Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 through 6 (PPV2–PPV6) in Polish Swine Farms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Samples Processing and DNA Extraction
2.3. Real-Time PCR Detection of Porcine Parvoviruses
2.4. Statistical Analysis
3. Results
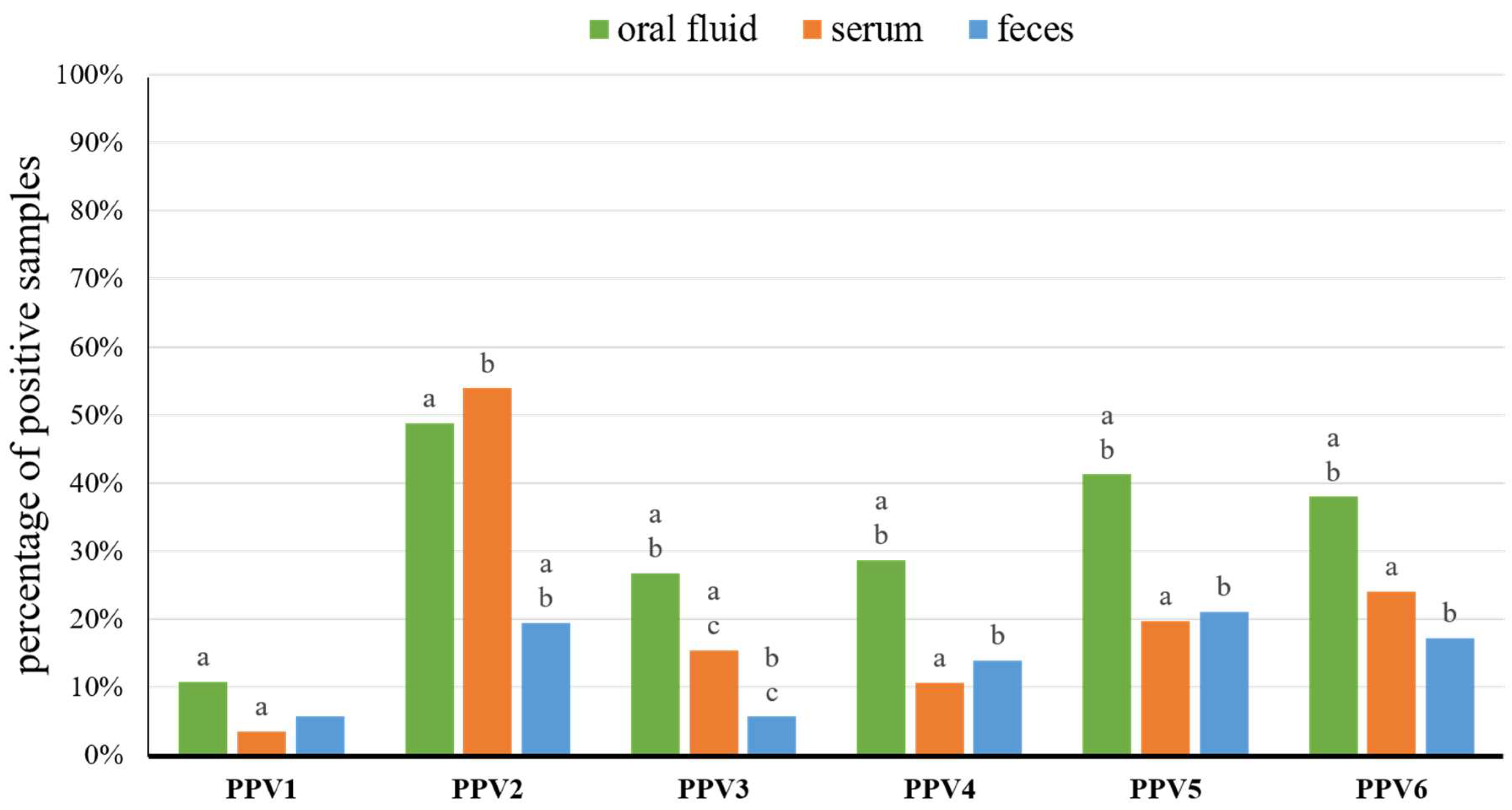
3.1. Detection Rates of Porcine Parvoviruses in Pig Farms
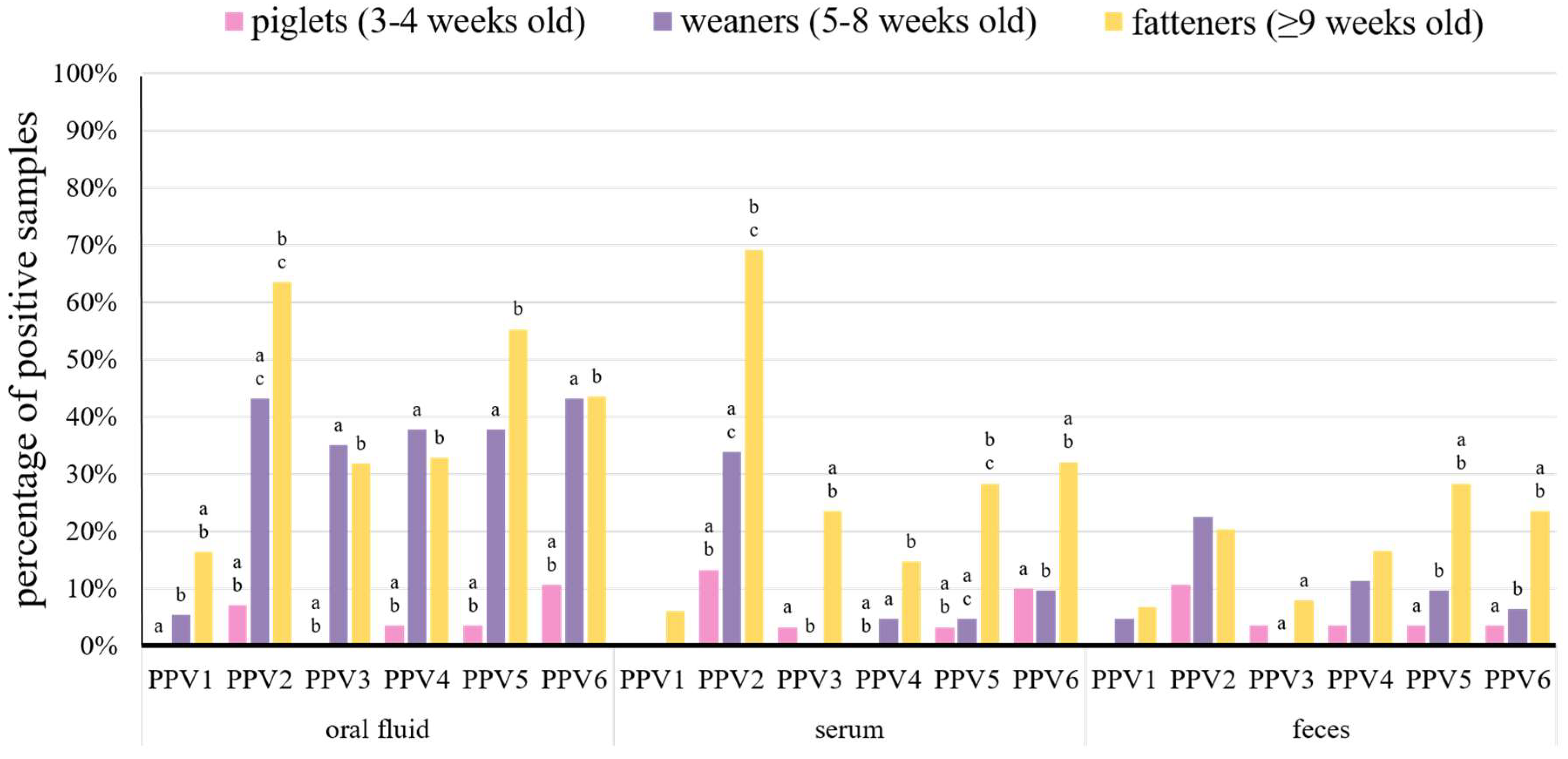
3.2. Detection Rates of Porcine Parvoviruses in Pigs of Different Age
3.3. Probability of PPV1–PPV6 Detection in Different Diagnostic Materials and Age Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tijssen, P.; Agbandje-McKenna, M.; Almendral, J.M.; Bergoin, M.; Flegel, T.W.; Hedman, K.; Kleinschmidt, J.; Li, Y.; Pintel, D.J.; Tattersall, P. Family Parvoviridae. In Virus Taxonomy. 9th report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Academic Press: London, UK, 2012; pp. 405–425. [Google Scholar]
- Mengeling, W.L.; Lager, K.M.; Zimmerman, J.K.; Samarikermani, N.; Beran, G.W. A current assessment of the role of porcine parvovirus as a cause of fetal porcine death. J. Vet. Diagn. Investig. 1991, 3, 33–35. [Google Scholar] [CrossRef]
- Mengeling, W.; Lager, K.; Vorwald, A. The effect of porcine parvovirus and porcine reproductive and respiratory syndrome virus on porcine reproductive performance. Anim. Reprod. Sci. 2000, 60, 199–210. [Google Scholar] [CrossRef]
- Allan, G.M.; Kennedy, S.; McNeilly, F.; Foster, J.C.; Ellis, J.A.; Krakowka, S.J.; Meehan, B.M.; Adair, B.M. Experimental reproduction of severe wasting disease by co-infection of pigs with porcine circovirus and porcine parvovirus. J. Comp. Pathol. 1999, 121, 1–11. [Google Scholar] [CrossRef]
- Hijikata, M.; Abe, K.; Win, K.M.; Shimizu, Y.K.; Keicho, N.; Yoshikura, H. Identification of new parvovirus DNA sequence in swine sera from Myanmar. Jpn. J. Infect. Dis. 2001, 54, 244–245. [Google Scholar]
- Wang, F.; Wei, Y.; Zhu, C.; Huang, X.; Xu, Y.; Yu, L.; Yu, X. Novel parvovirus sublineage in the family of Parvoviridae. Virus Genes 2010, 41, 305–308. [Google Scholar] [CrossRef]
- Lau, S.K.; Woo, P.C.; Tse, H.; Fu, C.T.; Au, W.K.; Chen, X.C.; Tsoi, H.W.; Tsang, T.H.; Chan, J.S.; Tsang, D.N.; et al. Identification of novel porcine and bovine parvoviruses closely related to human parvovirus 4. J. Gen. Virol. 2008, 89, 1840–1848. [Google Scholar] [CrossRef]
- Cheung, A.K.; Wu, G.; Wang, D.; Bayles, D.O.; Lager, K.M.; Vincent, A.L. Identification and molecular cloning of a novel porcine parvovirus. Arch. Virol. 2010, 155, 801–806. [Google Scholar] [CrossRef]
- Xiao, C.T.; Giménez-Lirola, L.G.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Characterization of a novel porcine parvovirus tentatively designated PPV5. PLoS ONE 2013, 8, e65312. [Google Scholar] [CrossRef]
- Ni, J.; Qiao, C.; Han, X.; Han, T.; Kang, W.; Zi, Z.; Cao, Z.; Zhai, X.; Cai, X. Identification and genomic characterization of a novel porcine parvovirus (PPV6) in China. Virol. J. 2014, 11, 203. [Google Scholar] [CrossRef]
- Palinski, R.M.; Mitra, N.; Hause, B.M. Discovery of a novel Parvovirinae virus, porcine parvovirus 7, by metagenomic sequencing of porcine rectal swabs. Virus Genes 2016, 52, 564–567. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef]
- Sun, J.; Huang, L.; Wei, Y.; Wang, Y.; Chen, D.; Du, W.; Wu, H.; Liu, C. Prevalence of emerging porcine parvoviruses and their co-infections with porcine circovirus type 2 in China. Arch. Virol. 2015, 160, 1339–1344. [Google Scholar] [CrossRef]
- Wu, R.; Wen, Y.; Huang, X.; Wen, X.; Yan, Q.; Huang, Y.; Ma, X.; Cao, S. First complete genomic characterization of a porcine parvovirus 5 isolate from China. Arch. Virol. 2014, 159, 1533–1536. [Google Scholar] [CrossRef]
- Xing, X.; Zhou, H.; Tong, L.; Chen, Y.; Sun, Y.; Wang, H.; Zhang, G. First identification of porcine parvovirus 7 in China. Arch. Virol. 2017, 163, 209–213. [Google Scholar] [CrossRef]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G. Identification of recently described porcine parvoviruses in archived North American samples from 1996 and association with porcine circovirus associated disease. Vet. Microbiol. 2014, 173, 9–16. [Google Scholar] [CrossRef]
- Schirtzinger, E.E.; Suddith, A.W.; Hause, B.M.; Hesse, R.A. First identification of porcine parvovirus 6 in North America by viral metagenomic sequencing of serum from pigs infected with porcine reproductive and respiratory syndrome virus. Virol. J. 2015, 12, 170. [Google Scholar] [CrossRef]
- Cadar, D.; Lőrincz, M.; Kiss, T.; Novosel, D.; Podgórska, K.; Becskei, Z.; Tuboly, T.; Cságola, A. Emerging novel porcine parvoviruses in Europe: Origin, evolution, phylodynamics and phylogeography. J. Gen. Virol. 2013, 94, 2330–2337. [Google Scholar] [CrossRef]
- Cui, J.; Biernacka, K.; Fan, J.; Gerber, P.F.; Stadejek, T.; Opriessnig, T. Circulation of Porcine Parvovirus Types 1 through 6 in Serum Samples Obtained from Six Commercial Polish Pig Farms. Transbound Emerg. Dis. 2017, 64, 1945–1952. [Google Scholar] [CrossRef]
- Miłek, D.; Woźniak, A.; Stadejek, T. The detection and genetic diversity of novel porcine parvovirus 7 (PPV7) on Polish pig farms. Res. Vet. Sci. 2018, 120, 28–32. [Google Scholar] [CrossRef]
- Cságola, A.; Lorincz, M.; Cadar, D.; Tombácz, K.; Biksi, I.; Tuboly, T. Detection, prevalence and analysis of emerging porcine parvovirus infections. Arch. Virol. 2012, 157, 1003–1010. [Google Scholar] [CrossRef]
- Saekhow, P.; Ikeda, H. Prevalence and genomic characterization of porcine parvoviruses detected in Chiangmai area of Thailand in 2011. Microbiol. Immunol. 2015, 59, 82–88. [Google Scholar] [CrossRef]
- Saekhow, P.; Kishizuka, S.; Sano, N.; Mitsui, H.; Akasaki, H.; Mawatari, T.; Ikeda, H. Coincidental detection of genomes of porcine parvoviruses and porcine circovirus type 2 infecting pigs in Japan. J. Vet. Med. Sci. 2016, 77, 1581–1586. [Google Scholar] [CrossRef]
- Afolabi, K.O.; Iweriebor, B.C.; Obi, L.C.; Okoh, A.I. Prevalence of porcine parvoviruses in some South African swine herds with background of porcine circovirus type 2 infection. Acta Trop. 2018, 190, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Szelei, J.; Liu, K.; Li, Y.; Fernandes, S.; Tijssen, P. Parvovirus 4-like virus in blood products. Emerg. Infect. Dis. 2010, 16, 561–564. [Google Scholar] [CrossRef]
- Gava, D.; Souza, C.K.; Schaefer, R.; Vincent, A.L.; Cantão, M.E.; Coldebella, A.; Ciacci-Zanella, J.R. A TaqMan-based real-time PCR for detection ad quantification of porcine parvovirus 4. J. Virol. Methods 2015, 19, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Ouh, I.O.; Park, S.; Lee, J.Y.; Song, J.Y.; Cho, I.S.; Kim, H.R.; Park, C.K. First detection and genetic characterization of porcine parvovirus 7 from Korean domestic pig farms. J. Vet. Sci. 2018, 19, 855–857. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G. Emergence of a novel mutant PCV2b variant associated with clinical PCVAD in two vaccinated pig farms in the U.S. concurrently infected with PPV2. Vet. Microbiol. 2013, 163, 177–183. [Google Scholar] [CrossRef]
- Novosel, D.; Cadar, D.; Tuboly, T.; Jungic, A.; Stadejek, T.; Ait-Ali, T.; Cságola, A. Investigating porcine parvoviruses genogroup 2 infection using in situ polymerase chain reaction. BMC Vet. Res. 2018, 14, 163. [Google Scholar] [CrossRef]
- Huang, L.; Zhai, S.L.; Cheung, A.K.; Zhang, H.B.; Long, J.X.; Yuan, S.S. Detection of a novel porcine parvovirus, PPV4, in Chinese swine herds. Virol. J. 2010, 7, 333. [Google Scholar] [CrossRef]
- Cibulski, S.P.; Teixeira, T.F.; Varela, A.P.M.; Scheffer, C.M.; Santos, H.F.; Lima, F.E.S.; Roehe, P.M. Ungulate copiparvovirus 2 in healthy and postweaning multisystemic wasting syndrome-affected pigs. Trop. Anim. Health Prod. 2017, 49, 945–949. [Google Scholar] [CrossRef]
- Xiao, C.T.; Gerber, P.F.; Giménez-Lirola, L.G.; Halbur, P.G.; Opriessnig, T. Increasing porcine PARV4 prevalence with pig age in the U.S. pig population. Vet. Microbiol. 2012, 160, 290–296. [Google Scholar] [CrossRef]
- Xiao, C.T.; Gerber, P.F.; Giménez-Lirola, L.G.; Halbur, P.G.; Opriessnig, T. Characterization of porcine parvovirus type 2 (PPV2) which is highly prevalent in the USA. Vet. Microbiol. 2013, 161, 325–330. [Google Scholar] [CrossRef]
- Cságola, A.; Zádori, Z.; Mészáros, I.; Tuboly, T. Detection of Porcine Parvovirus 2 (Ungulate Tetraparvovirus 3) Specific Antibodies and Examination of the Serological Profile of an Infected Swine Herd. PLoS ONE 2016, 11, e0151036. [Google Scholar] [CrossRef]
- Opriessnig, T.; Shen, H.G.; Pal, N.; Ramamoorthy, S.; Huang, Y.W.; Lager, K.M.; Beach, N.M.; Halbur, P.G.; Meng, X.J. A live-attenuated chimeric porcine circovirus type 2 (PCV2) vaccine is transmitted to contact pigs but is not upregulated by concurrent infection with porcine parvovirus (PPV) and porcine reproductive and respiratory syndrome virus (PRRSV) and is efficacious in a PCV2b-PRRSV-PPV challenge model. Clin. Vaccine Immunol. 2011, 18, 1261–1268. [Google Scholar] [CrossRef]
- Cui, J.; Fan, J.; Gerber, P.F.; Biernacka, K.; Stadejek, T.; Xiao, C.T.; Opriessnig, T. First identification of porcine parvovirus 6 in Poland. Virus Genes 2017, 53, 100–104. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www/R-project.org/ (accessed on 17 May 2019).
- Harrell, F.E. rms: Regression Modeling Strategies. R package version 5.1-3. 2019. Available online: https://CRAN.R-project.org/package=rms (accessed on 17 May 2019).
- Prickett, J.R.; Zimmerman, J.J. The development of oral fluid-based diagnostics and applications in veterinary medicine. Anim. Health Res. Rev. 2010, 11, 207–216. [Google Scholar] [CrossRef]
- Mészáros, I.; Olasz, F.; Cságola, A.; Tijssen, P.; Zádori, Z. Biology of Porcine Parvovirus (Ungulate parvovirus 1). Viruses 2017, 9, 393. [Google Scholar] [CrossRef]
- Joo, H.; Donaldson-Wood, C.; Johnson, R. Observations on the pathogenesis of porcine parvovirus infection. Arch. Virol. 1976, 51, 123–129. [Google Scholar] [CrossRef]
- Gava, D.; Souza, C.K.; Mores, T.J.; Argenti, L.E.; Streck, A.F.; Canal, C.W.; Bortolozzo, F.P.; Wentz, I. Dynamics of vanishing of maternally derived antibodies of Ungulate protoparvovirus 1 suggests an optimal age for gilts vaccination. Trop. Anim. Health Prod. 2017, 49, 1085–1088. [Google Scholar] [CrossRef]
- Etoh, M.; Morishita, E.; Ochiai, M.; Watanabe, Y. Transitional antibodies and spontaneous infection in the Transitional antibodies and spontaneous infection in the porcine (Sus scrofa) parvo viral infections. Jpn. J. Swine Husb. Res. (Japan) 1979, 16, 237–239. [Google Scholar]
- Foerster, T.; Streck, A.F.; Speck, S.; Selbitz, H.-J.; Lindner, T.; Truyen, U. An inactivated whole-virus porcine parvovirus vaccine protects pigs against disease but does not prevent virus shedding even after homologous virus challenge. J. Gen. Virol. 2016, 97, 1408–1413. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Farm ID | PPV1 | PPV2 | PPV3 | PPV4 | PPV5 | PPV6 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | w | fa | p | w | fa | p | w | fa | p | w | fa | p | w | fa | p | w | fa | |
| AK | ▲ | □ | ● | ♦ | ||||||||||||||
| GO | ▲ | ▲ | ■ | ■ | ● | ○ | ○ | ♦ | ||||||||||
| WA | ▲ | ▲ | ■ | ■ | □ | □ | ● | ● | ○ | ♦ | ♦ | |||||||
| PB | ▲ | ■ | ■ | □ | □ | □ | ● | ● | ○ | ○ | ○ | ♦ | ♦ | ♦ | ||||
| PA | ■ | ■ | □ | ● | ○ | ♦ | ||||||||||||
| BA | ■ | ■ | □ | □ | ● | ● | ○ | ○ | ♦ | ♦ | ||||||||
| BO | ■ | ■ | ● | ○ | ○ | |||||||||||||
| KS | ■ | □ | □ | ○ | ♦ | ♦ | ||||||||||||
| KO | ■ | □ | □ | ● | ● | ○ | ○ | ♦ | ♦ | |||||||||
| KU | ■ | □ | □ | ● | ● | ● | ○ | ○ | ♦ | ♦ | ♦ | |||||||
| A | ■ | ● | ○ | ○ | ♦ | |||||||||||||
| B | ■ | ■ | ■ | ○ | ♦ | |||||||||||||
| C | ■ | ■ | □ | ● | ○ | |||||||||||||
| PR | ■ | ■ | ● | ● | ||||||||||||||
| RO | ▲ | ■ | □ | □ | ● | ● | ○ | ♦ | ||||||||||
| SU | ■ | ■ | ■ | ○ | ♦ | |||||||||||||
| WT | ■ | ■ | ■ | □ | □ | ● | ○ | ○ | ♦ | ♦ | ♦ | |||||||
| ZD | ▲ | ■ | ■ | □ | ● | ○ | ♦ | ♦ | ||||||||||
| GK | ▲ | ■ | ■ | □ | ○ | ♦ | ♦ | |||||||||||
| No. of positive farms in each age group | 0 | 3 | 6 | 3 | 13 | 18 | 2 | 8 | 12 | 1 | 10 | 12 | 1 | 9 | 16 | 4 | 11 | 13 |
| Total no. of positive farms | 7 | 18 | 13 | 15 | 17 | 16 | ||||||||||||
 Samples not collected.
Samples not collected.| Age Group (weeks) | PPV1 | PPV2 | PPV3 | PPV4 | PPV5 | PPV6 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| of | s | fe | of | s | fe | of | s | fe | of | s | fe | of | s | fe | of | s | fe | ||
| This study | piglets (3–4) | − | − | − | 7.1% | 13.3% | 10.7% | − | 3.3% | 3.6% | 3.6% | − | 3.6% | 3.6% | 3.3% | 3.6% | 10.7% | 10.0% | 3.6% |
| weaners (5–8) | 5.4% | − | 4.8% | 43.2% | 33.9% | 22.6% | 35.1% | − | − | 37.8% | 4.8% | 11.3% | 37.8% | 4.8% | 9.7% | 43.2% | 9.7% | 6.5% | |
| fatteners (≥9) | 16.5% | 6.2% | 6.8% | 63.5% | 69.1% | 20.4% | 31.8% | 23.5% | 8.0% | 32.9% | 14.8% | 16.7% | 55.3% | 28.4% | 28.4% | 43.5% | 32.1% | 23.5% | |
| TOTAL | 10.7% | 3.5% | 5.6% | 48.7% | 53.9% | 19.4% | 26.7% | 15.4% | 5.6% | 28.7% | 10.6% | 13.9% | 41.3% | 19.7% | 21.0% | 38.0% | 24.0% | 17.1% | |
| Earlier studies [references] | piglets (3–4) | 3.0% [9] | |||||||||||||||||
| weaners (5–8) | 1.6% [19] | 5.1% [33] | 1.8% [9] | 1.6% [19] | |||||||||||||||
| fatteners (≥9) | 26.6% [19] | 10.7% [33] | 11.0% [19] | 3.5% [19] | 3.6% [9] | 5.2% [19] | 5.4% [9] | 8.8–15.5% [10,19] | |||||||||||
| TOTAL | 1.1–7.2% [16,21] | 5.4–19.0% [19,21] 35.2–55.0% [16,28,34] | 6.0–7.6% [21,33] | 9.4–14.4% [16,21] | 10.0% [7] | 2.4–3.3% [16,19,21,28] | 1.9% [9] 7.0–18.2% [21,26] | 3.4–4.0% [16,19] | 2.6% [9] | 6.1–15.2% [10,17,19] | |||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miłek, D.; Woźniak, A.; Guzowska, M.; Stadejek, T. Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 through 6 (PPV2–PPV6) in Polish Swine Farms. Viruses 2019, 11, 474. https://doi.org/10.3390/v11050474
Miłek D, Woźniak A, Guzowska M, Stadejek T. Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 through 6 (PPV2–PPV6) in Polish Swine Farms. Viruses. 2019; 11(5):474. https://doi.org/10.3390/v11050474
Chicago/Turabian StyleMiłek, Dagmara, Aleksandra Woźniak, Magdalena Guzowska, and Tomasz Stadejek. 2019. "Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 through 6 (PPV2–PPV6) in Polish Swine Farms" Viruses 11, no. 5: 474. https://doi.org/10.3390/v11050474
APA StyleMiłek, D., Woźniak, A., Guzowska, M., & Stadejek, T. (2019). Detection Patterns of Porcine Parvovirus (PPV) and Novel Porcine Parvoviruses 2 through 6 (PPV2–PPV6) in Polish Swine Farms. Viruses, 11(5), 474. https://doi.org/10.3390/v11050474