Animals as Reservoir for Human Norovirus



Abstract
:1. Introduction
2. Results
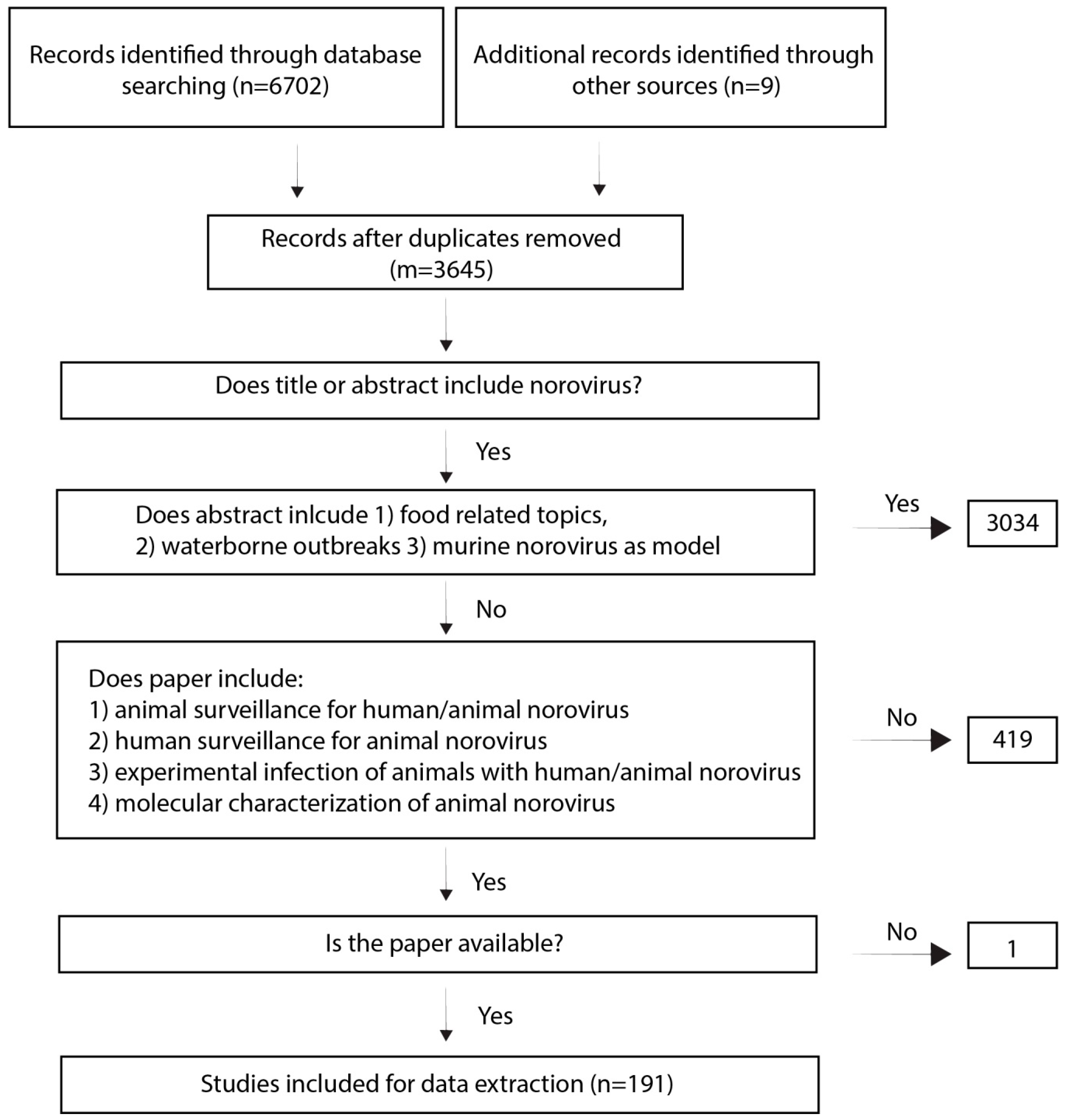
2.1. Search Output:
2.2. Noroviruses in Domesticated and Wild Animals
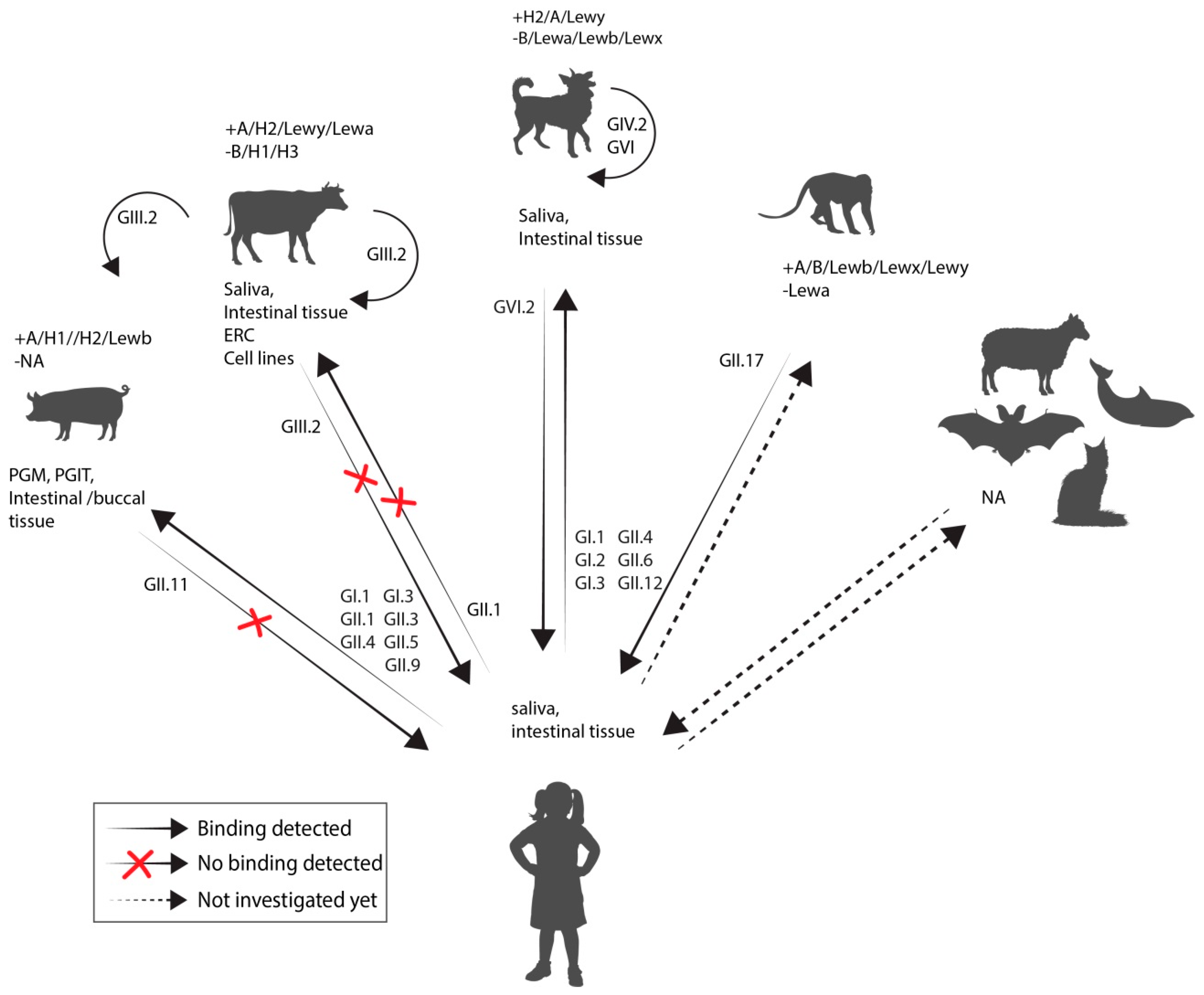
2.3. Is There Evidence for Cross Species Transmission?
2.3.1. Animal-to-Human Transmission
2.3.2. Human-to-Animal Transmission
2.3.3. Susceptibility of Animals to Human Norovirus Strains
3. Discussion and Conclusions
4. Methods
4.1. Search Strategy
4.1.1. embase.com (2903)
4.1.2. Medline Ovid (1550)
4.1.3. Web of Science (2049)
4.1.4. Google Scholar (200)
4.2. Selection Criteria
4.3. Data Extraction
- General description.Location (country, district, city), duration of study, date of study, species and number of tested animals and age of animals.For studies describing experimental infections of animals with human or animal noroviruses, the following information was collected if described in the paper:
- Details on experimental infection methods.Regarding the experimental infection, the route of inoculation was documented since this may affect which subclasses of immunoglobulins are induced. In addition, genogroup/genotype of the virus inoculate, as well as amount used (number of genome copies) and the sample type collected (e.g., saliva, feces, sera) were registered. It was further recorded how virus replication was confirmed, which methods was used to detect virus (RT-PCR, real-time RT-PCR, antigen capturing ELISA, EM), how much was detected and at what time points.
- Details on clinical picture; description of the health state of the animals; which symptoms (e.g., diarrhea, vomiting), as well as the duration of symptoms.
- Pathology; pathological examination results.
- Immunohistochemistry data was extracted to for information regarding the organ and cell tropism.
- Host response was assessed by collecting serological data including method of antibody detection, type of immunoglobulins (Igs) tested (IgM, IgG, IgA), origin of Igs (saliva, sera, feces), the time period Igs were detected and if available whether they were blocking virus from binding to HBGAs. Since some animal noroviruses cluster close to human norovirus, information about cross-reactivity was also collected. Host factors such as HBGA, secretor and non-secretor status were of interest, since they are known to be important for susceptibility in humans, while in animals this link is less evident.
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Vinje, J. Advances in Laboratory Methods for Detection and Typing of Norovirus. J. Clin. Microbiol. 2015, 53, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Throne, L.; Goodfellow, I. Norovirus gene expression and replication. J. Gen. Virol. 2014, 95, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Tanaka, M.M.; White, P.A. Norovirus recombination. J. Gen. Virol. 2007, 88, 3347–3359. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Hansman, G.S.; Clancy, L.E.; Tanaka, M.M.; Rawlinson, W.D.; White, P.A. Norovirus Recombination in ORF1/ORF2 Overlap. Emerg. Infect. Dis. 2005, 11, 1079–1085. [Google Scholar] [CrossRef]
- Kroneman, A.; Vega, E.; Vennema, H.; Vinjé, J.; White, P.A.; Hansman, G.; Green, K.; Martella, V.; Katayama, K.; Koopmans, M. Proposal for a unified norovirus nomenclature and genotyping. Arch. Virol. 2013, 158, 2059–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Zhu, C.; Wang, Y.; Ai, L.; Yang, L.; Ye, F.; Ding, C.; Chen, J.; He, B.; Zhu, J.; et al. Virome analysis for identification of novel mammalian viruses in bats from Southeast China. Sci. Rep. 2017, 7, 10917. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.D.; Chen, X.; Tian, J.H.; Chen, L.J.; Li, K.; Wang, W.; Eden, J.S.; Shen, J.J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Siebenga, J.J.; Vennema, H.; Zheng, D.P.; Vinjé, J.; Lee, B.E.; Pang, X.L.; Ho, E.C.M.; Lim, W.; Choudekar, A.; Broor, S.; et al. Norovirus illness is a global problem: Emergence and spread of norovirus GII. 4 variants, 2001–2007. J. Infect. Dis. 2009, 200, 802–812. [Google Scholar] [CrossRef]
- Van Der Poel, W.H.M.; Vinjé, J.; Van Der Heide, R.; Herrera, M.I.; Vivo, A.; Koopmans, M.P.G. Norwalk-like calicivirus genes in farm animals. Emerg. Infect. Dis. 2000, 6, 36–41. [Google Scholar] [CrossRef]
- Sugieda, M.; Nagaoka, H.; Kakishima, Y.; Ohshita, T.; Nakamura, S.; Nakajima, S. Detection of Norwalk-like virus genes in the caecum contents of pigs. Brief report. Arch. Virol. 1998, 143, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, A.M.; Green, J.; Gallimore, C.I.; Brown, D.W.G.; Bridger, J.C. The bovine Newbury agent-2 is genetically more closely related to human SRSVs than to animal caliciviruses. Virology 1999, 254, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.L.; Lambden, P.R.; Günther, H.; Otto, P.; Elschner, M.; Clarke, I.N. Molecular characterization of a bovine enteric calicivirus: Relationship to the Norwalk-like viruses. J. Virol. 1999, 73, 819–825. [Google Scholar]
- Menon, V.K.; George, S.; Shanti, A.A.; Saravanabavan, A.; Samuel, P.; Ramani, S.; Estes, M.K.; Kang, G. Exposure to human and bovine noroviruses in a birth Cohort in southern India from 2002 to 2006. J. Clin. Microbiol. 2013, 51, 2391–2395. [Google Scholar] [CrossRef]
- Vildevall, M.; Grahn, A.; Oliver, S.L.; Bridger, J.C.; Charpilienne, A.; Poncet, D.; Larson, G.; Svensson, L. Human antibody responses to bovine (newbury-2) norovirus (GIII.2) and association to histo-blood group antigens. J. Med. Virol. 2010, 82, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, J.R.; Costantini, V.P.; Cannon, J.L.; Lin, S.C.; Nascimento, M.S.J.; Vinjé, J. Presence of Antibodies against Genogroup VI Norovirus in Humans. Virol. J. 2013. [Google Scholar] [CrossRef]
- Martino, B.D.; Di Profio, F.; Ceci, C.; Felice, E.D.; Green, K.Y.; Bok, K.; Grazia, S.D.; Giammanco, G.M.; Massirio, I.; Lorusso, E.; et al. Seroprevalence of norovirus genogroup IV antibodies among humans, Italy, 2010–2011. Emerg. Infect. Dis. 2014, 20, 1828–1832. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.W.; Skilling, D.E.; Cherry, N.; Mead, J.H.; Matson, D.O. Calicivirus emergence from ocean reservoirs: Zoonotic and interspecies movements. Emerg. Infect. Dis. 1998, 4, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M.; Wobus, C.E. Viruses in Rodent Colonies: Lessons Learned from Murine Noroviruses. Annu. Rev. Virol. 2015, 2, 525–548. [Google Scholar] [CrossRef]
- Karst, S.M.; Wobus, C.E.; Goodfellow, I.G.; Green, K.Y.; Virgin, H.W. Advances in norovirus biology. Cell Host Microbe 2014, 15, 668–680. [Google Scholar] [CrossRef]
- Baldridge, M.T.; Turula, H.; Wobus, C.E. Norovirus Regulation by Host and Microbe. Trends Mol. Med. 2016, 22, 1047–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Deng, Z. Detection and forecasting of oyster norovirus outbreaks: Recent advances and future perspectives. Mar. Environ. Res. 2012, 80, 62–69. [Google Scholar] [CrossRef]
- Hassard, F.; Harris, J.P.; Jones, D.L.; Sharp, J.H.; Taft, H.; LeVay, L.; McDonald, J.E.; Tuson, K.; Malham, S.K. Critical Review on the Public Health Impact of Norovirus Contamination in Shellfish and the Environment: A UK Perspective. Food Environ. Virol. 2017, 9, 123–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, J.L.; Zickl, R. Winter Vomiting Disease. J. Infect. Dis. 1969, 119, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Kapikian, A.Z. The Discovery of the 27-nm Norwalk Virus: An Historic Perspective. J. Infect. Dis. 2000, 181 (Suppl. 2), S295–S302. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.H.; Myung, G.H.; Cheetham, S.; Souza, M.; Funk, J.A.; Saif, L.J. Porcine noroviruses related to human noroviruses. Emerg. Infect. Dis. 2005, 11, 1874–1881. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, W.; Yang, S.; Cui, L.; Hua, X. Complete genome sequence of a new-genotype porcine norovirus isolated from piglets with diarrhea. J. Virol. 2012, 86, 7015–7016. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhang, W.; Yang, S.; Yang, Z.; Chen, Y.; Cui, L.; Zhu, J.; Hua, X. Recombinant porcine norovirus identified from piglet with diarrhea. BMC Vet. Res. 2012, 8, 155. [Google Scholar] [CrossRef]
- Scheuer, K.A.; Oka, T.; Hoet, A.E.; Gebreyes, W.A.; Molla, B.Z.; Saif, L.J.; Wang, Q. Prevalence of porcine Noroviruses, molecular characterization of emerging porcine sapoviruses from finisher swine in the United States, and unified classification scheme for sapoviruses. J. Clin. Microbiol. 2013, 51, 2344–2353. [Google Scholar] [CrossRef]
- Niendorf, S.; Klemm, U.; Marques, A.M.; Bock, C.T.; Höhne, M. Infection with the persistent murine norovirus strain MNV-S99 suppresses IFN-Beta release and activation of stat1 In vitro. PLoS ONE 2016, 11, e0156898. [Google Scholar] [CrossRef]
- Mattison, K.; Shukla, A.; Cook, A.; Pollari, F.; Friendship, R.; Kelton, D.; Bidawid, S.; Farber, J.M. Human noroviruses in swine and cattle. Emerg. Infect. Dis. 2007, 13, 1184–1188. [Google Scholar] [CrossRef]
- L’Homme, Y.; Sansregret, R.; Plante-Fortier, É.; Lamontagne, A.M.; Lacroix, G.; Ouardani, M.; Deschamps, J.; Simard, G.; Simard, C. Genetic diversity of porcine Norovirus and Sapovirus: Canada, 2005–2007. Arch. Virol. 2009, 154, 581–593. [Google Scholar] [CrossRef]
- L’Homme, Y.; Sansregret, R.; Simard, C. Broad range RT-PCR assays targeting human noroviruses also detect swine noroviruses. Food Microbiol. 2009, 26, 552–555. [Google Scholar] [CrossRef]
- Shen, Q.; Ren, R.; Zhang, W.; Yang, Z.; Yang, S.; Chen, Y.; Cui, L.; Hua, X. Prevalence of hepatitis E virus and porcine caliciviruses in pig farms of Guizhou province, China. Hepat. Mon. 2011, 11, 459–463. [Google Scholar]
- Chao, D.Y.; Wei, J.Y.; Chang, W.F.; Wang, J.; Wang, L.C. Detection of Multiple Genotypes of Calicivirus Infection in Asymptomatic Swine in Taiwan. Zoonoses Public Health 2012, 59, 434–444. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, W.; Yang, S.; Chen, Y.; Ning, H.; Shan, T.; Liu, J.; Yang, Z.; Cui, L.; Zhu, J.; et al. Molecular detection and prevalence of porcine caliciviruses in eastern China from 2008 to 2009. Arch. Virol. 2009, 154, 1625–1630. [Google Scholar] [CrossRef]
- Nakamura, K.; Saga, Y.; Iwai, M.; Obara, M.; Horimoto, E.; Hasegawa, S.; Kurata, T.; Okumura, H.; Nagoshi, M.; Takizawa, T. Frequent detection of noroviruses and sapoviruses in swine and high genetic diversity of porcine sapovirus in Japan during fiscal year 2008. J. Clin. Microbiol. 2010, 48, 1215–1222. [Google Scholar] [CrossRef]
- Song, Y.J.; Yu, J.N.; Nam, H.M.; Bak, H.R.; Lee, J.B.; Park, S.Y.; Song, C.S.; Seo, K.H.; Choi, I.S. Identification of genetic diversity of porcine Norovirus and Sapovirus in Korea. Virus Genes 2011, 42, 394–401. [Google Scholar] [CrossRef]
- Wolf, S.; Williamson, W.; Hewitt, J.; Lin, S.; Rivera-Aban, M.; Ball, A.; Scholes, P.; Savill, M.; Greening, G.E. Molecular detection of norovirus in sheep and pigs in New Zealand farms. Vet. Microbiol. 2009, 133, 184–189. [Google Scholar] [CrossRef]
- Martínez, M.A.; Alcalá, A.C.; Carruyo, G.; Botero, L.; Liprandi, F.; Ludert, J.E. Molecular detection of porcine enteric caliciviruses in Venezuelan farms. Vet. Microbiol. 2006, 116, 77–84. [Google Scholar] [CrossRef]
- Collins, P.J.; Martella, V.; Buonavoglia, C.; O’Shea, H. Detection and characterization of porcine sapoviruses from asymptomatic animals in Irish farms. Vet. Microbiol. 2009, 139, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Machnowska, P.; Ellerbroek, L.; Johne, R. Detection and characterization of potentially zoonotic viruses in faeces of pigs at slaughter in Germany. Vet. Microbiol. 2014, 168, 60–68. [Google Scholar] [CrossRef]
- Mijovski, J.Z.; Poljšak-Prijatelj, M.; Steyer, A.; Barlič-Maganja, D.; Koren, S. Detection and molecular characterisation of noroviruses and sapoviruses in asymptomatic swine and cattle in Slovenian farms. Infect. Genet. Evol. 2010, 10, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Sisay, Z.; Djikeng, A.; Berhe, N.; Belay, G.; Abegaz, W.E.; Wang, Q.H.; Saif, L.J. First detection and molecular characterization of sapoviruses and noroviruses with zoonotic potential in swine in Ethiopia. Arch. Virol. 2016, 161, 2739–2747. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.F.; Alfieri, A.F.; Barry, A.F.; de Arruda Leme, R.; Gardinali, N.R.; van der Poel, W.H.; Alfieri, A.A. High frequency of porcine norovirus infection in finisher units of Brazilian pig-production systems. Trop. Anim. Health Prod. 2015, 47, 237–241. [Google Scholar] [CrossRef]
- Yin, Y.; Tohya, Y.; Ogawa, Y.; Numazawa, D.; Kato, K.; Akashi, H. Genetic analysis of calicivirus genomes detected in intestinal contents of piglets in Japan. Arch. Virol. 2006, 151, 1749–1759. [Google Scholar] [CrossRef]
- Farkas, T.; Nakajima, S.; Sugieda, M.; Deng, X.; Zhong, W.; Jiang, X. Seroprevalence of noroviruses in swine. J. Clin. Microbiol. 2005, 43, 657–661. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Delgado, I.; Costantini, V.; Heenemann, K.; Vahlenkamp, T.W.; Vinjé, J.; Nascimento, M.S.J. Seroprevalence of canine norovirus in 14 European countries. Clin. Vaccine Immunol. 2014, 21, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Sisay, Z.; Wang, Q.; Oka, T.; Saif, L. Prevalence and molecular characterization of porcine enteric caliciviruses and first detection of porcine kobuviruses in US swine. Arch. Virol. 2013, 158, 1583–1588. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Jung, K.; Han, M.G.; Hoet, A.; Scheuer, K.; Wang, Q.; Saif, L.J. Retrospective serosurveillance of bovine norovirus (GIII.2) and nebovirus in cattle from selected feedlots and a veal calf farm in 1999 to 2001 in the United States. Arch. Virol. 2014, 159, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.I.; Han, J.I.; Wang, C.; Cooper, V.; Schwartz, K.; Engelken, T.; Yoon, K.J. Case-control study of microbiological etiology associated with calf diarrhea. Vet. Microbiol. 2013, 166, 375–385. [Google Scholar] [CrossRef]
- Wise, A.G.; Monroe, S.S.; Hanson, L.E.; Grooms, D.L.; Sockett, D.; Maes, R.K. Molecular characterization of noroviruses detected in diarrheic stools of Michigan and Wisconsin dairy calves: Circulation of two distinct subgroups. Virus Res. 2004, 100, 165–177. [Google Scholar] [CrossRef]
- Smiley, J.R.; Hoet, A.E.; Tråvén, M.; Tsunemitsu, H.; Saif, L.J. Reverse transcription-PCR assays for detection of bovine enteric caliciviruses (BEC) and analysis of the genetic relationships among BEC and human caliciviruses. J. Clin. Microbiol. 2003, 41, 3089–3099. [Google Scholar] [CrossRef]
- Pinto, P.; Wang, Q.; Chen, N.; Dubovi, E.J.; Daniels, J.B.; Millward, L.M.; Buonavoglia, C.; Martella, V.; Saif, L.J. Discovery and genomic characterization of noroviruses from a gastroenteritis outbreak in domestic cats in the us. PLoS ONE 2012, 7, e32739. [Google Scholar] [CrossRef]
- Zhang, W.; Li, L.; Deng, X.; Kapusinszky, B.; Pesavento, P.A.; Delwart, E. Faecal virome of cats in an animal shelter. J. Gen. Virol. 2014, 95, 2553–2564. [Google Scholar] [CrossRef]
- Li, L.; Shan, T.; Wang, C.; Côté, C.; Kolman, J.; Onions, D.; Gulland, F.M.D.; Delwart, E. The fecal viral flora of California sea lions. J. Virol. 2011, 85, 9909–9917. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, A.C.; Hidalgo, M.A.; Obando, C.; Vizzi, E.; Liprandi, F.; Ludert, J.E. Molecular identification of bovine enteric caliciviruses in Venezuela. Acta Cient. Venez. 2003, 54, 148–152. [Google Scholar] [PubMed]
- Ferragut, F.; Vega, C.G.; Mauroy, A.; Conceição-Neto, N.; Zeller, M.; Heylen, E.; Uriarte, E.L.; Bilbao, G.; Bok, M.; Matthijnssens, J.; et al. Molecular detection of bovine Noroviruses in Argentinean dairy calves: Circulation of a tentative new genotype. Infect. Genet. Evol. 2016, 40, 144–150. [Google Scholar] [CrossRef]
- Cunha, J.B.; de Mendonça, M.C.L.; Miagostovich, M.P.; Leite, J.P.G. First detection of porcine norovirus GII.18 in Latin America. Res. Vet. Sci. 2010, 89, 126–129. [Google Scholar] [CrossRef]
- Cunha, J.B.; de Mendonça, M.C.L.; Miagostovich, M.P.; Leite, J.P.G. Genetic diversity of porcine enteric caliciviruses in pigs raised in Rio de Janeiro State, Brazil. Arch. Virol. 2010, 155, 1301–1305. [Google Scholar] [CrossRef]
- Das Merces Hernandez, J.; Stangarlin, D.C.; Siqueira, J.A.M.; de Souza Oliveira, D.; Portal, T.M.; Barry, A.F.; Dias, F.A.; de Matos, J.C.S.; Mascarenhas, J.D.P.; Gabbay, Y.B. Genetic diversity of porcine sapoviruses in pigs from the Amazon region of Brazil. Arch. Virol. 2014, 159, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Almeida, P.R.; Lorenzetti, E.; Cruz, R.S.; Watanabe, T.T.; Zlotowski, P.; Alfieri, A.A.; Driemeier, D. Diarrhea caused by rotavirus A, B, and C in suckling piglets from southern Brazil: Molecular detection and histologic and immunohistochemical characterization. J. Vet. Diagn. Investig. 2018. [Google Scholar] [CrossRef] [PubMed]
- Castro, T.X.; Cubel Garcia, R.C.N.; Fumian, T.M.; Costa, E.M.; Mello, R.; White, P.A.; Leite, J.P.G. Detection and molecular characterization of caliciviruses (vesivirus and norovirus) in an outbreak of acute diarrhea in kittens from Brazil. Vet. J. 2015, 206, 115–117. [Google Scholar] [CrossRef]
- Guo, Z.; He, Q.; Yue, H.; Zhang, B.; Tang, C. First detection of Nebovirus and Norovirus from cattle in China. Arch. Virol. 2018, 163, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Q.; Xu, L.; Tu, C.; Huang, X.; He, B. Detection and Characterization of a Novel Norovirus in Bats, China. Virol. Sin. 2018, 33, 100–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soma, T.; Nakagomi, O.; Nakagomi, T.; Mochizuki, M. Detection of norovirus and sapovirus from diarrheic dogs and cats in Japan. Microbiol. Immunol. 2015, 59, 123–128. [Google Scholar] [CrossRef]
- Tsunesumi, N.; Sato, G.; Iwasa, M.; Kabeya, H.; Maruyama, S.; Tohya, Y. Novel murine norovirus-like genes in wild rodents in Japan. J. Vet. Med. Sci. 2012, 74, 1221–1224. [Google Scholar] [CrossRef]
- Keum, H.O.; Moon, H.J.; Park, S.J.; Kim, H.K.; Rho, S.M.; Park, B.K. Porcine noroviruses and sapoviruses on Korean swine farms. Arch. Virol. 2009, 154, 1765–1774. [Google Scholar] [CrossRef]
- Lyoo, K.S.; Jung, M.C.; Yoon, S.W.; Kim, H.K.; Jeong, D.G. Identification of canine norovirus in dogs in South Korea. BMC Vet. Res. 2018, 14, 413. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Jeong, C.; Kim, H.H.; Park, S.H.; Park, S.J.; Hyun, B.H.; Yang, D.K.; Kim, S.K.; Kang, M.I.; Cho, K.O. Molecular epidemiology of bovine noroviruses in South Korea. Vet. Microbiol. 2007, 124, 125–133. [Google Scholar] [CrossRef]
- Pourasgari, F.; Kaplon, J.; Sanchooli, A.; Fremy, C.; Karimi-Naghlani, S.; Otarod, V.; Ambert-Balay, K.; Mojgani, N.; Pothier, P. Molecular prevalence of bovine noroviruses and neboviruses in newborn calves in Iran. Arch. Virol. 2018. [Google Scholar] [CrossRef]
- Mahzounieh, M.; Moghtadaei, E.; Zahraei Salehi, T. Detection of calicivirus genomes in calves using Ni/E3 primers in Shahrekord Area, Iran. Pak. J. Biol. Sci. 2006, 9, 227–230. [Google Scholar]
- Yilmaz, H.; Turan, N.; Altan, E.; Bostan, K.; Yilmaz, A.; Helps, C.R.; Cho, K.O. First report on the phylogeny of bovine norovirus in Turkey. Arch. Virol. 2011, 156, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Thuran, T.; Irehan, B. Detection and molecular analysis of bovine enteric norovirus and nebovirus in Turkey. J. Vet. Res. 2018, 62, 129–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haga, K.; Fujimoto, A.; Takai-Todaka, R.; Miki, M.; Doan, Y.H.; Murakami, K.; Yokoyama, M.; Murata, K.; Nakanishi, A.; Katayama, K. Functional receptor molecules CD300lf and CD300ld within the CD300 family enable murine noroviruses to infect cells. Proc. Natl. Acad. Sci. USA 2016, 113, E6248–E6255. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Williamson, W.M.; Hewitt, J.; Rivera-Aban, M.; Lin, S.; Ball, A.; Scholes, P.; Greening, G.E. Sensitive multiplex real-time reverse transcription-PCR assay for the detection of human and animal noroviruses in clinical and environmental samples. Appl. Environ. Microbiol. 2007, 73, 5464–5470. [Google Scholar] [CrossRef]
- Monini, M.; Di Bartolo, I.; Ianiro, G.; Angeloni, G.; Magistrali, C.F.; Ostanello, F.; Ruggeri, F.M. Detection and molecular characterization of zoonotic viruses in swine fecal samples in Italian pig herds. Arch. Virol. 2015, 160, 2547–2556. [Google Scholar] [CrossRef]
- Di Bartolo, I.; Tofani, S.; Angeloni, G.; Ponterio, E.; Ostanello, F.; Ruggeri, F.M. Detection and characterization of porcine caliciviruses in Italy. Arch. Virol. 2014, 159, 2479–2484. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Di Felice, E.; Melegari, I.; Ceci, C.; Mauroy, A.; Thiry, E.; Martella, V.; Marsilio, F. Genetic heterogeneity of bovine noroviruses in Italy. Arch. Virol. 2014, 159, 2717–2722. [Google Scholar] [CrossRef]
- Di Bartolo, I.; Ponterio, E.; Monini, M.; Ruggeri, F.M. A pilot survey of bovine norovirus in northern Italy. Vet. Rec. 2011, 169, 73. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Sarchese, V.; Massirio, I.; Palermo, G.; Romito, G.; Lorusso, E.; Lanave, G.; Bodnar, L.; et al. Seroprevalence for norovirus genogroup II, IV and VI in dogs. Vet. Microbiol. 2017, 203, 68–72. [Google Scholar] [CrossRef]
- Di Martino, B.; Marsilio, F.; Di Profio, F.; Lorusso, E.; Friedrich, K.G.; Buonavoglia, C.; Martella, V. Detection of antibodies against Norovirus genogroup GIV in carnivores. Clin. Vaccine Immunol. 2010, 17, 180–182. [Google Scholar] [CrossRef]
- Martella, V.; Decaro, N.; Lorusso, E.; Radogna, A.; Moschidou, P.; Amorisco, F.; Lucente, M.S.; Desario, C.; Mari, V.; Elia, G.; et al. Genetic heterogeneity and recombination in canine noroviruses. J. Virol. 2009, 83, 11391–11396. [Google Scholar] [CrossRef]
- Bodnar, L.; Lorusso, E.; Di Martino, B.; Catella, C.; Lanave, G.; Elia, G.; Bányai, K.; Buonavoglia, C.; Martella, V. Identification of a novel canine norovirus. Infect. Genet. Evol. 2017, 52, 75–81. [Google Scholar] [CrossRef]
- Martella, V.; Campolo, M.; Lorusso, E.; Cavicchio, P.; Camero, M.; Bellacicco, A.L.; Decaro, N.; Elia, G.; Greco, G.; Corrente, M.; et al. Norovirus in captive lion cub (Panthera leo). Emerg. Infect. Dis. 2007, 13, 1071–1073. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Sarchese, V.; Cafiero, M.A.; Robetto, S.; Aste, G.; Lanave, G.; Marsilio, F.; Martella, V. A novel feline norovirus in diarrheic cats. Infect. Genet. Evol. 2016, 38, 132–137. [Google Scholar] [CrossRef]
- Halaihel, N.; Masía, R.M.; Fernández-Jiménez, M.; Ribes, J.M.; Montava, R.; De Blas, I.; Gironés, O.; Alonso, J.L.; Buesa, J. Enteric calicivirus and rotavirus infections in domestic pigs. Epidemiol. Infect. 2010, 138, 542–548. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Nascimento, M.S. Molecular epidemiology of canine norovirus in dogs from Portugal, 2007–2011. BMC Vet. Res. 2012, 8, 107. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Nascimento, M.S.J. Gastroenteritis outbreak associated with faecal shedding of canine norovirus in a portuguese kennel following introduction of imported dogs from russia. Transbound. Emerg. Dis. 2012, 59, 456–459. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Barclay, L.; Nascimento, M.S.J.; Vinjé, J. Novel norovirus in dogs with diarrhea. Emerg. Infect. Dis. 2010, 16, 980–982. [Google Scholar] [CrossRef]
- Ntafis, V.; Xylouri, E.; Radogna, A.; Buonavoglia, C.; Martella, V. Outbreak of canine norovirus infection in young dogs. J. Clin. Microbiol. 2010, 48, 2605–2608. [Google Scholar] [CrossRef]
- Kaplon, J.; Guenau, E.; Asdrubal, P.; Pothier, P.; Ambert-Balay, K. Possible novel Nebovirus genotype in cattle, France. Emerg. Infect. Dis. 2011, 17, 1120–1123. [Google Scholar] [CrossRef]
- Kaplon, J.; Fremy, C.; Bernard, S.; Rehby, L.; Aho, S.; Pothier, P.; Ambert-Balay, K. Impact of rotavirus vaccine on rotavirus genotypes and caliciviruses circulating in French cattle. Vaccine 2013, 31, 2433–2440. [Google Scholar] [CrossRef]
- Oliver, S.L.; Wood, E.; Asobayire, E.; Wathes, D.C.; Brickell, J.S.; Elschner, M.; Otto, P.; Lambden, P.R.; Clarke, I.N.; Bridger, J.C. Serotype 1 and 2 bovine noroviruses are endemic in cattle in the United Kingdom and Germany. J. Clin. Microbiol. 2007, 45, 3050–3052. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Batten, C.A.; Liu, B.L.; Lambden, P.R.; Elschner, M.; Günther, H.; Otto, H.P.; Schnürch, P.; Eichhorn, W.; Herbst, W.; et al. Studies of epidemiology and seroprevalence of bovine Noroviruses in Germany. J. Clin. Microbiol. 2003, 41, 2300–2305. [Google Scholar] [CrossRef]
- Ike, A.C.; Roth, B.N.; Böhm, R.; Pfitzner, A.J.; Marschang, R.E. Identification of bovine enteric Caliciviruses (BEC) from cattle in Baden-Württemberg. DTW Dtsch Tierarztl Wochenschr 2007, 114, 12–15. [Google Scholar]
- Sachsenröder, J.; Braun, A.; Machnowska, P.; Ng, T.F.F.; Deng, X.; Guenther, S.; Bernstein, S.; Ulrich, R.G.; Delwart, E.; Johne, R. Metagenomic identification of novel enteric viruses in urban wild rats and genome characterization of a group A rotavirus. J. Gen. Virol. 2014, 95, 2734–2747. [Google Scholar] [CrossRef] [Green Version]
- Van Der Poel, W.H.M.; Van Der Heide, R.; Verschoor, F.; Gelderblom, H.; Vinjé, J.; Koopmans, M.P.G. Epidemiology of Norwalk-like virus infections in cattle in The Netherlands. Vet. Microbiol. 2003, 92, 297–309. [Google Scholar] [CrossRef]
- De Graaf, M.; Bodewes, R.; van Elk, C.E.; van de Bildt, M.; Getu, S.; Aron, G.I.; Verjans, G.M.G.M.; Osterhaus, A.D.M.E.; van den Brand, J.M.A.; Kuiken, T.; et al. Norovirus infection in harbor porpoises. Emerg. Infect. Dis. 2017, 23, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Mauroy, A.; Scipioni, A.; Mathijs, E.; Miry, C.; Ziant, D.; Thys, C.; Thiry, E. Noroviruses and sapoviruses in pigs in Belgium. Arch. Virol. 2008, 153, 1927–1931. [Google Scholar] [CrossRef]
- Mauroy, A.; Scipioni, A.; Mathijs, E.; Saegerman, C.; Mast, J.; Bridger, J.C.; Ziant, D.; Thys, C.; Thiry, E. Epidemiological study of bovine norovirus infection by RT-PCR and a VLP-based antibody ELISA. Vet. Microbiol. 2009, 137, 243–251. [Google Scholar] [CrossRef]
- Mauroy, A.; Scipioni, A.; Mathijs, E.; Ziant, D.; Daube, G.; Thirya, E. Complete genome sequence of a novel bovine norovirus: Evidence for slow genetic evolution in genogroup III genotype 2 noroviruses. J. Virol. 2012, 86, 12449–12450. [Google Scholar] [CrossRef]
- Mauroy, A.; Scipioni, A.; Mathijs, E.; Thys, C.; Thiry, E. Molecular detection of kobuviruses and recombinant noroviruses in cattle in continental Europe. Arch. Virol. 2009, 154, 1841–1845. [Google Scholar] [CrossRef] [PubMed]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med. Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef]
- Milnes, A.S.; Binns, S.H.; Oliver, S.L.; Bridger, J.C. Retrospective study of noroviruses in samples of diarrhoea from cattle, using the Veterinary Laboratories Agency’s Farmfile database. Vet. Rec. 2007, 160, 326–330. [Google Scholar] [CrossRef]
- Caddy, S.; Emmott, E.; El-Attar, L.; Mitchell, J.; De Rougemont, A.; Brownlie, J.; Goodfellow, I. Serological evidence for multiple strains of canine norovirus in the UK dog population. PLoS ONE 2013, 8, e81596. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.L.; De Rougemont, A.; Emmott, E.; El-Attar, L.; Mitchell, J.A.; Hollinshead, M.; Belliot, G.; Brownlie, J.; Le Pendu, J.; Goodfellow, I. Evidence for human norovirus infection of dogs in the United Kingdom. J. Clin. Microbiol. 2015, 53, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; McFadden, N.; Blundell, R.J.; Meredith, A.; Simmonds, P. Diversity of murine norovirus in wild-rodent populations: Species-specific associations suggest an ancient divergence. J. Gen. Virol. 2012, 93, X259–266. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Reetz, J.; Johne, R.; Heiberg, A.C.; Petri, S.; Kanig, H.; Ulrich, R.G. The simultaneous occurrence of human norovirus and hepatitis E virus in a Norway rat (Rattus norvegicus). Arch. Virol. 2013, 158, 1575–1578. [Google Scholar] [CrossRef]
- Jor, E.; Myrmel, M.; Jonassen, C.M. SYBR Green based real-time RT-PCR assay for detection and genotype prediction of bovine noroviruses and assessment of clinical significance in Norway. J. Virol. Methods 2010, 169, 1–7. [Google Scholar] [CrossRef]
- Summa, M.; von Bonsdorff, C.H.; Maunula, L. Pet dogs-A transmission route for human noroviruses? J. Clin. Virol. 2012, 53, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Summa, M.; Henttonen, H.; Maunula, L. Human noroviruses in the faeces of wild birds and rodents-new potential transmission routes. Zoonoses Public Health 2018. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Bíró, H.; Szucs, G. Enteric caliciviruses in domestic pigs in Hungary. Arch. Virol. 2007, 152, 611–614. [Google Scholar] [CrossRef]
- Mihalov-Kovács, E.; Marton, S.; Fehér, E.; Lengyel, G.; Jakab, F.; Tuboly, T.; Bányai, K. Enteric viral infections of sheltered dogs in Hungary. Magy. Allatorv. Lapja 2014, 136, 661–670. [Google Scholar]
- Farkas, T.; Fey, B.; Keller, G.; Martella, V.; Egyed, L. Molecular detection of murine noroviruses in laboratory and wild mice. Vet. Microbiol. 2012, 160, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, F.F.; Mansour, S.M.; El-Araby, I.E.; Mor, S.K.; Goyal, S.M. Molecular detection of enteric viruses from diarrheic calves in Egypt. Arch. Virol. 2017, 162, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Hassine-Zaafrane, M.; Kaplon, J.; Sdiri-Loulizi, K.; Aouni, Z.; Pothier, P.; Aouni, M.; Ambert-Balay, K. Molecular prevalence of bovine noroviruses and neboviruses detected in central-eastern Tunisia. Arch. Virol. 2012, 157, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Taku, O.; Iweriebor, B.C.; Nwodo, U.U.; Obi, L.C.; Okoh, A.I. Occurrence of Norovirus in pig faecal samples in the Eastern Cape, South Africa. Asian Pac. J. Trop. Dis. 2017, 7, 151–155. [Google Scholar] [CrossRef]
- Günther, H.; Otto, P.; Heilmann, P. Diarrhea in young calves. 6. Determination of the pathogenicity of a bovine coronavirus and an unidentified icosahedral virus. Arch. Exp. Vet. 1984, 38, 781–792. [Google Scholar]
- Jung, K.; Scheuer, K.A.; Zhang, Z.; Wang, Q.; Saif, L.J. Pathogenesis of GIII.2 bovine norovirus, CV186-OH/00/US strain in gnotobiotic calves. Vet. Microbiol. 2014, 168, 202–207. [Google Scholar] [CrossRef]
- Otto, P.H.; Clarke, I.N.; Lambden, P.R.; Salim, O.; Reetz, J.; Liebler-Tenorio, E.M. Infection of calves with bovine norovirus GIII.1 strain Jena virus: An experimental model to study the pathogenesis of norovirus infection. J. Virol. 2011, 85, 12013–12021. [Google Scholar] [CrossRef] [PubMed]
- Han, M.G.; Wang, Q.; Smiley, J.R.; Change, K.O.; Saif, L.J. Self-assembly of the recombinant capsid protein of a bovine norovirus (BoNV) into virus-like particles and evaluation of cross-reactivity of BoNV with human noroviruses. J. Clin. Microbiol. 2005, 43, 778–785. [Google Scholar] [CrossRef]
- Han, M.G.; Cheetham, S.; Azevedo, M.; Thomas, C.; Saif, L.J. Immune responses to bovine norovirus-like particles with various adjuvants and analysis of protection in gnotobiotic calves. Vaccine 2006, 24, 317–326. [Google Scholar] [CrossRef]
- Souza, M.; Azevedo, M.S.P.; Jung, K.; Cheetham, S.; Saif, L.J. Pathogenesis and immune responses in gnotobiotic calves after infection with the genogroup II.4-HS66 strain of human norovirus. J. Virol. 2008, 82, 1777–1786. [Google Scholar] [CrossRef]
- Oliver, S.L.; Asobayire, E.; Charpilienne, A.; Cohen, J.; Bridger, J.C. Complete genomic characterization and antigenic relatedness of genogroup III, genotype 2 bovine noroviruses. Arch. Virol. 2007, 152, 257–272. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, G.; Iaconelli, M.; Pourshaban, M.; Fratini, M. Molecular detection and genetic diversity of norovirus genogroup IV: A yearlong monitoring of sewage throughout Italy. Arch. Virol. 2010, 155, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, J.; de Graaf, M.; Al-Hello, H.; Allen, D.J.; Ambert-Balay, K.; Botteldoorn, N.; Brytting, M.; Buesa, J.; Cabrerizo, M.; Chan, M.; et al. NoroNet, Molecular surveillance of norovirus, 2005–2016: An epidemiological analysis of data collected from the NoroNet network. Lancet Infect. Dis. 2018, 18, 545–553. [Google Scholar] [CrossRef]
- Takano, T.; Kusuhara, H.; Kuroishi, A.; Takashina, M.; Doki, T.; Nishinaka, T.; Hohdatsu, T. Molecular characterization and pathogenicity of a genogroup GVI feline norovirus. Vet. Microbiol. 2015, 178, 201–207. [Google Scholar] [CrossRef]
- Takano, T.; Hiramatsu, K.; Matsuyama, M.; Mutoh, K.; Matsumoto, Y.; Fukushima, T.; Doki, T.; Kusuhara, H.; Hohdatsu, T. Viral shedding and clinical status of feline-norovirus-infected cats after reinfection with the same strain. Arch. Virol. 2018, 163, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Kocher, J.F.; Lindesmith, L.C.; Huynh, J.; Gates, J.E.; Debbink, K.; Beall, A.; Mallory, M.L.; Donaldson, E.F.; Barica, R.S. Bat Caliciviruses and Human Noroviruses Are Antigenically Similar and Have Overlapping Histo-Blood Group Antigen Binding Profile. MBio 2018, 9, e00869-18. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Ren, X.; He, G.; Zhang, J.; Yang, J.; Qian, Z.; Dong, J.; Sun, L.; Zhu, Y.; et al. Deciphering the bat virome catalog to better understand the ecological diversity of bat viruses and the bat origin of emerging infectious diseases. ISME J. 2016, 10, 609–620. [Google Scholar] [CrossRef]
- Humphrey, T.J. An outbreak of calicivirus associated gastroenteritis in an elderly persons home. A possible zoonosis? J. Hyg. (Lond) 1984, 93, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widdowson, M.A.; Rockx, B.; Schepp, R.; Van Der Poel, W.H.M.; Vinje, J.; Van Duynhoven, Y.T.; Koopmans, M.P. Detection of serum antibodies to bovine norovirus in veterinarians and the general population in The Netherlands. J. Med. Virol. 2005, 76, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Bucardo, F.; González, F.; Reyes, Y.; Blandón, P.; Saif, L.; Nordgren, J. Seroprevalence in Household Raised Pigs Indicate High Exposure to GII Noroviruses in Rural Nicaragua. Zoonoses Public Health 2016, 63, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.W.; Hu, Y.; Chen, H.; Podkolzin, A.T.; Zaytseva, E.V.; Komano, J.; Sakon, N.; Poovorawan, Y.; Vongpunsawad, S.; Thanusuwannasak, T.; et al. Global Spread of Norovirus GII.17 Kawasaki 308, 2014–2016. Emerg. Infect. Dis. 2017, 23, 1350–1354. [Google Scholar] [CrossRef]
- De Graaf, M.; van Beek, J.; Vennema, H.; Podkolzin, A.T.; Hewitt, J.; Bucardo, F.; Templeton, K.; Nordgren, J.; Reuter, G.; Lynch, M.; et al. Emergence of a novel GII.17 norovirus - End of the GII.4 era? Euro Surveill 2015, 20, 1–8. [Google Scholar] [CrossRef]
- He, Z.; Liu, B.; Tao, Y.; Li, C.; Xia, M.; Zhong, W.; Jiang, X.; Liu, H.; Tan, M. Norovirus GII.17 natural infections in Rhesus monkeys, China. Emerg. Infect. Dis. 2017, 23, 316–319. [Google Scholar] [CrossRef]
- Liu, B.; Tao, Y.; Li, C.; Li, X.; Liu, J.; He, Z.; Xia, M.; Jiang, X.; Tan, M.; Liu, H. Complete Genome Sequence of a GII.17 Norovirus Isolated from a Rhesus Monkey in China. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T.; Cross, R.W.; Hargitt, I.E.; Lerche, N.W.; Morrow, A.L.; Sestak, K. Genetic diversity and histo-blood group antigen interactions of rhesus enteric caliciviruses. J. Virol. 2010, 84, 8617–8625. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T. Natural norovirus infections in rhesus macaques. Emerg. Infect. Dis. 2016, 22, 1272–1274. [Google Scholar] [CrossRef]
- Jiang, B.; McClure, H.M.; Fankhauser, R.L.; Monroe, S.S.; Glass, R.I. Prevalence of rotavirus and norovirus antibodies in non-human primates. J. Med. Primatol. 2004, 33, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T.; Dofour, J.; Jiang, X.; Sestak, K. Detection of norovirus-, sapovirus- and rhesus enteric calicivirus-specific antibodies in captive juvenile macaques. J. Gen. Virol. 2010, 91, 734–738. [Google Scholar] [CrossRef]
- Rockx, B.H.G.; Bogers, W.M.J.M.; Heeney, J.L.; van Amerongen, G.; Koopmans, M.P.G. Experimental norovirus infections in non-human primates. J. Med. Virol. 2005, 75, 313–320. [Google Scholar] [CrossRef]
- De Graaf, M.; van Beek, J.; Koopmans, M.P.G. Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 2016, 14, 421–433. [Google Scholar] [CrossRef]
- Taube, S.; Kolawole, A.O.; Höhne, M.; Wilkinson, J.E.; Handley, S.A.; Perry, J.W.; Thackray, L.B.; Akkina, R.; Wobus, C.E. A mouse model for human norovirus. mBio 2013, 4. [Google Scholar] [CrossRef]
- Seo, D.J.; Jung, D.; Jung, S.; Ha, S.K.; Ha, S.D.; Choi, I.S.; Myoung, J.; Choi, C. Experimental miniature piglet model for the infection of human norovirus GII. J. Med. Virol. 2018, 90, 655–662. [Google Scholar] [CrossRef]
- Cheetham, S.; Souza, M.; McGregor, R.; Meulia, T.; Wang, Q.; Saif, L.J. Binding patterns of human norovirus-like particles to buccal and intestinal tissues of gnotobiotic pigs in relation to A/H histo-blood group antigen expression. J. Virol. 2007, 81, 3535–3544. [Google Scholar] [CrossRef]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinjé, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell–derived human enteroids. Science 2016, 353, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Duizer, E.; Schwab, K.J.; Neill, F.H.; Atmar, R.L.; Koopmans, M.P.G.; Estes, M.K. Laboratory efforts to cultivate noroviruses. J. Gen. Virol. 2004, 85, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Stoltzfus, G.T.; Zhu, C.; Jung, K.; Wang, Q.; Saif, L.J. Attempts to grow human noroviruses, a sapovirus, and a bovine norovirus in vitro. PLoS ONE 2018, 13, e0178157. [Google Scholar] [CrossRef] [PubMed]
- Lou, F.; Ye, M.; Ma, Y.; Li, X.; DiCaprio, E.; Chen, H.; Krakowka, S.; Hughes, J.; Kingsley, D.; Li, J. A Gnotobiotic Pig Model for Determining Human Norovirus Inactivation by High-Pressure Processing. Appl. Environ. Microbiol. 2015, 81, 6679–6687. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.; Kocher, J.; Li, Y.; Wen, K.; Li, G.; Liu, F.; Yang, X.; Leroith, T.; Tan, M.; Xia, M.; et al. Median infectious dose of human norovirus GII.4 in gnotobiotic pigs is decreased by simvastatin treatment and increased by age. J. Gen. Virol. 2013, 94, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Kocher, J.; Bui, T.; Giri-Rachman, E.; Wen, K.; Li, G.; Yang, X.; Liu, F.; Tan, M.; Xia, M.; Zhong, W.; et al. Intranasal P particle vaccine provided partial cross-variant protection against human GII.4 Norovirus diarrhea in gnotobiotic pigs. J. Virol. 2014, 8, 9728–9743. [Google Scholar] [CrossRef]
- Lei, S.; Ramesh, A.; Twitchell, E.; Wen, K.; Bui, T.; Weiss, M.; Yang, X.; Kocher, J.; Li, G.; Giri-Rachman, E.; et al. High protective efficacy of probiotics and rice bran against human norovirus infection and diarrhea in gnotobiotic pigs. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Ryu, J.; Wen, K.; Twitchell, E.; Bui, T.; Ramesh, A.; Weiss, M.; Li, G.; Samuel, H.; Clark-Deener, S.; et al. Increased and prolonged human norovirus infection in RAG2/IL2RG deficient gnotobiotic pigs with severe combined immunodeficiency. Sci. Rep. 2016, 6, 25222. [Google Scholar] [CrossRef] [Green Version]
- Takanashi, S.; Wang, Q.; Chen, N.; Shen, Q.; Jung, K.; Zhang, Z.; Yokoyama, M.; Lindesmith, L.C.; Baric, R.S.; Saif, L.J. Characterization of emerging GII.g/GII.12 noroviruses from a gastroenteritis outbreak in the United States in 2010. J. Clin. Microbiol. 2011, 49, 3234–3244. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.; Cheetham, S.M.; Azevedo, M.S.P.; Costantini, V.; Saif, L.J. Cytokine and antibody responses in gnotobiotic pigs after infection with human norovirus genogroup II.4 (HS66 strain). J. Virol. 2007, 81, 9183–9192. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Samuel, H.; Twitchell, E.; Bui, T.; Ramesh, A.; Wen, K.; Weiss, M.; Li, G.; Yang, X.; Jiang, X.; et al. Enterobacter cloacae inhibits human norovirus infectivity in gnotobiotic pigs. Sci. Rep. 2016, 6, 25017. [Google Scholar] [CrossRef] [Green Version]
- Cheetham, S.; Menira, S.; Meulia, T.; Grimes, S.; Han, M.G.; Saif, L.J. Pathogenesis of a genogroup II human norovirus in gnotobiotic pigs. J. Virol. 2006, 80, 10372–10381. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Jung, S.T.; Choi, C.S.; Myoung, J.; Ahn, H.S.; Han, S.H.; Kim, Y.H.; Go, H.J.; Lee, J.B.; Park, S.Y.; et al. Pathogenesis of Human Norovirus Genogroup II Genotype 4 in PostWeaning Gnotobiotic Pigs. J. Microbiol. Biotechnol. 2018, 28, 2133–2140. [Google Scholar] [CrossRef]
- Sestak, K.; Feely, S.; Fey, B.; Dufour, J.; Hargitt, E.; Alvarez, X.; Pahar, B.; Gregoricus, N.; Vinjé, J.; Farkas, T. Experimental inoculation of juvenile rhesus macaques with primate enteric caliciviruses. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Subekti, D.S.; Tjaniadi, P.; Lesmana, M.; McArdle, J.; Iskandriati, D.; Budiarsa, I.N.; Walujo, P.; Suparto, I.H.; Winoto, I.; Campbell, J.R.; et al. Experimental infection of Macaca nemestrina with a Toronto Norwalk-like virus of epidemic viral gastroenteritis. J. Med. Virol. 2002, 66, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Bok, K.; Parra, G.I.; Mitra, T.; Abente, E.; Shaver, C.K.; Boon, D.; Engle, R.; Yu, C.; Kapikian, A.Z.; Sosnovtsev, S.V.; et al. Chimpanzees as an animal model for human norovirus infection and vaccine development. Proc. Natl. Acad. Sci. USA 2011, 108, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, R.G.; Greenberg, H.B.; Dalgard, D.W.; Allen, W.P.; Sly, D.L.; Thornhill, T.S.; Chanock, R.M.; Kapikian, A.Z. Experimental infection of chimpanzees with the Norwalk agent of epidemic viral gastroenteritis. J. Med. Virol. 1978, 2, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Marionneau, S.; Cailleau-Thomas, A.; Rocher, J.; Le Moullac-Vaidye, B.; Ruvoën, N.; Monique, C.; Le Pendu, J. ABH and Lewis histo-blood group antigens, a model for the meaning of oligosaccharide diversity in the face of a changing world. Biochimie 2004, 83, 565–573. [Google Scholar] [CrossRef]
- Huang, P.; Farkas, T.; Marionneau, S.; Zhong, W.; Ruvoën-Clouet, N.; Morrow, A.L.; Altaye, M.; Pickering, L.K.; Newburg, D.S.; LePendu, J.; et al. Noroviruses Bind to Human ABO, Lewis, and Secretor Histo–Blood Group Antigens: Identification of 4 Distinct Strain-Specific Patterns. J. Infect. Dis. 2002, 188, 19–31. [Google Scholar] [CrossRef]
- Ming, T.; Jiang, X. Histo-blood group antigens: A common niche for norovirus and rotavirus. Expert Rev. Mol. Med. 2014, 16, e5. [Google Scholar]
- Tan, M.; Jiang, X. Norovirus Gastroenteritis, Carbohydrate Receptors, and Animal Models. PLoS Pathog. 2010, 6, e1000983. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Farkas, T.; Zhong, W.; Tan, M.; Thornton, S.; Morrow, A.L.; Jiang, X. Norovirus and Histo-Blood Group Antigens: Demonstration of a Wide Spectrum of Strain Specificities and Classification of Two Major Binding Groups among Multiple Binding Patterns. J. Virol. 2005, 79, 6714–6722. [Google Scholar] [CrossRef] [Green Version]
- Marionneau, S.; Ruvoën, N.; Le Moullac-Vaidye, B.; Clement, M.; Cailleau-Thomas, A.; Ruiz-Palacois, G.; Huang, P.; Jiang, X.; Le Pendu, J. Norwalk Virus Binds to Histo-Blood Group Antigens Present on Gastroduodenal Epithelial Cells of Secretor Individuals. Gastroenterology 2002, 122, 1967–1977. [Google Scholar] [CrossRef]
- Tamura, M.; Natori, K.; Kobayashi, M.; Miyamura, T.; Takeda, N. Genogroup II Noroviruses Efficiently Bind to Heparan Sulfate Proteoglycan Associated with the Cellular Membrane. J. Virol. 2004, 78, 3817–3826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rydell, G.E.; Nilsson, J.; Rodriguez-Diaz, J.; Ruvoen-Clouet, N.; Svensson, L.; Le Pendu, J.; Larson, G. Human noroviruses recognize sialyl Lewis x neoglycoprotein. Glycobiology 2009, 19, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Tan, M.; Xia, M.; Kitova, E.N.; Jiang, X.; Klassen, J.S. Gangliosides are ligands for human noroviruses. J. Am. Chem. Soc. 2014, 136, 12631–12637. [Google Scholar] [CrossRef] [PubMed]
- Almand, E.A.; Moore, M.D.; Jaykus, L.A. Norovirus Binding to Ligands Beyond Histo-Blood Group Antigens. Front. Microbiol. 2017, 8, 2549. [Google Scholar] [CrossRef] [Green Version]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Host and viral traits predict zoonotic spillover from mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.; Breiman, A.; le Pendu, J.; Goodfellow, I. Genogroup IV and VI canine noroviruses interact with histo-blood group antigens. J. Virol. 2014, 88, 10377–10391. [Google Scholar] [CrossRef] [PubMed]
- Zakhour, M.; Ruvoën-Clouet, N.; Charpilienne, A.; Langpap, B.; Poncet, D.; Peters, T.; Bovin, N.; Le Pendu, J. The αGal epitope of the histo-blood group antigen family is a ligand for bovine norovirus Newbury2 expected to prevent cross-species transmission. PLoS Pathog. 2009, 5, e1000504. [Google Scholar] [CrossRef]
- Taube, S.; Perry, J.W.; McGreevy, E.; Yetming, K.; Perkins, C.; Henderson, K.; Wobus, C.E. Murine Noroviruses Bind Glycolipid and Glycoprotein Attachment Receptors in a Strain-Dependent Manner. J. Virol. 2012, 86, 5584–5593. [Google Scholar] [CrossRef] [Green Version]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldridge, M.T.; McCune, B.T.; Lee, Y.C.J.; Lee, S.; Pruett-Miller, S.M.; Nelson, C.A.; Fremont, D.H.; et al. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, F.; Cid, E.; Yamamoto, M.; Saitou, N.; Bertranpetit, J.; Blancher, A. An integrative evolution theory of histo-blood group ABO and related genes. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef]
- Jones, K.E.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Lowther, J.A.; Henshilwood, K.; Lees, D.N.; Hill, V.R.; Vinjé, J. Rapid and sensitive detection of noroviruses by using TaqMan-based one-step reverse transcription-PCR assays and application to naturally contaminated shellfish samples. Appl. Environ. Microbiol. 2005, 71, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Parra, G.I.; Bok, K.; Taylor, R.; Haynes, J.R.; Sosnovtsev, S.V.; Richardson, C.; Green, K.Y. Immunogenicity and specificity of norovirus Consensus GII.4 virus-like particles in monovalent and bivalent vaccine formulations. Vaccine 2012, 30, 3580–3586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, M.K.; Balasubramanian, G.B.; Sahoo, G.C.; Bhattacharya, R.; Vinje, J.; Kobayashi, N.; Sarkar, M.C.; Bhattacharya, M.K.; Krishnan, T. Detection of a novel intergenogroup recombinant Norovirus from Kolkata, India. Virology 2008, 377, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, G.P.; Kaneshi, K.; Ueda, Y.; Nakaya, S.; Nishimura, S.; Yamamoto, A.; Sugita, K.; Takanashi, S.; Okitsu, S.; Ushijima, H. Genetic heterogeneity, evolution, and recombination in noroviruses. J. Med. Virol. 2007, 79, 1388–1400. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; Loisy, F.; Atmar, R.L.; Hutson, A.M.; Estes, M.K.; Ruvoën-Clouet, N.; Pommepuy, M.; Le Pendu, J. Norwalk virus-specific binding to oyster digestive tissues. Emerg. Infect. Dis. 2006, 12, 931–936. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; Bon, F.; DeMedici, D.; Parnaudeau, S.; Bertone, A.; Crudeli, S.; Doyle, A.; Zidane, M.; Suffredini, E.; Kohli, E.; et al. Detection of multiple noroviruses associated with an international gastroenteritis outbreak linked to oyster consumption. J. Clin. Microbiol. 2006, 44, 3878–3882. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; Atmar, R.L.; Le Pendu, J. Transmission of viruses through shellfish: When specific ligands come into play. Curr. Opin. Virol. 2012, 2, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Costantini, V.; Loisy, F.; Joens, L.; Le Guyader, F.S.; Saif, L.J. Human and animal enteric caliciviruses in oysters from different coastal regions of the United States. Appl. Environ. Microbiol. 2006, 72, 1800–1809. [Google Scholar] [CrossRef]
- Becker-Dreps, S.; Cuthbertson, C.C.; Bucardo, F.; Vinje, J.; Paniagua, M.; Giebultowicz, S.; Espinoza, F.; Emch, M. Environmental factors associated with childhood norovirus diarrhoea in León, Nicaragua. Epidemiol. Infect. 2017, 145, 1597–1605. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Host | Genotype | Prevalence in % (References) | ||
|---|---|---|---|---|---|
| Serology | Feces | ||||
| The Americas | USA | Pigs | GII.18, GII.11, GII.19 | 71 [46] | 0–19 [25,28,46,48] |
| Cattle | GIII.1, GIII.2 | 100 [49] | 29–72 [50,51,52] | ||
| Cats | GIV.2 | 17–43 [53,54] | |||
| Sea lion | GII/GIV | 9 [55] | |||
| Canada | Pigs | GII, GII.11, GII.18 | 2–85 [30,31,32] | ||
| Cattle | GIII.2 | 1 [30] | |||
| Venezuela | Pigs | all | 0 [39] | ||
| Cattle | GIII | 0.7 [56] | |||
| Argentina | Cattle | GIII.1, GIII.2 | 3 [57] | ||
| Brazil | Pigs | GII.11, GII.18, GII.19 | 0–52 [44,58,59,60,61] | ||
| Cats | GIV,2 | 3 [62] | |||
| Asia/New Zealand | China | Pigs | GII.11, GII.18, GII.19 | 0–17 [26,27,33,35] | |
| Cattle | GIII.1 | 11 [63] | |||
| Bats | NA | 3–4 [6,64] | |||
| Taiwan | Pigs | GII.11 | 1.6 [34] | ||
| Japan | Pigs | GII.11 | 36 [46] | 0.4–15 [10,36,45] | |
| Dogs | GIV | 2 [65] | |||
| Cats | GIV.2 | 1.2 [65] | |||
| Rodents | GV | 0–14 [66] | |||
| Korea | Pigs | GII.11, GII.18 | 0.5–2 [37,67] | ||
| Dogs | Canine norovirus | 16 [68] | 3 [68] | ||
| Cattle | GIII.1, GIII.2 | 9 [69] | |||
| Iran | Cattle | GIII.1, GIII.2 | 18–40 [70,71] | ||
| Turkey | Cattle | GIII.2 | 4–9 [72,73] | ||
| India | Cattle | GIII.1 | 0.4 [74] | ||
| New Zealand | Pigs | GII.11 | 9 [38] | ||
| Cattle | GIII.1 | 54 [75] | |||
| Sheep | GIII.3 | 24 [38] | |||
| Europe | Italy | Pigs | GII.11 | 0–0.5 [76,77] | |
| Cattle | GIII.1, GIII.2 | 11–21 [78,79] | |||
| Dogs | GIV, GVI | 5–60 [47,80,81] | 2–5 [82,83] | ||
| Lion | GIV.2 | 100 [84] | |||
| Cats | GIV.2 | 16 [85] | 3 [81] | ||
| Spain | Pigs | all | 12 [86] | ||
| Dogs | GVI | 8 [83] | |||
| Portugal | Dogs | GIV, GVI | 64 [47] | 23–28 [87,88,89] | |
| Greece | Dogs | GIV.2 | 8 [90] | ||
| France | Cattle | GIII.1, GIII.2 | 20–37 [91,92] | ||
| Dogs | GVI.2 | 20 [47,83] | 0 [83] | ||
| Switzerland | Dogs | GVI.2 | 20 [47] | ||
| Germany | Pigs | GII.18 | 14 [41] | ||
| Cattle | GIII.1, GIII.2 | 66–99 [93,94] | 93 [95] | ||
| Dogs | GIV, GVI.2 | 16 [47] | 4 [83] | ||
| Rodents | GV | 10 [96] | |||
| Netherlands | Pigs | GII.11 | 2 [9] | ||
| Cattle | GIII.2 | 0–44 [9] | 4 [97] | ||
| Dogs | GVI.2 | 34 [47] | |||
| Porpoise | not classified yet | 24 [98] | 10 [98] | ||
| Belgium | Pigs | GII.19 | 4.6 [99] | ||
| Cattle | GIII.2 | 93 [100] | 4–9 [80,100,101,102] | ||
| UK | Cattle | GIII.1, GIII.2 | 66–98 [93,103] | 11 [104] | |
| Dogs | GIV, GVI, GVII | 45–48 [47,105,106] | 0 [106] | ||
| Rodents | GV | 22–67 [107] | |||
| Ireland | Pigs | none | 0 [40] | ||
| Dogs | none | 0 [47] | |||
| Denmark | Dogs | GVI.2 | 20 [47] | ||
| Rodents | none | 0 [108] | |||
| Sweden | Dogs | GVI.2 | 40 [47] | ||
| Norway | Cattle | GIII.1, GIII.2 | 50 [109] | ||
| Dogs | GVI.2 | 32 [47] | |||
| Finland | Dogs | GVI.2 | 70 [47] | 0 [110] | |
| Rodents | none | 0 [111] | |||
| Poland | Dogs | GIV.2 | 32 [47] | ||
| Slovenia | Pigs | GII.11, GII.18 | 1.2 [42] | ||
| Cattle | GIII.2 | 2 [42] | |||
| Hungary | Pigs | GII.11 | 6 [112] | ||
| Dogs | GVI | 0 [47] | 3 [113] | ||
| Rodents | GV | 24–67 [114] | |||
| Africa | Egypt | Cattle | GIII.2 | 24 [115] | |
| Tunisia | Cattle | GIII.2 | 17 [116] | ||
| South Africa | Pigs | none | 0 [117] | ||
| Ethiopia | Pigs | GII.1 | 0 [43] | ||
| Gnotobiotic Calf [123] | Gnotobiotic Pig [151,152,153,154,155,156,157,158,159,160] | Mini Piglet [145] | Rhesus Macaque [136,142,161] | Pigtail Macaque [162] | Chimpanzee [163,164] | Balb/c RAG/γc−/− Mouse [144] | |
|---|---|---|---|---|---|---|---|
| Virus | GII.4 | GII.4, GII.12 | GII.3 | GI.1, GII.2, GII.4, GII.17 | GII.3 | GI.1 | GII.4 |
| Inoculation (route and virus quantity) | Oral 1.62 × 107 genomes | Oral/intranasal 104–1010 genomes | Intragastric 107 genomes | Oral/intragastric 105–106 genomes | Nasogastric, Quantity not clear | Intravenous/intragastric 4 × 106–4 × 108 genomes | Intraperitoneal 4 × 103–7 × 104 genomes |
| Shedding | 3 days | 2–16 days | 7 days | 1–19 days | Up to 21 days | 2 days–17 weeks | No shedding 1 |
| Seroconversion | Yes | Yes | NA | Yes/no 2 | Yes | Yes | No |
| Pathology | Lesions, mild villous atrophy, enterocyte vacuolization in small intestine | Increase in inflammatory cells in LM, necrosis, shortening of villous tips | No damage | No damage | NA | No damage | No damage |
| Tropism (detection of viral antigen or genome) | Positive enterocytes in the ileum, cells in LM | Enterocytes and cells in LM of duodenum, jejunum, ileum. Spleen and MLN | Immune cells in the small/large intestine, tonsils, spleen, lymph nodes, MLN | NA | NA | Cells in LM of duodenum and jejunum | Macrophage-like cells in liver and spleen. Viral genomes detected in various tissue 3 |
| Disease | Diarrhea | Diarrhea | Diarrhea | Asymptomatic | Diarrhea | Asymptomatic | Asymptomatic |
| Viremia | Yes (low) | Yes | Yes | NA | NA | NA | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villabruna, N.; Koopmans, M.P.G.; de Graaf, M. Animals as Reservoir for Human Norovirus. Viruses 2019, 11, 478. https://doi.org/10.3390/v11050478
Villabruna N, Koopmans MPG, de Graaf M. Animals as Reservoir for Human Norovirus. Viruses. 2019; 11(5):478. https://doi.org/10.3390/v11050478
Chicago/Turabian StyleVillabruna, Nele, Marion P. G. Koopmans, and Miranda de Graaf. 2019. "Animals as Reservoir for Human Norovirus" Viruses 11, no. 5: 478. https://doi.org/10.3390/v11050478
APA StyleVillabruna, N., Koopmans, M. P. G., & de Graaf, M. (2019). Animals as Reservoir for Human Norovirus. Viruses, 11(5), 478. https://doi.org/10.3390/v11050478