Global Transcriptomic Analysis Reveals Insights into the Response of ‘Etrog’ Citron (Citrus medica L.) to Citrus Exocortis Viroid Infection

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
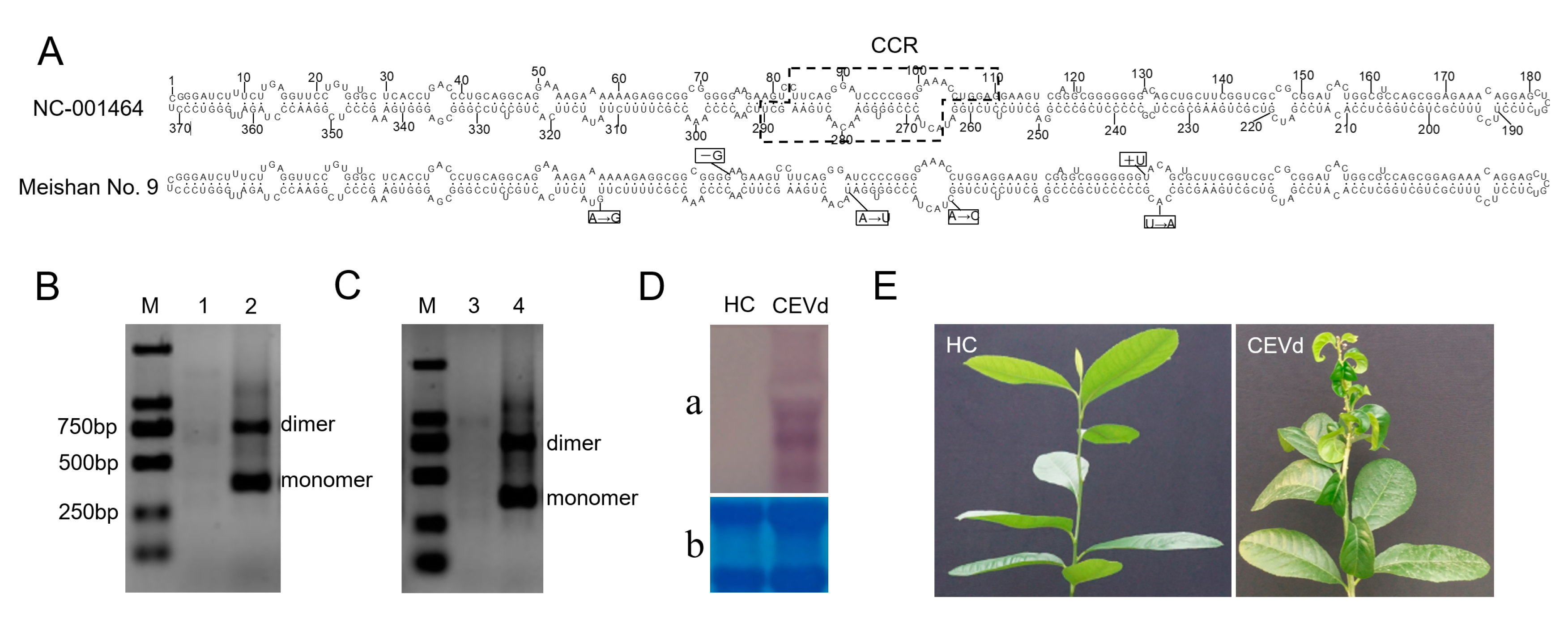
2.1. Preparation of Infectious CEVd RNAs
2.2. Inoculation of the Viroid-Free Plants
2.3. Northern Blot Hybridization
2.4. Library Construction and Transcriptome Sequencing
2.5. Identification and Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.6. Validation of DEGs by RT-qPCR
3. Results
3.1. Infectivity Confirmation of CEVd RNAs
3.2. Transcriptome Sequencing and Gene Expression Analysis
3.3. Gene Enrichment Analysis
3.4. CEVd Infection Induces Expression of Many Genes That Are Related to Basal Defense Responses
3.5. CEVd Infection Impacts Plant Hormone Signaling
3.6. RNA Silencing Responses to CEVd Infection
3.7. Validation of RNA-seq Results by RT-qPCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flores, R.; Minoia, S.; Carbonell, A.; Gisel, A.; Delgado, S.; López-Carrasco, A.; Navarro, B.; Serio, F.D. Viroids, the simplest RNA replicons: How they manipulate their hosts for being propagated and how their hosts react for containing the infection. Virus Res. 2015, 209, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Serra, P.; Messmer, A.; Sanderson, D.; James, D.; Flores, R. Apple hammerhead viroid-like RNA is a bona fide viroid: autonomous replication and structural features support its inclusion as a new member in the genus Pelamoviroid. Virus Res. 2018, 249, 8–15. [Google Scholar] [CrossRef]
- Duranvila, N. Viroids as companions of a professional career. Viruses 2019, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Hernández, C.; Martínez de Alba, A.E.; Daròs, J.A.; Serio, F.D. Viroids and viroid-host interactions. Annu. Rev. Phytopathol. 2005, 43, 117–139. [Google Scholar] [CrossRef]
- Ding, S.W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef]
- Itaya, A.; Folimonov, A.; Matsuda, Y.; Nelson, R.S.; Ding, B. Potato spindle tuber viroid as inducer of RNA silencing in infected tomato. Mol. Plant Microbe Interact. 2001, 14, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Papaefthimiou, I.; Hamilton, A.; Denti, M.; Baulcombe, D.; Tsagris, M.; Tabler, M. Replicating potato spindle tuber viroid RNA is accompanied by short RNA fragments that are characteristic of post-transcriptional gene silencing. Nucleic Acids Res. 2001, 29, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyko, A.; Kathiria, P.; Zemp, F.J.; Yao, Y.; Pogribny, I.; Kovalchuk, I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants: (virus-induced plant genome instability). Nucleic Acids Res. 2007, 35, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.; Griffiths, J.S.; Huang, T.S.; Wang, A. Altered gene expression changes in Arabidopsis leaf tissues and protoplasts in response to Plum pox virus infection. BMC Genomics 2008, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tessitori, M.; Maria, G.; Capasso, C.; Catara, G.; Rizza, S.; de Luca, V.; Catara, A.; Capasso, A.; Carginale, V. Differential display analysis of gene expression in Etrog citron leaves infected by citrus viroid III. Biochim. Biophys. Acta 2007, 1769, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Owens, R.A.; Tech, K.B.; Shao, J.Y.; Sano, T.; Baker, C.J. Global analysis of tomato gene expression during potato spindle tuber viroid infection reveals a complex array of changes affecting hormone signaling. Mol. Plant Microbe Interact. 2012, 25, 582–598. [Google Scholar] [CrossRef]
- Rizza, S.; Conesa, A.; Juarez, J.; Catara, A.; Navarro, L.; Duran-Vila, N.; Ancillo, G. Microarray analysis of etrog citron (Citrus medica L.) reveals changes in chloroplast, cell wall, peroxidase and symporter activities in response to viroid infection. Mol. Plant Pathol. 2012, 13, 852–864. [Google Scholar] [CrossRef]
- Itaya, A.; Matsuda, Y.; Gonzales, R.A.; Nelson, R.S.; Ding, B. Potato spindle tuber viroid strains of different pathogenicity induces and suppresses expression of common and unique genes in infected tomato. Mol. Plant Microbe Interact. 2002, 15, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shibuya, M.; Taneda, A.; Kurauchi, T.; Senda, M.; Owens, R.A.; Sano, T. Accumulation of Potato spindle tuber viroid-specific small RNAs is accompanied by specific changes in gene expression in two tomato cultivars. Virology 2011, 413, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Marguerat, S.; Bahler, J. RNA-seq: From technology to biology. Cell. Mol. Life Sci. 2010, 67, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Van Verk, M.C.; Hickman, R.; Pieterse, C.M.; Van Wees, S.C. RNA-Seq: revelation of the messengers. Trends Plant Sci. 2013, 18, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Katsarou, K.; Wu, Y.; Zhang, R.; Bonar, N.; Morris, J.; Hedley, P.E.; Bryan, G.J.; Kalantidis, K.; Hornyik, C. Insight on genes affecting tuber development in potato upon potato spindle tuber viroid (PSTVd) infection. PLoS ONE 2016, 11, e0150711. [Google Scholar] [CrossRef] [PubMed]
- Herranz, M.C.; Niehl, A.; Rosales, M.; Fiore, N.; Zamorano, A.; Granell, A.; Pallas, V. A remarkable synergistic effect at the transcriptomic level in peach fruits doubly infected by prunus necrotic ringspot virus and peach latent mosaic viroid. Virol. J. 2013, 10, 164. [Google Scholar] [CrossRef]
- Kappagantu, M.; Bullock, J.M.; Nelson, M.E.; Eastwell, K.C. Hop stunt viroid: Effect on host (Humulus lupulus) transcriptome and its interactions with hop powdery mildew (Podospheara macularis). Mol. Plant Microbe Interact. 2017, 30, 842–851. [Google Scholar] [CrossRef]
- Xia, C.; Li, S.; Hou, W.; Fan, Z.; Xiao, H.; Lu, M.; Sano, T.; Zhang, Z. Global transcriptomic changes induced by infection of cucumber (Cucumis sativus L.) with mild and severe variants of hop stunt viroid. Front Microbiol. 2017, 12, 2427. [Google Scholar] [CrossRef]
- Pokorn, T.; Radišek, S.; Javornik, B.; Štajner, N.; Jakše, J. Development of hop transcriptome to support research into host-viroid interactions. PLoS ONE 2017, 12, e0184528. [Google Scholar] [CrossRef]
- Duranvila, N.; Roistacher, C.N.; Riverabustamante, R.; Semancik, J.S. A definition of citrus viroid groups and their relationship to the exocortis disease. J. Gen. Virol. 1988, 69, 3069–3080. [Google Scholar] [CrossRef]
- Verniere, C.; Perrier, X.; Dubois, C.; Dubois, A.; Botella, L.; Chabrier, C.; Bove, J.M.; Duranvila, N. Citrus viroids: symptom expression and effect on vegetative growth and yield of clementine trees grafted on trifoliate orange. Plant Dis. 2004, 88, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Verniere, C.; Perrier, X.; Dubois, C.; Dubois, A.; Botella, L.; Chabrier, C.; Bove, J.M.; Duranvila, N. Interactions between citrus viroids affect symptom expression and field performance of Clementine trees grafted on trifoliate orange. Phytopathology 2006, 96, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Murcia, N.; Hashemian, S.M.B.; Serra, P.; Pina, J.A.; Duranvila, N. Citrus viroids: symptom expression and performance of washington navel sweet orange trees grafted on carrizo citrange. Plant Dis. 2015, 99, 125–136. [Google Scholar] [CrossRef]
- Roistacher, C.N.; Calavan, E.C.; Blue, R.L.; Navarro, L.; Gonzales, R. A new more sensitive citron indicator for detection of mild isolates of citrus exocortis viroid (CEV). Plant Dis. Rep. 1977, 61, 135–139. [Google Scholar]
- Mishra, M.D.; Hammond, R.W.; Owens, R.A.; Smith, D.R.; Diener, T.O. Indian bunchy top disease of tomato plants is caused by a distinct strain of citrus exocortis viroid. J. Gen. Virol. 1991, 72, 1781. [Google Scholar] [CrossRef]
- Jthj, V.; Ccc, J.; Willemen, T.M.; Lff, K.; Owens, R.A.; Roenhorst, J.W. Natural infections of tomato by Citrus exocortis viroid, Columnea latent viroid, Potato spindle tuber viroid and Tomato chlorotic dwarf viroid. Eur. J. Plant Pathol. 2004, 110, 823–831. [Google Scholar]
- Bernad, L.; Duranvila, N.; Elena, S.F. Effect of citrus hosts on the generation, maintenance and evolutionary fate of genetic variability of citrus exocortis viroid. J. Gen. Virol. 2009, 90, 2040. [Google Scholar] [CrossRef]
- Bernad, L.; Duranvila, N. A novel RT-PCR approach for detection and characterization of citrus viroids. Mol. Cell. Probe. 2006, 20, 105–113. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Zhang, S.; Cao, L.; Huang, Y.; Cheng, J.; Wu, G.; Tian, S.; Chen, C.; Liu, Y. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nat. Genet. 2017, 49, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Pillitteri, L.J.; Lovatt, C.J.; Walling, L.L. Isolation and characterization of LEAFY and APETALA1 homologues from Citrus sinensis L. Osbeck ’Washington’. J. Am. Soc. Hortic. Sci. 2004, 129, 846–856. [Google Scholar] [CrossRef]
- Whitham, S.A.; Yang, C.; Goodin, M.M. Global impact: elucidating plant responses to viral infection. Mol. Plant Microbe Interact. 2006, 19, 1207–1215. [Google Scholar] [CrossRef]
- Pallas, V.; García, J. How do plant viruses induce disease? Interactions and interference with host components. J. Gen. Virol. 2011, 92, 2691–2705. [Google Scholar] [CrossRef] [Green Version]
- Allie, F.; Pierce, E.J.; Okoniewski, M.J.; Rey, C. Transcriptional analysis of South African cassava mosaic virus-infected susceptible and tolerant landraces of cassava highlights differences in resistance, basal defense and cell wall associated genes during infection. BMC Genomics 2014, 15, 1006. [Google Scholar] [CrossRef]
- Whenham, R.J.; Fraser, R.S.; Brown, L.P.; Payne, J.A. Tobacco-mosaic-virus-induced increase in abscisic-acid concentration in tobacco leaves. Planta 1986, 168, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small rnas. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Gisel, A.; Rodio, M.E.; Delgado, S.; Flores, R.; Serio, F.D. Small RNAs containing the pathogenic determinant of a chloroplast-replicating viroid guide the degradation of a host mRNA as predicted by RNA silencing. Plant J. 2012, 70, 991–1003. [Google Scholar] [CrossRef]
- Wang, M.B.; Masuta, C.; Smith, N.A.; Shimura, H. RNA silencing and plant viral diseases. Mol. Plant Microbe Interact. 2012, 25, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Shan, L.; He, P. Microbial signature-triggered plant defense responses and early signaling mechanisms. Plant Sci. 2014, 228, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.; Ma, W.; Lemtiri-Chlieh, F.; Tsaltas, D.; Leng, Q.; Bodman, S.V.; Berkowitz, G.A. Death don’t have no mercy and neither does calcium: Arabidopsis CYCLIC NUCLEOTIDE GATED CHANNEL2 and innate immunity. Plant Cell 2007, 19, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Prueger, J. Seasonal patterns of carbon dioxide flux over corn canopies. Plos Pathog. 2013, 9, 74–80. [Google Scholar]
- Gómezgómez, L.; Boller, T. FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 2000, 5, 1003–1011. [Google Scholar] [CrossRef]
- Tena, G.; Boudsocq, M.; Sheen, J. Protein kinase signaling networks in plant innate immunity. Curr. Opin. Plant Biol. 2011, 14, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Ye, T.; Hao, L.; Chen, H.; Wang, S.; Fan, Z.; Guo, L.; Zhou, T. Infection of apple by apple stem grooving virus leads to extensive alterations in gene expression patterns but no disease symptoms. PLoS ONE 2014, 9, e95239. [Google Scholar] [CrossRef] [PubMed]
- Lópezgresa, M.P.; Lisón, P.; Yenush, L.; Conejero, V.; Rodrigo, I.; Bellés, J.M. Salicylic acid is involved in the basal resistance of tomato plants to citrus exocortis viroid and tomato spotted wilt virus. PLoS ONE 2016, 11, e0166938. [Google Scholar]
- Alam, S.B.; Rochon, D. Cucumber necrosis virus recruits cellular heat shock protein 70 homologs at several stages of infection. J. Virol. 2015, 90, 3302–3317. [Google Scholar] [CrossRef] [PubMed]
- Hafren, A.; Hofius, D.; Ronnholm, G.; Sonnewald, U.; Makinen, K. HSP70 and its cochaperone CPIP promote potyvirus infection in Nicotiana benthamiana by regulating viral coat protein functions. Plant Cell 2010, 22, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.S.; Dadalto, S.P.; Gonçalves, A.B.; de Souza, G.B.; Barros, V.A.; Fietto, L.G. Transcription factor functional protein-protein interactions in plant defense responses. Proteomes 2014, 2, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Orctová, L.; Steger, G.; Riesner, D. Biolistic inoculation of plants with viroid nucleic acids. J. Virol. Methods 2004, 122, 153–164. [Google Scholar] [CrossRef]
- Hammond, R.W.; Yan, Z. Modification of tobacco plant development by sense and antisense expression of the tomato viroid-induced AGC VIIIa protein kinase PKV suggests involvement in gibberellin signaling. BMC Plant Biol. 2009, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Novak, O.; Napier, R.; Ljung, K. Zooming in on plant hormone analysis: Tissue- and cell-specific approaches. Annu. Rev. Plant Biol. 2017, 68, 323–348. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, F.; Cao, X.; Chen, M.; Ye, G.; Wei, C.; Li, Y. The rice dwarf virus P2 protein interacts with ent-kaurene oxidases in vivo, leading to reduced biosynthesis of gibberellins and rice dwarf symptoms. Plant Physiol. 2005, 139, 1935–1945. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Conti, G.; Zavallo, D.; Manacorda, C.A.; Asurmendi, S. TMV-Cg Coat Protein stabilizes DELLA proteins and in turn negatively modulates salicylic acid-mediated defense pathway during Arabidopsis thaliana viral infection. BMC Plant Biol. 2014, 14, 210. [Google Scholar] [CrossRef]
- Collum, T.D.; Culver, J.N. The impact of phytohormones on virus infection and disease. Curr. Opin. Virol. 2016, 17, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, P.M.; Fusaro, A.F. Plant science. Viruses face a double defense by plant small RNAs. Science 2006, 313, 54–55. [Google Scholar] [CrossRef] [PubMed]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef]
- Garciaruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Parent, J.S.; Bouteiller, N.; Elmayan, T.; Vaucheret, H. Respective contributions of Arabidopsis DCL2 and DCL4 to RNA silencing. Plant J. 2015, 81, 223–232. [Google Scholar] [CrossRef]
- Harvey, J.J.W.; Lewsey, M.G.; Patel, K.; Westwood, J.; Heimstädt, S.; Carr, J.P.; Baulcombe, D.C. An antiviral defense role of AGO2 in plants. PLoS ONE 2011, 6, e14639. [Google Scholar] [CrossRef]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.S.; Perry, K.L.; Moffett, P. ARGONAUTE2 mediates RNA-silencing antiviral defenses against Potato virus X in Arabidopsis. Plant Physiol. 2011, 156, 1556–1564. [Google Scholar] [CrossRef]
- Scholthof, H.B.; Alvarado, V.Y.; Vega-Arreguin, J.C.; Ciomperlik, J.; Odokonyero, D.; Brosseau, C.; Jaubert, M.; Zamora, A.; Moffett, P. Identification of an ARGONAUTE for Antiviral RNA Silencing in Nicotiana benthamiana. Plant Physiol. 2011, 156, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A.; Fahlgren, N.; Garcia-Ruiz, H.; Gilbert, K.B.; Montgomery, T.A.; Nguyen, T.; Cuperus, J.T.; Carrington, J.C. Functional analysis of three Arabidopsis ARGONAUTES using slicer-defective mutants. Plant Cell 2012, 24, 3613–3629. [Google Scholar] [CrossRef]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [Green Version]
- Rakhshandehroo, F.M.; Squires, J.; Palukaitis, P. The influence of RNA-dependent RNA polymerase 1 on potato virus Y infection and on other antiviral response genes. Mol. Plant Microbe Interact. 2009, 22, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Fu, S.F.; Li, Z.; Murphy, A.M.; Dobson, E.A.; Garland, L.; Chaluvadi, S.R.; Lewsey, M.G.; Nelson, R.S.; Carr, J.P. Salicylic acid treatment and expression of an RNA-dependent RNA polymerase 1 transgene inhibit lethal symptoms and meristem invasion during tobacco mosaic virus infection in Nicotiana benthamiana. BMC Plant Biol. 2016, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Schiebel, W.; Pélissier, T.; Riedel, L.; Thalmeir, S.; Schiebel, R.; Kempe, D.; Lottspeich, F.; Sänger, H.L.; Wassenegger, M. Isolation of an RNA-directed RNA polymerase–specific cDNA clone from tomato. Plant Cell 1998, 10, 2087–2101. [Google Scholar] [PubMed]
- Ding, B. The Biology of Viroid-Host Interactions. Annu. Rev. Phytopathol. 2009, 47, 105–131. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science 2002, 296, 1270–1273. [Google Scholar] [CrossRef] [PubMed]
- Diazpendon, J.A.; Li, F.; Li, W.X.; Ding, S.W. Suppression of antiviral silencing by cucumber mosaic virus 2b protein in Arabidopsis is associated with drastically reduced accumulation of three classes of viral small interfering RNAs. Plant Cell 2007, 19, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Cao, M.; Du, P.; Wang, X.; Yu, Y.Q.; Qiu, Y.H.; Li, W.; Gal-On, A.; Zhou, C.; Li, Y.; Ding, S.W. Virus infection triggers widespread silencing of host genes by a distinct class of endogenous siRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14613–14618. [Google Scholar] [CrossRef] [Green Version]
- Dalmay, T.; Horsefield, R.; Braunstein, T.H.; Baulcombe, D.C. SDE3 encodes an RNA helicase required for post-transcriptional gene silencing in Arabidopsis. Embo J. 2014, 20, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Kocábek, T.; Patzak, J.; Bˇríza, J.; Siglová, K.; Mishra, A.K.; Duraisamy, G.S.; Týcová, A.; Ono, E.; Krofta, K. The “putative” role of transcription factors from HlWRKY family in the regulation of the final steps of prenylflavonid and bitter acids biosynthesis in hop (Humulus lupulus L.). Plant Mol. Biol. 2016, 92, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Clean Reads | Clean Bases | Q20 (%) | Q 30(%) | GC Content (%) |
|---|---|---|---|---|---|---|
| HC1 | 86,594,558 | 83,854,342 | 12.45G | 97.66 | 93.99 | 44.75 |
| HC2 | 73,264,886 | 68,562,642 | 10.16G | 97.51 | 94.13 | 48.73 |
| HC3 | 95,947,986 | 89,863,580 | 13.24G | 97.68 | 94.31 | 44.36 |
| CEV1 | 117,995,510 | 113,662,956 | 16.91G | 97.72 | 94.10 | 44.35 |
| CEV2 | 107,136,484 | 104,075,096 | 15.50G | 97.87 | 94.72 | 42.76 |
| CEV3 | 80,867,964 | 75,965,424 | 11.32G | 97.73 | 94.39 | 42.65 |
| GO Accession | Description | Term Type | Corrected p-Value |
|---|---|---|---|
| GO:0006351 | transcription, DNA-templated | BP | 8.13 × 10−5 |
| GO:0097659 | nucleic acid-templated transcription | BP | 8.13 × 10−5 |
| GO:0032774 | RNA biosynthetic process | BP | 8.13 × 10−5 |
| GO:0019219 | regulation of nucleobase-containing compound metabolic process | BP | 8.13 × 10−5 |
| GO:0006355 | regulation of transcription, DNA-templated | BP | 8.13 × 10−5 |
| GO:1903506 | regulation of nucleic acid-templated transcription | BP | 8.13 × 10−5 |
| GO:0051252 | regulation of RNA metabolic process | BP | 8.13 × 10−5 |
| GO:2001141 | regulation of RNA biosynthetic process | BP | 8.13× 10−5 |
| GO:0006030 | chitin metabolic process | BP | 8.13 × 10−5 |
| GO:1901071 | glucosamine-containing compound metabolic process | BP | 8.13 × 10−5 |
| GO:0006032 | chitin catabolic process | BP | 8.13 × 10−5 |
| GO:0046348 | amino sugar catabolic process | BP | 8.13 × 10−5 |
| GO:1901072 | glucosamine-containing compound catabolic process | BP | 8.13 × 10−5 |
| GO:0016998 | cell wall macromolecule catabolic process | BP | 0.000133 |
| GO:0006026 | aminoglycan catabolic process | BP | 0.000133 |
| GO:2000112 | regulation of cellular macromolecule biosynthetic process | BP | 0.001066 |
| GO:0006040 | amino sugar metabolic process | BP | 0.001132 |
| GO:0010556 | regulation of macromolecule biosynthetic process | BP | 0.001132 |
| GO:0031326 | regulation of cellular biosynthetic process | BP | 0.001132 |
| GO:0009889 | regulation of biosynthetic process | BP | 0.001235 |
| GO:0051171 | regulation of nitrogen compound metabolic process | BP | 0.001391 |
| GO:0034654 | nucleobase-containing compound biosynthetic process | BP | 0.002117 |
| GO:0044036 | cell wall macromolecule metabolic process | BP | 0.004343 |
| GO:0010468 | regulation of gene expression | BP | 0.004949 |
| GO:0080090 | regulation of primary metabolic process | BP | 0.006186 |
| GO:0031323 | regulation of cellular metabolic process | BP | 0.00663 |
| GO:0060255 | regulation of macromolecule metabolic process | BP | 0.009714 |
| GO:0018130 | heterocycle biosynthetic process | BP | 0.013439 |
| GO:1901362 | organic cyclic compound biosynthetic process | BP | 0.013439 |
| GO:0006022 | aminoglycan metabolic process | BP | 0.014413 |
| GO:0019438 | aromatic compound biosynthetic process | BP | 0.015638 |
| GO:0019222 | regulation of metabolic process | BP | 0.02637 |
| GO:0043207 | response to external biotic stimulus | BP | 0.033447 |
| GO:0051707 | response to other organism | BP | 0.033447 |
| GO:0006468 | protein phosphorylation | BP | 0.040882 |
| GO:0004568 | chitinase activity | MF | 8.13 × 10−5 |
| GO:0001071 | nucleic acid binding transcription factor activity | MF | 0.000204 |
| GO:0003700 | transcription factor activity, sequence-specific DNA binding | MF | 0.000204 |
| GO:0004674 | protein serine/threonine kinase activity | MF | 0.000761 |
| GO:0004672 | protein kinase activity | MF | 0.014413 |
| GO:0008061 | chitin binding | MF | 0.0321 |
| GO:0020037 | heme binding | MF | 0.033855 |
| GO Accession | Description | Term Type | Corrected p-Value |
|---|---|---|---|
| GO:0009733 | response to auxin | BP | 1.70 × 10−6 |
| GO:0009725 | response to hormone | BP | 1.07 × 10−5 |
| GO:0009719 | response to endogenous stimulus | BP | 1.07 × 10−5 |
| GO:0010033 | response to organic substance | BP | 3.29 × 10−5 |
| GO:0045229 | external encapsulating structure organization | BP | 0.002848 |
| GO:0044264 | cellular polysaccharide metabolic process | BP | 0.004845 |
| GO:0009832 | plant-type cell wall biogenesis | BP | 0.004996 |
| GO:0010215 | cellulose microfibril organization | BP | 0.004996 |
| GO:0030198 | extracellular matrix organization | BP | 0.004996 |
| GO:0043062 | extracellular structure organization | BP | 0.004996 |
| GO:0070726 | cell wall assembly | BP | 0.004996 |
| GO:0071668 | plant-type cell wall assembly | BP | 0.004996 |
| GO:0042221 | response to chemical | BP | 0.007365 |
| GO:0071555 | cell wall organization | BP | 0.007365 |
| GO:0006073 | cellular glucan metabolic process | BP | 0.007365 |
| GO:0044042 | glucan metabolic process | BP | 0.007365 |
| GO:0005976 | polysaccharide metabolic process | BP | 0.008838 |
| GO:0016049 | cell growth | BP | 0.013705 |
| GO:0044262 | cellular carbohydrate metabolic process | BP | 0.025726 |
| GO:0009664 | plant-type cell wall organization | BP | 0.027923 |
| GO:0071669 | plant-type cell wall organization or biogenesis | BP | 0.027923 |
| GO:0042546 | cell wall biogenesis | BP | 0.036742 |
| GO:0071554 | cell wall organization or biogenesis | BP | 0.037842 |
| GO:0040007 | growth | BP | 0.038351 |
| GO:0005618 | cell wall | CC | 0.004996 |
| GO:0031225 | anchored component of membrane | CC | 0.007945 |
| GO:0030312 | external encapsulating structure | CC | 0.018609 |
| GO:0005507 | copper ion binding | MF | 9.43 × 10−5 |
| Term | ID | Input Number | Background Number | Corrected p-Value |
|---|---|---|---|---|
| Up-Regulated Kegg Pathways (Corrected p-Value<0.05) | ||||
| Glutathione metabolism | cit00480 | 18 | 95 | 6.45 × 10−6 |
| Plant-pathogen interaction | cit04626 | 18 | 208 | 0.044203 |
| Biosynthesis of secondary metabolites | cit01110 | 52 | 930 | 0.047811 |
| Amino sugar and nucleotide sugar metabolism | cit00520 | 12 | 120 | 0.047811 |
| Down-Regulated Kegg Pathways (Corrected p-Value < 0.05) | ||||
| Plant hormone signal transduction | cit04075 | 10 | 217 | 0.000251 |
| Phenylpropanoid biosynthesis | cit00940 | 7 | 159 | 0.004052 |
| Phenylalanine metabolism | cit00360 | 5 | 115 | 0.022122 |
| Gene ID | log2 Fold Change | p-adjusted | Function |
|---|---|---|---|
| Cm116590 | 1.5929 | 0.048035 | DRL27: Disease resistance protein At4g27190 |
| Cm029390 | 6.1457 | 2.21 × 10−18 | RBOHD: Respiratory burst oxidase homolog protein D |
| Cm070920 | 1.6338 | 0.020128 | EFTM: Elongation factor Tu, mitochondrial |
| Cm216070 | Inf * | 5.50 × 10−7 | CML8: Calmodulin-like protein 8 |
| Cm056030 | 6.7263 | 3.39 × 10−24 | CML19: Putative calcium-binding protein CML19 |
| Cm113060 | 2.7079 | 4.10 × 10−6 | CML27: Probable calcium-binding protein CML27 |
| Cm128110 | 5.2652 | 0.00012824 | CML31: Probable calcium-binding protein CML31 |
| Cm040570 | 3.4838 | 1.15 × 10−7 | CML44: Probable calcium-binding protein CML44 |
| Cm241600 | 3.0724 | 4.07 × 10−7 | ALLB3: Calcium-binding allergen Bet v 3 |
| Cm050430 | 2.513 | 0.00030948 | ALL8: Calcium-binding allergen Ole e 8 |
| Novel01586 | –2.1223 | 0.036209 | PREDICTED: calcium-dependent protein kinase 9-like |
| Cm106570 | 4.4746 | 2.62 × 10−6 | CNG13: Putative cyclic nucleotide-gated ion channel 13 |
| Cm226290 | 2.8711 | 0.00020247 | WRK33: Probable WRKY transcription factor 33 |
| Cm011420 | 3.1569 | 9.55 × 10−8 | TIF9: Protein TIFY 9 |
| Cm220810 | 3.0776 | 3.14 × 10−6 | TI10A: Protein TIFY 10A |
| Cm157630 | 4.425 | 3.17 × 10−8 | TI10A: Protein TIFY 10A |
| Cm199700 | 3.0945 | 0.0029031 | CERK1: Chitin elicitor receptor kinase 1 |
| Cm178700 | 4.3046 | 0.030672 | RIN4: RPM1-interacting protein 4 |
| Cm178710 | 2.4288 | 0.0032793 | RIN4: RPM1-interacting protein 4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wu, J.; Qiu, Y.; Atta, S.; Zhou, C.; Cao, M. Global Transcriptomic Analysis Reveals Insights into the Response of ‘Etrog’ Citron (Citrus medica L.) to Citrus Exocortis Viroid Infection. Viruses 2019, 11, 453. https://doi.org/10.3390/v11050453
Wang Y, Wu J, Qiu Y, Atta S, Zhou C, Cao M. Global Transcriptomic Analysis Reveals Insights into the Response of ‘Etrog’ Citron (Citrus medica L.) to Citrus Exocortis Viroid Infection. Viruses. 2019; 11(5):453. https://doi.org/10.3390/v11050453
Chicago/Turabian StyleWang, Yafei, Jiaxing Wu, Yuanjian Qiu, Sagheer Atta, Changyong Zhou, and Mengji Cao. 2019. "Global Transcriptomic Analysis Reveals Insights into the Response of ‘Etrog’ Citron (Citrus medica L.) to Citrus Exocortis Viroid Infection" Viruses 11, no. 5: 453. https://doi.org/10.3390/v11050453
APA StyleWang, Y., Wu, J., Qiu, Y., Atta, S., Zhou, C., & Cao, M. (2019). Global Transcriptomic Analysis Reveals Insights into the Response of ‘Etrog’ Citron (Citrus medica L.) to Citrus Exocortis Viroid Infection. Viruses, 11(5), 453. https://doi.org/10.3390/v11050453