Pathogenicity and Transmissibility of North American H7 Low Pathogenic Avian Influenza Viruses in Chickens and Turkeys


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
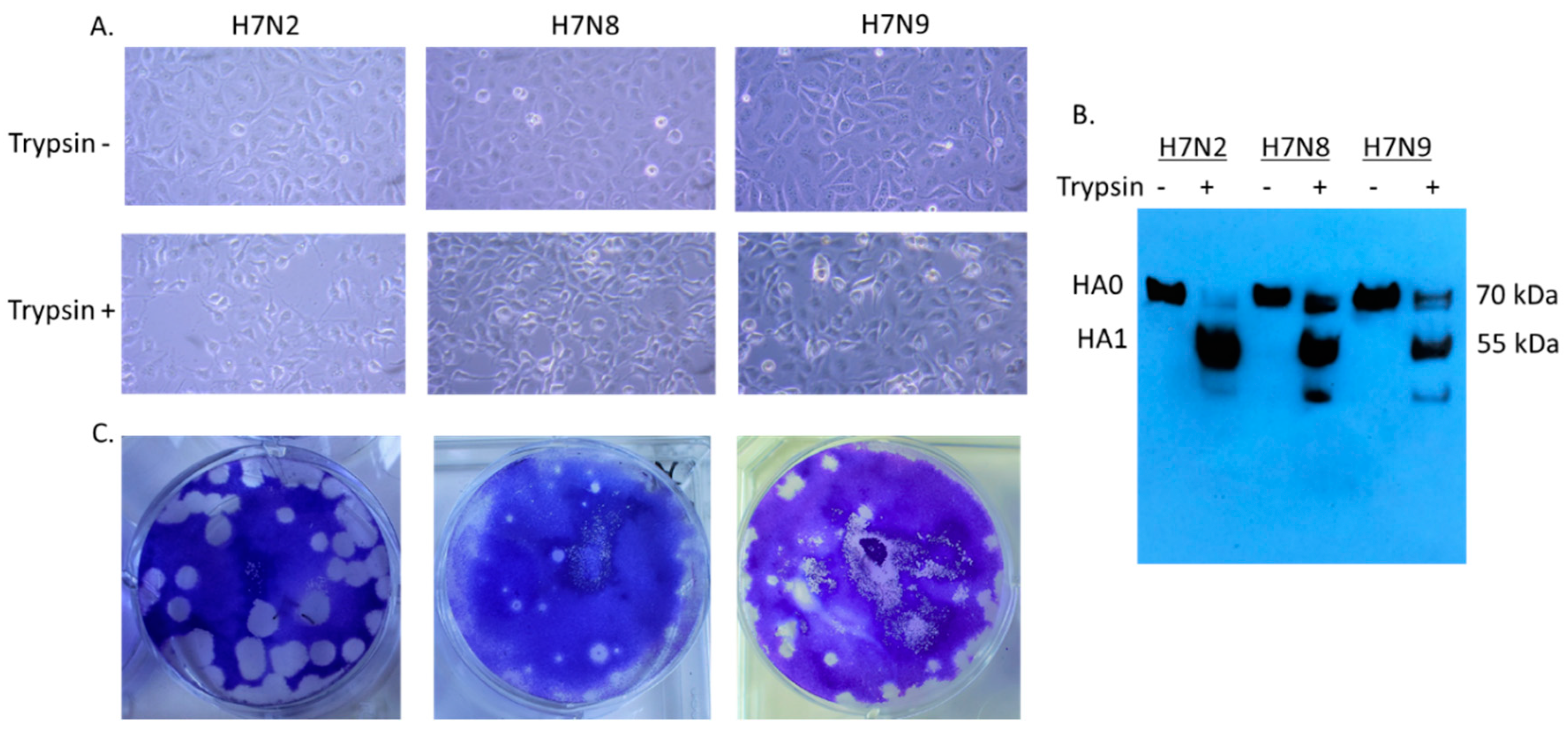
2.2. Cytopathic Effect of H7 Viruses and the HA Cleavability in MDCK Cells
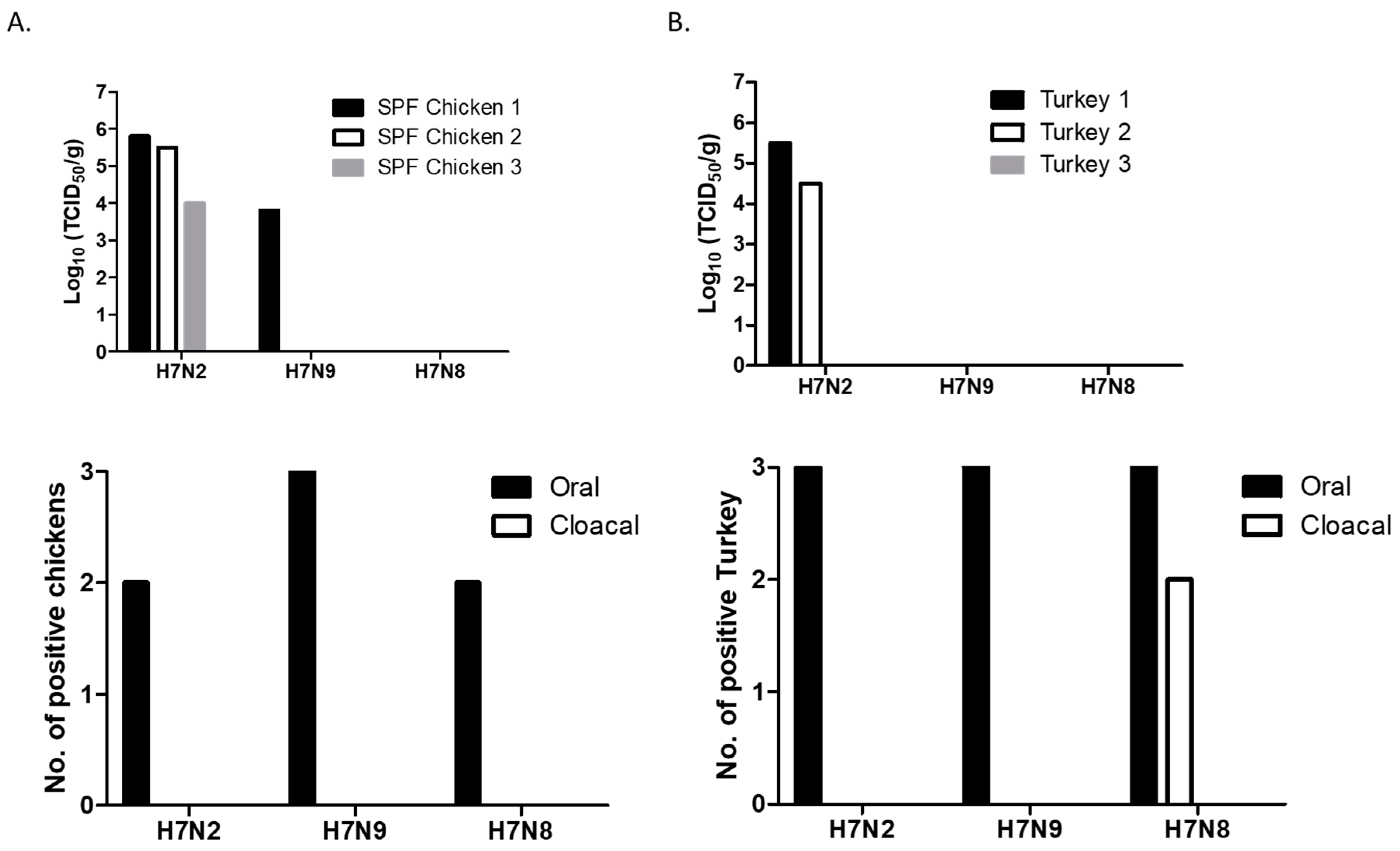
2.3. Replication of H7 Viruses in 2-Week-Old Chickens and Turkeys
2.4. Replication and Transmission of H7N2 and H7N9 Viruses in 4-Week-Old SPF and Broiler Chickens
2.5. Statistical Analysis
3. Results
3.1. In Vitro Characterization of H7 Viruses
3.2. Replication of H7 Viruses Was Restricted to Upper Respiratory Tract in 2-Week-Old Chickens and Turkeys
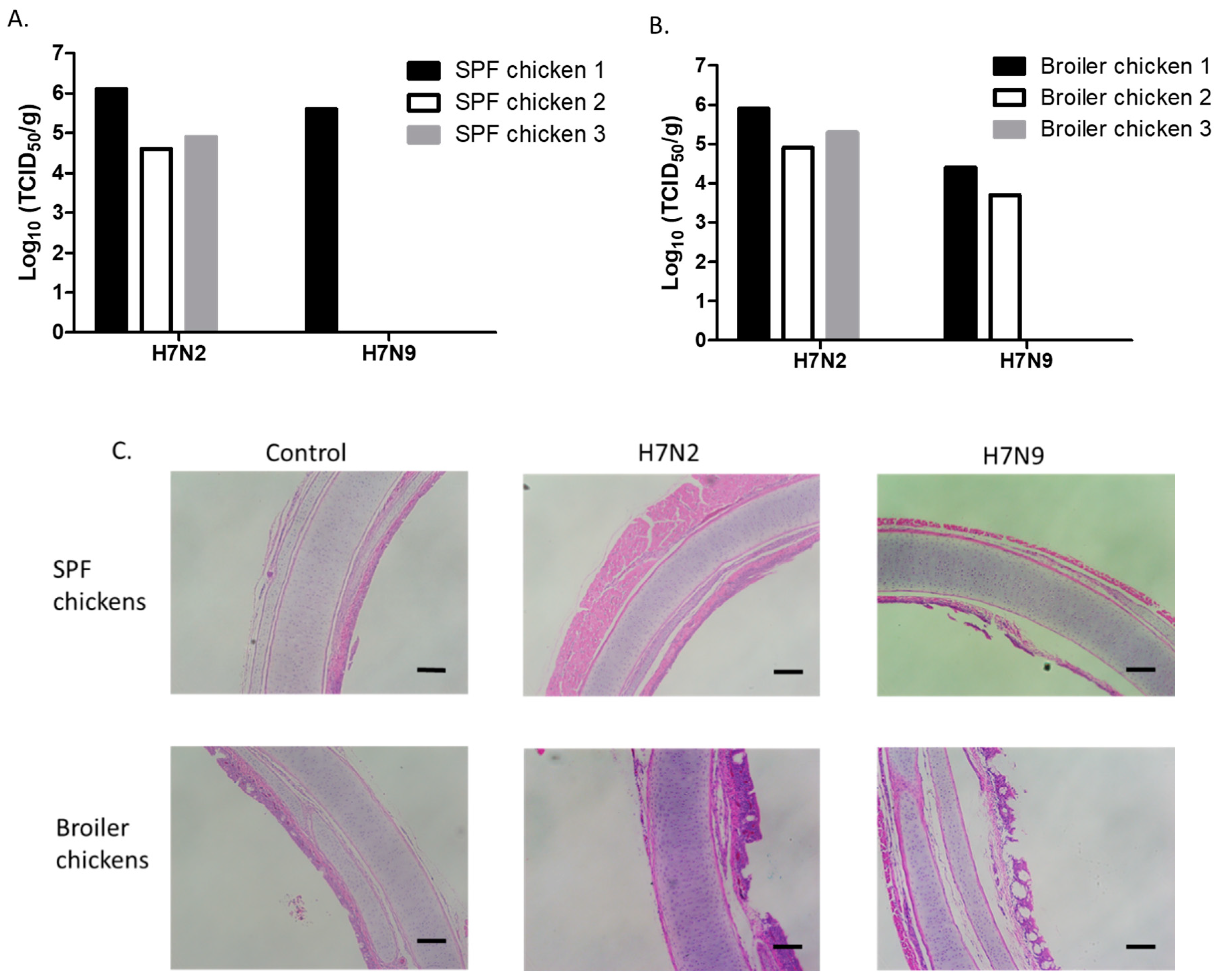
3.3. Attenuation of H7N9 in 4-Week-Old SPF and Broiler Chickens
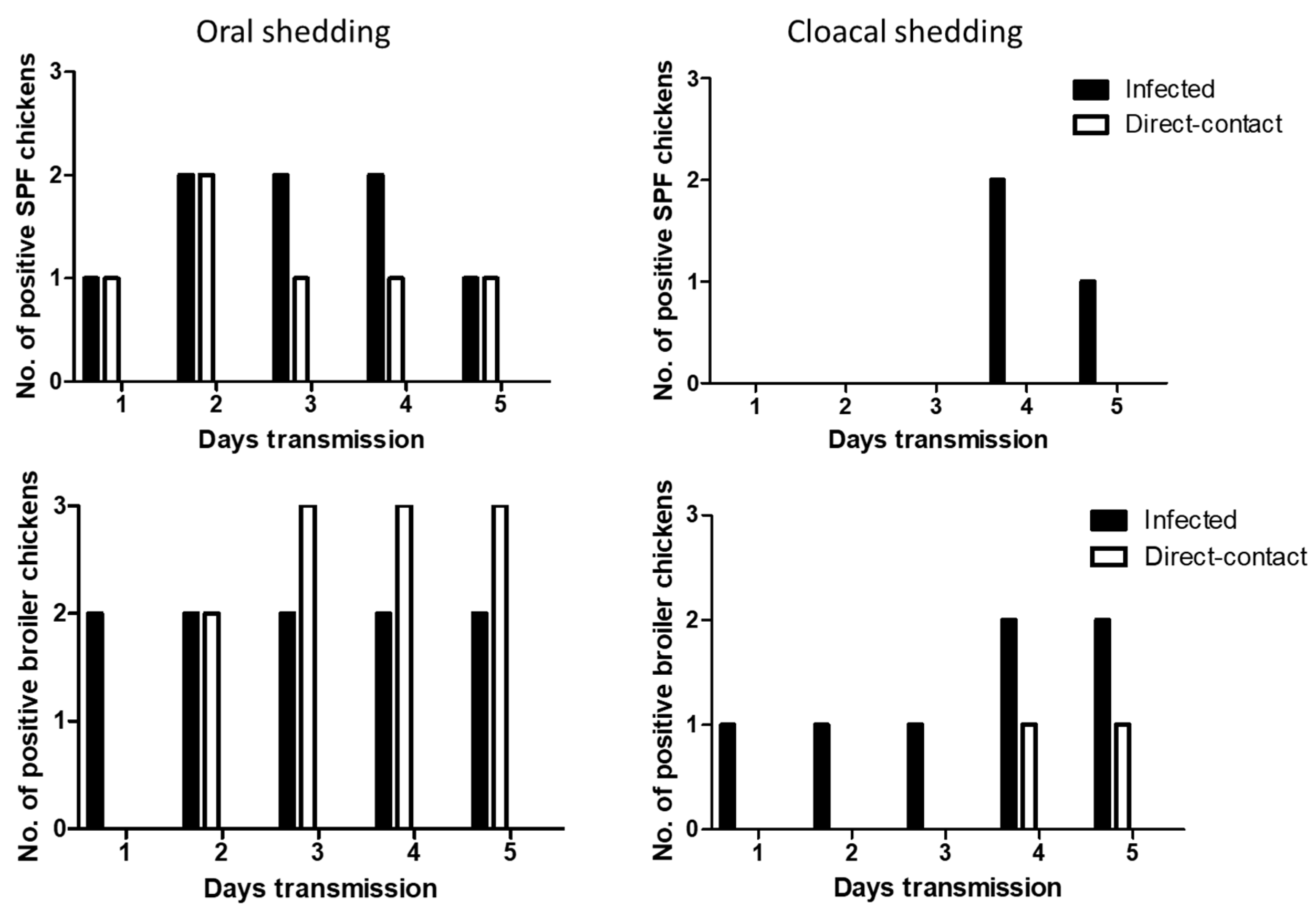
3.4. Efficient Seroconversion of Chickens by Direct-Contact Transmission of H7 Viruses
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Palese, P.; Shaw, M.L. Orthomyxoviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1647–1689. [Google Scholar]
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N1. Virus Res. 2013, 178, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.J. Summary of avian influenza activity in Europe, Asia, Africa, and Australasia, 2002–2006. Avian Dis. 2007, 51, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Kawaoka, Y.; Bean, W.J. What is the potential of avirulent influenza viruses to complement a cleavable hemagglutinin and generate virulent strains. Virology 1989, 171, 484–492. [Google Scholar] [CrossRef]
- Webster, R.G.; Govorkova, E.A. H5N1 influenza—Continuing evolution and spread. N. Engl. J. Med. 2006, 355, 2174–2177. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Cumulative Number of Confirmed Human Cases of Avian Influenza A (H5N1) Reported to WHO, 2003–2017; World Health Organization: Geneva, Switzerland. Available online: http://www.who.int/influenza/human_animal_interface/2017_02_14_tableH5N1.pdf?ua=1 (accessed on 26 December 2018).
- Abdelwhab, E.M.; Veits, J.; Mettenleiter, T.C. Prevalence and control of H7 avian influenza viruses in birds and humans. Epidemiol. Infect. 2014, 142, 896–920. [Google Scholar] [CrossRef] [PubMed]
- Monne, I.; Fusaro, A.; Nelson, M.I.; Bonfanti, L.; Mulatti, P.; Hughes, J.; Murcia, P.R.; Schivo, A.; Valastro, V.; Moreno, A.; et al. Emergence of a highly pathogenic avian influenza virus from a low-pathogenic progenitor. J. Virol. 2014, 88, 4375–4388. [Google Scholar] [CrossRef] [PubMed]
- Seekings, A.H.; Slomka, M.J.; Russell, C.; Howard, W.A.; Choudhury, B.; Nuñéz, A.; Löndt, B.Z.; Cox, W.; Ceeraz, V.; Thorén, P.; et al. Direct evidence of H7N7 avian influenza virus mutation from low to high virulence on a single poultry premises during an outbreak in free range chickens in the UK, 2008, Infect. Genet. Evol. 2018, 64, 13–31. [Google Scholar] [CrossRef]
- Krauss, S.; Stucker, K.M.; Schobel, S.A.; Danner, A.; Friedman, K.; Knowles, J.P.; Kayali, G.; Niles, L.J.; Dey, A.D.; Raven, G. Long-term surveillance of H7 influenza viruses in American wild aquatic birds: Are the H7N3 influenza viruses in wild birds the precursors of highly pathogenic strains in domestic poultry? Emerg. Microbes Infect. 2015, 4, e35. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.L.; Kim-Torchetti, M.; Hines, N.; Yingst, S.; DeLiberto, T.; Lee, D.-H. Outbreak of H7N8 low pathogenic avian influenza in commercial turkeys with spontaneous mutation to highly pathogenic avian influenza. Genome Announc. 2016, 4, e00457-16. [Google Scholar] [CrossRef]
- Xu, Y.; Ramey, A.M.; Bowman, A.S.; Deliberto, T.J.; Killian, M.L.; Krauss, S.; Nolting, J.M.; Torchetti, M.K.; Reeves, A.B.; Webby, R.J.; et al. Low-pathogenic influenza A viruses in North American diving ducks contribute to the emergence of a novel highly pathogenic influenza A(H7N8) virus. J. Virol. 2017, 91, e02208-16. [Google Scholar] [CrossRef]
- Lee, D.H.; Torchetti, M.K.; Killian, M.L.; Berhane, Y.; Swayne, D.E. Highly pathogenic avian influenza A(H7N9) virus, Tennessee, USA, March 2017. Emerg. Infect. Dis. 2017, 23, 1860–1863. [Google Scholar] [CrossRef] [PubMed]
- Ladman, B.S.; Rosenberger, S.C.; Rosenberger, J.K.; Pope, C.R.; Gelb, J., Jr. Virulence of low pathogenicity H7N2 avian influenza viruses from the Delmarva peninsula for broiler and leghorn chickens and turkeys. Avian Dis. 2008, 52, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.; Matrosovich, T.; Garten, W.; Klenk, H.-D. New low-viscosity overlay medium for viral plaque assays. Virol. J. 2006, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method for estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Davison, S.; Eckroade, R.J.; Ziegler, A.F. A review of the 1996-98 nonpathogenic H7N2 avian influenza outbreak in Pennsylvania. Avian Dis. 2003, 47, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A.; Suarez, D.L.; Stallnecht, D.E.; Pedersen, J.C.; Panigrahy, B. Ecology and epidemiology of avian influenza in North and South America. Dev. Biol. 2006, 124, 37–44. [Google Scholar]
- OIE (World Organisation for Animal Health) Avian Influenza. 2015. Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.03.04_AI.pdf (accessed on 26 December 2018).
- Tumpey, T.M.; Kapczynski, D.R.; Swayne, D.E. Comparative susceptibility of chickens and turkeys to avian influenza A H7N2 virus infection and protective efficacy of a commercial avian influenza H7N2 virus vaccine. Avian Dis. 2004, 48, 167–176. [Google Scholar] [CrossRef]
- Swayne, D.E. The global nature of avian influenza. In Avian Influenza, 1st ed.; Swayne, D.E., Ed.; Blackwell: Ames, IA, USA, 2008; pp. 123–143. [Google Scholar]
- Kawaoka, Y.; Webster, R.G. Evolution of the A/Chicken/Pennsylvania/83 (H5N2) influenza virus. Virology 1985, 146, 130–137. [Google Scholar] [CrossRef]
- Wester, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar]
- Swayne, D.E. Impact of vaccines and vaccination on global control of avian influenza. Avian Dis. 2012, 56, 818–828. [Google Scholar] [CrossRef]
- Bertran, K.; Balzli, C.; Lee, D.H.; Suarez, D.L.; Kapczynski, D.R.; Swayne, D.E. Protection of white leghorn chickens by U.S. emergency H5 vaccination against clade 2.3.4.4 H5N2 high pathogenicity avian influenza virus. Vaccine 2017, 35, 6336–6344. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E. Avian influenza vaccine use during 2001. In Proceedings of the 104th Annual Meeting of the U.S. Animal Health Association, Hershey, PA, USA, 9–14 October 2001; USAHA: Richmond, VA, USA, 2001; pp. 469–471. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy Chowdhury, I.; Yeddula, S.G.R.; Kim, S.-H. Pathogenicity and Transmissibility of North American H7 Low Pathogenic Avian Influenza Viruses in Chickens and Turkeys. Viruses 2019, 11, 163. https://doi.org/10.3390/v11020163
Roy Chowdhury I, Yeddula SGR, Kim S-H. Pathogenicity and Transmissibility of North American H7 Low Pathogenic Avian Influenza Viruses in Chickens and Turkeys. Viruses. 2019; 11(2):163. https://doi.org/10.3390/v11020163
Chicago/Turabian StyleRoy Chowdhury, Ishita, Sai Goutham Reddy Yeddula, and Shin-Hee Kim. 2019. "Pathogenicity and Transmissibility of North American H7 Low Pathogenic Avian Influenza Viruses in Chickens and Turkeys" Viruses 11, no. 2: 163. https://doi.org/10.3390/v11020163
APA StyleRoy Chowdhury, I., Yeddula, S. G. R., & Kim, S. -H. (2019). Pathogenicity and Transmissibility of North American H7 Low Pathogenic Avian Influenza Viruses in Chickens and Turkeys. Viruses, 11(2), 163. https://doi.org/10.3390/v11020163