellier, France. Description of the Culicoides species communities, from the 2008 collections in different ecological and climatic French regions, using ecological measures of species assemblages, 2019.
- Jarnemo, A. Male red deer (Cervus elaphus) dispersal during the breeding season. J. Ethol. 2011, 29, 329–336. [Google Scholar] [CrossRef]
- Garros, C.; Balenghien, T. Les culicoïdes (Diptera: Ceratopogonidae). In Entomologie Médicale et Vétérinaire; Duvallet, G., Fontenille, D., Robert, V., Eds.; IRD: Marseille, France, 2017; pp. 345–365. [Google Scholar]
- Calvo, J.; Berzal, B.; Calvete, C.; Miranda, M.; Estrada, R.; Lucientes, J. Host feeding patterns of Culicoides species (Diptera: Ceratopogonidae) within the Picos de Europa National Park in northern Spain. Bull. Èntomol. Res. 2012, 102, 692–697. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Havelka, P.; Schaefer, H.M.; Segelbacher, G. Bloodmeal Analysis Reveals Avian Plasmodium Infections and Broad Host Preferences of Culicoides (Diptera: Ceratopogonidae) Vectors. PLoS ONE 2012, 7, e31098. [Google Scholar] [CrossRef]
- Vasic, A.; Zdravkovic, N.; Anita, D.; Bojkovski, J.; Marinov, M.; Mathis, A.; Niculaua, M.; Oslobanu, E.L.; Pavlovic, I.; Petric, D.; et al. Species diversity, host preference and arbovirus detection of Culicoides (Diptera: Ceratopogonidae) in south-eastern Serbia. Parasite Vector 2019, 12, 61. [Google Scholar] [CrossRef]
- Carpenter, S.; Veronesi, E.; Mullens, B.; Venter, G. Vector competence of Culicoides for arboviruses: Three major periods of research, their influence on current studies and future directions. Rev. Sci. Tech. OIE 2015, 34, 97–112. [Google Scholar] [CrossRef]
- Di Gialleonardo, L.; Migliaccio, P.; Teodori, L.; Savini, G. The length of BTV-8 viraemia in cattle according to infection doses and diagnostic techniques. Res. Vet. Sci. 2011, 91, 316–320. [Google Scholar] [CrossRef]
- Saubusse, T.; Masson, J.-D.; Le Dimma, M.; Abrial, D.; Marcé, C.; Martin-Schaller, R.; Dupire, A.; Le Potier, M.-F.; Rossi, S. How to survey classical swine fever in wild boar (Sus scrofa) after the completion of oral vaccination? Chasing away the ghost of infection at different spatial scales. Vet. Res. 2016, 47, 21. [Google Scholar] [CrossRef]
- Hard, J.J.; Mills, L.S.; Peek, J.M. Genetic implications of reduced survival of male red deer Cervus elaphus under harvest. Wildl. Biol. 2006, 12, 427–441. [Google Scholar] [CrossRef]
- Bournez, L.; Cavalerie, L.; Sailleau, C.; Breard, E.; Zanella, G.; De Almeida, R.S.; Pedarrieu, A.; Garin, E.; Tourette, I.; Dion, F.; et al. Estimation of French cattle herd immunity against bluetongue serotype 8 at the time of its re-emergence in 2015. BMC Vet. Res. 2018, 14, 65. [Google Scholar] [CrossRef]
- Durand, B.; Zanella, G.; Biteau-Coroller, F.; Locatelli, C.; Baurier, F.; Simon, C.; Le Dréan, E.; Delaval, J.; Prengère, E.; Beauté, V.; et al. Anatomy of Bluetongue virus Serotype 8 Epizootic Wave, France, 2007–2008. Emerg. Infect. Dis. 2010, 16, 1861–1868. [Google Scholar] [CrossRef]






| Department | Zone | 2008–2009 | 2009–2010 | 2010–2013 | 2013–2014 | 2014–2015 | 2015–2016 |
|---|---|---|---|---|---|---|---|
| 05 Hautes-Alpes | B | 6 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 06 Alpes Maritimes | B | 3 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 21 Côte d’Or | B | 593 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 2A Corse du Sud | / | 0 | 0 | 0 | 80 (BTV1) | 0 | 0 |
| 2B Haute-Corse | / | 0 | 0 | 0 | 89 (BTV1) | 0 | 0 |
| 36* Indre | A | 560 (BTV8) | 0 | 0 | 0 | 0 | 1 (BTV8) |
| 41 Loir-et-Cher | B | 177 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 45* Loiret | A | 102 (BTV8) | 0 | 0 | 0 | 0 | 1 (BTV8) |
| 52 Haute-Marne | B | 313 (BTV8) | 1 (BTV8) | 0 | 0 | 0 | 0 |
| 57 Moselle | B | 116 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 58* Nièvre | A | 1169 (BTV8) | 2 (BTV8) | 0 | 0 | 0 | 7 (BTV8) |
| 60 Oise | B | 28 (BTV8) | 0 | 0 | 0 | 0 | 0 |
| 64 Pyrénées Altantiques | B | 412 (BTV1, 8) | 0 | 0 | 0 | 0 | 0 |
| 65 Hautes Pyrénées | B | 115 (BTV1, 8) | 0 | 0 | 0 | 0 | 0 |
| Year | 2008–2009 | 2009–2010 | 2010–2011 | 2011–2012 | 2012–2013 | 2013–2014 | 2014–2015 | 2015–2016 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zone | Dep | pos | tot | pos | tot | pos | tot | pos | tot | pos | tot | pos | tot | pos | tot | pos | tot |
| B | 05 | 1 | 11 | 0 | 45 | ||||||||||||
| B | 06 | 0 | 1 | 1 | 13 | ||||||||||||
| B | 21 | 32* | 46 | 23* | 49 | 10* | 34 | 5 | 13 | ||||||||
| / | 2A | 4 | 15 | 5* | 12 | ||||||||||||
| / | 2B | 2 | 23 | 0 | 33 | 20* | 33 | 11 | 65 | ||||||||
| A | 36 | 59* | 122 | 34* | 117 | 3 | 55 | ||||||||||
| B | 41 | 40* | 62 | 27* | 108 | 12* | 54 | 20* | 130 | 17 | 101 | 16 | 93 | 15 | 115 | 11 | 50 |
| A | 45 | 7 | 14 | 1 | 11 | 1 | 9 | 0 | 6 | ||||||||
| B | 52 | 32* | 47 | 16* | 39 | 4* | 40 | 9* | 63 | ||||||||
| B | 57 | 5* | 33 | 6 | 32 | 1 | 21 | 1 | 25 | ||||||||
| A | 58 | 3 | 28 | 1 | 40 | 2 | 27 | 0 | 12 | ||||||||
| B | 60 | 7 | 68 | 3 | 71 | ||||||||||||
| B | 64 | 14* | 23 | 4 | 20 | 4 | 30 | 3 | 21 | ||||||||
| B | 65 | 7 | 19 | 19* | 52 | 4 | 17 | 12 | 49 | 5 | 33 | ||||||
| B | 67 | 5 | 39 | 7 | 62 | 1 | 41 | 2 | 56 | 1 | 62 | ||||||
| Zone | Dep | Result | Type | 2008–2009 | 2009–2010 | 2010–2011 | 2011–2012 | 2012–2013 | 2013–2014 | 2014–2015 | 2015–2016 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| / | 2A | neg | BTV1 | - | - | - | - | 4 | 9 | - | - |
| pos | - | - | - | - | 0 | 3 | - | - | |||
| / | 2B | neg | BTV1 | - | - | - | 20 | - | 22 | - | 5 |
| pos | - | - | - | 0 | - | 11 | - | 0 | |||
| B | 64 | neg | BTV1 | 0 | 4 | - | 4 | 3 | - | - | - |
| pos | 14 | 0 | - | 0 | 0 | - | - | - | |||
| B | 65 | neg | BTV1 | 0 | 18 | 4 | 6 | 5 | - | - | - |
| pos | 8 | 1 | 0 | 0 | 0 | - | - | - | |||
| B | 21 | neg | BTV8 | 0 | 26 | - | 9 | 5 | - | - | - |
| pos | 32 | 7 | - | 0 | 0 | - | - | - | |||
| B | 41 | neg | BTV8 | 4 | 2 | 12 | 20 | 17 | 16 | - | 5 |
| pos | 36 | 25 | 0 | 0 | 0 | 0 | - | 0 | |||
| B | 52 | neg | BTV8 | 9 | 15 | - | 4 | 9 | - | - | - |
| pos | 23 | 1 | - | 0 | 0 | - | - | - | |||
| B | 57 | neg | BTV8 | 0 | 3 | 1 | 1 | - | - | - | - |
| pos | 5 | 3 | 0 | 0 | - | - | - | - | |||
| B | 60 | neg | BTV8 | - | - | 7 | 4 | - | - | - | |
| B | 67 | neg | BTV8 | 0 | 5 | 1 | 2 | 1 | - | - | - |
| pos | 5 | 2 | 0 | 0 | 0 | - | - | - | |||
| Total of RD PCR tested % of BTV8 PCR positive | 136 | 112 | 18 | 73 | 48 | 61 | 0 | 10 | |||
| 90.4 | 34.8 | 0 | 0 | 0 | 23 | 0 | 0 | ||||
| VNT Titers | 0 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | |
|---|---|---|---|---|---|---|---|---|---|
| zone | dep | ||||||||
| B | 05 | 1 | 3 | ||||||
| B | 06 | 1 | |||||||
| B | 21 | 2 | 3 | 3 | 4 | 10 | 8 | 3 | 1 |
| / | 2B* | 2* | |||||||
| A | 36 | 7 | 7 | 32 | 51 | 15 | 8 | 5 | |
| B | 41 | 5 | 9 | 19 | 25 | 17 | 16 | 3 | |
| A | 45 | 2 | 3 | 1 | 2 | 1 | |||
| B | 52 | 2 | 2 | 2 | 6 | 3 | 1 | ||
| B | 57 | 1 | 3 | ||||||
| A | 58 | 1 | 3 | 3 | |||||
| B | 60 | 1 | 3 | 5 | 1 | ||||
| B | 64 | 1 | 2 | 4 | 3 | ||||
| B | 65 | 1 | 3 | ||||||
| B | 67 | 4 | 3 | 1 | 3 | ||||
| VNT Titers | 0 | 16 | 32 | 64 | 128 | 256 | |
|---|---|---|---|---|---|---|---|
| zone | dep | ||||||
| B | 21* | 6* | |||||
| / | 2A | 9 | 2 | 1 | |||
| / | 2B | 13 | 7 | 7 | 4 | 2 | 1 |
| A | 36* | 38* | |||||
| A | 41* | 5* | |||||
| B | 52* | 3* | |||||
| B | 64 | 1 | 2 | 1 | 1 | ||
| Year/VNT Titers | 0 | 4 | 8 | 16 | 32 | 64 | 128 | 256 |
|---|---|---|---|---|---|---|---|---|
| 2008–2009 | 14 | 23 | 23 | 9 | 1 | |||
| 2010–2011 | 2 | 3 | 17 | 30 | 10 | 5 | 4 | |
| 2011–2012 | 10 | 9 | 20 | 31 | 18 | 11 | 3 | |
| 2012–2013 | 1 | 3 | 10 | 8 | 11 | 3 | ||
| 2013–2014 | 4 | 3 | 5 | 5 | ||||
| 2014–2015 | 2 | 7 | 6 | 4 | 1 | |||
| 2015–2016 | 4 | 3 | 6 | 2 |
| Species | Collections in Natural Zones | Collections in Farms | All Locations | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alsace Region (10 Sites, 11 Collections) | Bourgogne Region (10 Sites, 11 Collections) | Alps Region (5 Sites, 6 Collections) | Haut-Languedoc Region (8 Sites, 26 Collections) | Pyrenees Region (1 site, 1 Collection) | Total | Rank | Alsace Region (1 site, 85 Collections) | Bourgogne Region (1 site, 92 Collections) | Alps Region (1 site, 50 Collections) | Haut-Languedoc Region (1 site, 100 Collections) | Pyrenees Region (1 site, 95 Collections) | Total | Rank | Weighted Average Collectionb | Rank | Cumulative Percentage | |
| C. obsoletus/C. scoticus | 29,774a | 435 | 34 | 75 | 71 | 30,389 | 1 | 3990 | 634 | 3661 | 2275 | 844 | 11,403 | 1 | 1587.2 | 1 | 70.55% |
| C. furcillatus | 9087 | 0 | 1 | 4 | 2 | 9094 | 2 | 50 | 3 | 66 | 51 | 7 | 176 | 8 | 151.4 | 2 | 77.28% |
| C. achrayi | 2787 | 1 | 0 | 1226 | 6 | 4020 | 3 | 20 | 34 | 10 | 376 | 0 | 441 | 3 | 103.2 | 3 | 81.86% |
| C. chiopterus | 20 | 8 | 1 | 0 | 0 | 29 | 19 | 98 | 600 | 11 | 20 | 6 | 735 | 2 | 73.9 | 4 | 90.40% |
| C. punctatus | 269 | 99 | 1 | 2692 | 83 | 3144 | 4 | 32 | 10 | 32 | 24 | 5 | 102 | 11 | 56.5 | 5 | 84.37% |
| C. pulicaris | 112 | 52 | 1 | 56 | 0 | 221 | 9 | 175 | 5 | 50 | 131 | 31 | 391 | 4 | 42.4 | 6 | 93.83% |
| C. lupicaris | 363 | 3 | 2 | 10 | 40 | 418 | 8 | 202 | 2 | 18 | 23 | 44 | 287 | 5 | 34.8 | 7 | 91.95% |
| C. pallidicornis | 1653 | 0 | 0 | 0 | 4 | 1657 | 5 | 15 | 0 | 0 | 19 | 8 | 42 | 15 | 28.6 | 8 | 85.65% |
| C. dewulfi | 14 | 34 | 0 | 5 | 4 | 57 | 16 | 71 | 44 | 1 | 5 | 95 | 216 | 6 | 22.4 | 9 | 94.83% |
| C. vexans | 0 | 0 | 0 | 51 | 0 | 51 | 17 | 2 | 2 | 204 | 3 | 0 | 211 | 7 | 21.8 | 10 | 95.80% |
| C. kibunensis | 720 | 6 | 12 | 6 | 10 | 754 | 6 | 35 | 4 | 24 | 15 | 2 | 78 | 12 | 18.9 | 11 | 86.49% |
| C. odiatus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 151 | 12 | 4 | 167 | 9 | 16.7 | 12 | 97.30% | |
| C. grisescens | 0 | 0 | 0 | 0 | 0 | 0 | 149 | 0 | 0 | 0 | 0 | 149 | 10 | 14.9 | 13 | 97.96% | |
| C. festivipennis | 481 | 264 | 0 | 3 | 2 | 750 | 7 | 6 | 14 | 4 | 3 | 5 | 31 | 18 | 14.1 | 14 | 87.11% |
| C. delta | 24 | 0 | 10 | 82 | 5 | 121 | 12 | 2 | 1 | 3 | 44 | 0 | 50 | 13 | 6.8 | 15 | 96.34% |
| C. fascipennis | 53 | 0 | 0 | 102 | 0 | 155 | 10 | 0 | 0 | 3 | 28 | 1 | 32 | 16 | 5.5 | 16 | 96.04% |
| C. subfasciipennis | 137 | 0 | 1 | 1 | 0 | 139 | 11 | 7 | 1 | 0 | 14 | 7 | 29 | 21 | 4.9 | 17 | 96.56% |
| C. poperinghensis | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 15 | 19 | 11 | 48 | 14 | 4.8 | 18 | 98.67% | |
| C. newsteadi | 0 | 17 | 0 | 0 | 0 | 17 | 20 | 2 | 6 | 16 | 4 | 3 | 31 | 18 | 3.3 | 19 | 98.46% |
| C. flavipulicaris | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 32 | 0 | 0 | 32 | 16 | 3.2 | 20 | 99.04% | |
| C. nubeculosus | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 0 | 15 | 2 | 2 | 31 | 18 | 3.1 | 21 | 99.17% | |
| C. brunnicans | 0 | 0 | 0 | 8 | 0 | 8 | 25 | 1 | 8 | 2 | 7 | 8 | 26 | 22 | 2.7 | 22 | 98.89% |
| C. fagineus | 38 | 0 | 0 | 0 | 0 | 38 | 18 | 0 | 0 | 0 | 20 | 0 | 20 | 25 | 2.6 | 23 | 98.31% |
| C. circumscriptus | 0 | 0 | 0 | 1 | 0 | 1 | 29 | 0 | 0 | 16 | 8 | 1 | 25 | 23 | 2.5 | 24 | 99.28% |
| C. picturatus | 12 | 0 | 0 | 1 | 0 | 13 | 21 | 1 | 2 | 13 | 4 | 2 | 22 | 24 | 2.3 | 25 | 98.78% |
| C. parroti | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 4 | 0 | 0 | 7 | 20 | 25 | 2.0 | 26 | 99.43% | |
| C. begueti | 0 | 0 | 0 | 0 | 105 | 105 | 13 | 0 | 3 | 1 | 0 | 0 | 4 | 38 | 1.9 | 27 | 98.05% |
| C. sejfadinei/C. tauricus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 | 0 | 0 | 19 | 27 | 1.9 | 28 | 99.52% | |
| C. impunctatus | 9 | 0 | 0 | 61 | 0 | 70 | 15 | 2 | 0 | 0 | 6 | 0 | 8 | 34 | 1.8 | 29 | 98.20% |
| C. clastrieri | 84 | 0 | 0 | 0 | 0 | 84 | 14 | 0 | 2 | 0 | 1 | 1 | 4 | 38 | 1.6 | 30 | 98.12% |
| C. simulator | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 3 | 0 | 0 | 15 | 28 | 1.5 | 31 | 99.65% | |
| C. alazanicus | 0 | 1 | 0 | 0 | 0 | 1 | 29 | 0 | 1 | 0 | 12 | 1 | 14 | 29 | 1.4 | 32 | 99.71% |
| C. segnis | 1 | 0 | 0 | 3 | 0 | 4 | 27 | 2 | 1 | 9 | 2 | 0 | 14 | 29 | 1.4 | 33 | 99.58% |
| C. cataneii/C. gejgelensis | 0 | 11 | 0 | 0 | 0 | 11 | 22 | 0 | 0 | 3 | 8 | 1 | 12 | 31 | 1.4 | 34 | 99.35% |
| C. longipennis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 11 | 32 | 1.1 | 35 | 99.76% | |
| C. tauricus | 0 | 0 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 11 | 32 | 1.1 | 35 | 99.81% | |
| C. subfagineus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 1 | 0 | 6 | 35 | 0.6 | 37 | 99.84% | |
| C. pictipennis | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 0 | 2 | 6 | 35 | 0.6 | 38 | 99.87% | |
| C. maritimus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 5 | 37 | 0.5 | 39 | 99.89% | |
| C. duddingstoni | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 4 | 38 | 0.4 | 40 | 99.91% | |
| C. stigma | 1 | 0 | 0 | 0 | 0 | 1 | 29 | 2 | 1 | 0 | 0 | 0 | 3 | 41 | 0.3 | 41 | 99.92% |
| C. minutissimus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 3 | 41 | 0.3 | 42 | 99.95% | |
| C. riethi | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 3 | 41 | 0.3 | 43 | 99.96% | |
| C. dendriticus | 0 | 0 | 0 | 2 | 0 | 2 | 28 | 0 | 0 | 0 | 2 | 0 | 2 | 44 | 0.2 | 44 | 99.93% |
| C. puncticollis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 44 | 0.2 | 45 | 99.97% | |
| C. salinarius | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 44 | 0.2 | 45 | 99.98% | |
| C. santonicus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 44 | 0.2 | 47 | 99.98% | |
| C. clintoni | 10 | 0 | 0 | 0 | 0 | 10 | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 48 | 99.81% | |
| C. cameroni | 0 | 0 | 7 | 0 | 0 | 7 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 49 | 99.82% | |
| C. jumineri | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 48 | 0.1 | 50 | 99.99% | |
| C. gornostaevae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 48 | 0.1 | 50 | 99.99% | |
| Culicoides sp. | 0 | 0 | 7 | 2 | 0 | 9 | 0 | 0 | 0 | 0 | 1 | 1 | 0.2 | 100.00% | |||
| Total | 45,649 | 931 | 77 | 4391 | 332 | 51,380 | 4903 | 1399 | 4399 | 3141 | 1099 | 14,942 | 2250 | ||||
| Collections in Natural Areas | Collections in Farms | |||||||
|---|---|---|---|---|---|---|---|---|
| WCFa | WCBa | DHMa | WMMa | WCFa | WCBa | DHMa | WMMa | |
| No. collections (sites x trapping) | 12 | 10 | 5 | 18 | 50 | 67 | 25 | 136 |
| Observed no. species | 21 | 12 | 10 | 21 | 29 | 27 | 30 | 36 |
| Chao expected no. species ± s.e.b | 24.67 ± 4.88 | 12.60 ± 1.20 | 19.80 ± 10.62 | 26.90 ± 7.16 | 33.08 ± 4.79 | 39.31 ±10.53 | 57.00 ± 20.09 | 41.03 ± 4.41 |
| Fisher α of log series | 2.103 | 1.944 | 3.193 | 2.830 | 4.091 | 4.761 | 4.343 | 5.441 |
| Berger-Parker dominance index | 0.652 | 0.467 | 0.486 | 0.588 | 0.814 | 0.444 | 0.830 | 0.724 |
| Evenness index | 0.377 | 0.590 | 0.680 | 0.425 | 0.271 | 0.387 | 0.258 | 0.357 |
| Species | No. Blood-Fed Females | No. Positive for Vertebrate Blood | No. Females Found Fed on | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red Deer Cervus elafus | Roe Deer Apreolus capreolus | Chamois Rupicapra rupicapra | Cattle Bos taurus | Goat Capra hircus | Sheep or Mouflon Ovis aries | Horse Equus caballus | Pig or Wild Boar Sus scrofa | Cat Felis catus | Dog Canis lupus familaris | Birds Sylvia genus | |||
| C. achrayi | 15 | 14 | 6 | 1 | 2 | 1 | |||||||
| C. dewulfi | 2 | 2 | 2 | ||||||||||
| C. fascipennis | 1 | 1 | |||||||||||
| C. festivipennis | 2 | 2 | |||||||||||
| C. kibunensis | 2 | 2 | 1 | 1 | |||||||||
| C. lupicaris | 4 | 3 | |||||||||||
| C. obsoletus/C. scoticus | 6 | 5 | 1 | 1 | 1 | ||||||||
| C. obsoletus | 16 | 13 | 2 | 1 | 2 | 1 | |||||||
| C. scoticus | 2 | 2 | 1 | ||||||||||
| C. pulicaris | 1 | 1 | 1 | ||||||||||
| C. punctatus | 10 | 7 | 1 | 1 | 3 | ||||||||
| C. segnis | 1 | 1 | 1 | ||||||||||
| C. vexans | 8 | 7 | 3 | 1 | |||||||||
| Total | 70 | 60 | 9 | 2 | 1 | 10 | 0 | 6 | 5 | 0 | 0 | 0 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, S.; Balenghien, T.; Viarouge, C.; Faure, E.; Zanella, G.; Sailleau, C.; Mathieu, B.; Delécolle, J.-C.; Ninio, C.; Garros, C.; et al. Red deer (Cervus elaphus) Did Not Play the Role of Maintenance Host for Bluetongue Virus in France: The Burden of Proof by Long-Term Wildlife Monitoring and Culicoides Snapshots. Viruses 2019, 11, 903. https://doi.org/10.3390/v11100903
Rossi S, Balenghien T, Viarouge C, Faure E, Zanella G, Sailleau C, Mathieu B, Delécolle J-C, Ninio C, Garros C, et al. Red deer (Cervus elaphus) Did Not Play the Role of Maintenance Host for Bluetongue Virus in France: The Burden of Proof by Long-Term Wildlife Monitoring and Culicoides Snapshots. Viruses. 2019; 11(10):903. https://doi.org/10.3390/v11100903
Chicago/Turabian StyleRossi, Sophie, Thomas Balenghien, Cyril Viarouge, Eva Faure, Gina Zanella, Corinne Sailleau, Bruno Mathieu, Jean-Claude Delécolle, Camille Ninio, Claire Garros, and et al. 2019. "Red deer (Cervus elaphus) Did Not Play the Role of Maintenance Host for Bluetongue Virus in France: The Burden of Proof by Long-Term Wildlife Monitoring and Culicoides Snapshots" Viruses 11, no. 10: 903. https://doi.org/10.3390/v11100903
APA StyleRossi, S., Balenghien, T., Viarouge, C., Faure, E., Zanella, G., Sailleau, C., Mathieu, B., Delécolle, J. -C., Ninio, C., Garros, C., Gardès, L., Tholoniat, C., Ariston, A., Gauthier, D., Mondoloni, S., Barboiron, A., Pellerin, M., Gibert, P., Novella, C., ... Bréard, E. (2019). Red deer (Cervus elaphus) Did Not Play the Role of Maintenance Host for Bluetongue Virus in France: The Burden of Proof by Long-Term Wildlife Monitoring and Culicoides Snapshots. Viruses, 11(10), 903. https://doi.org/10.3390/v11100903