A Global Perspective on H9N2 Avian Influenza Virus




Abstract
:1. Introduction
2. History and Phylogeography of H9N2 Virus in Poultry
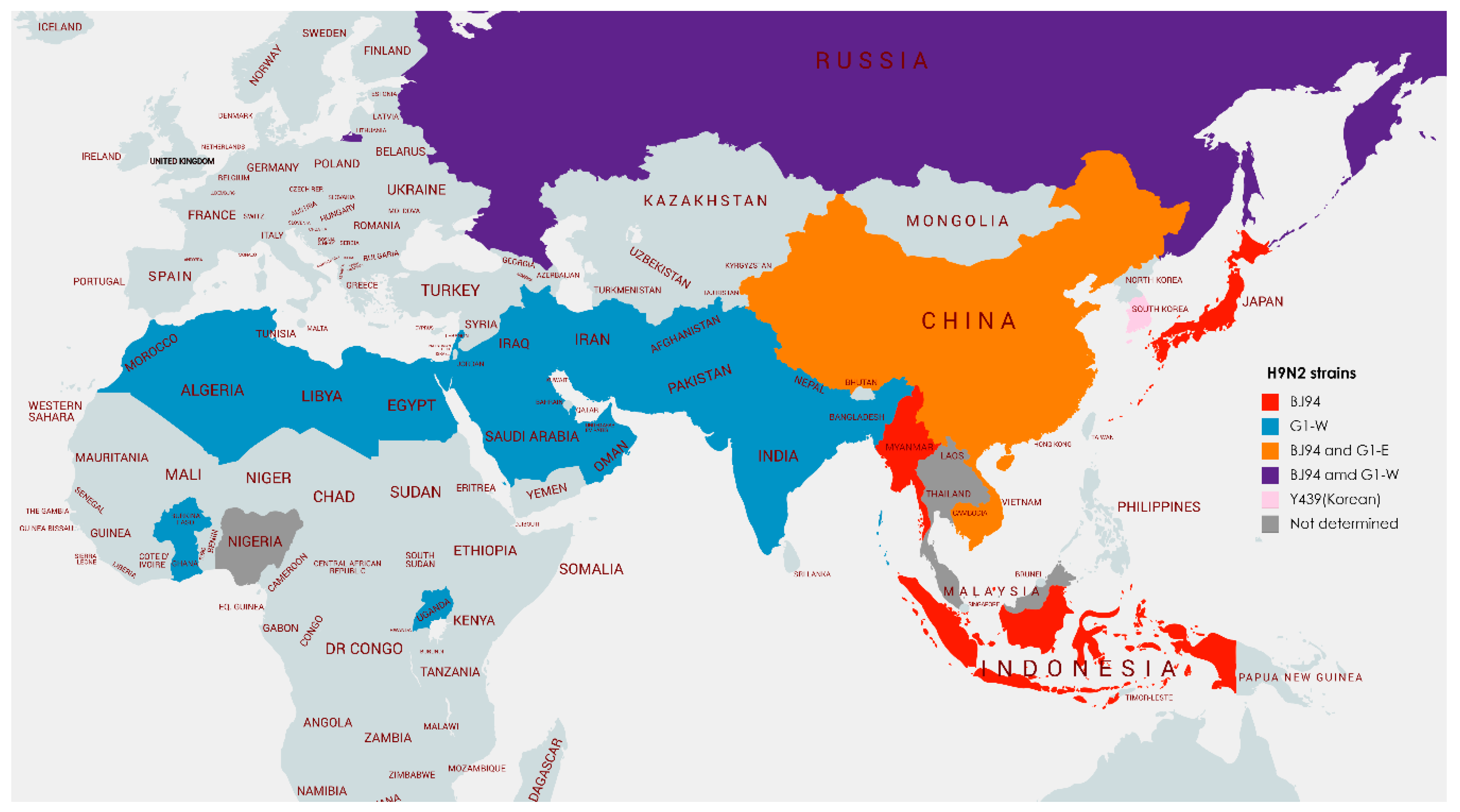
2.1. Phylogeography of H9N2 Viruses
2.1.1. East and Southeast Asia
2.1.2. South Asia
2.1.3. The Middle East
2.1.4. Africa
2.1.5. Europe
2.1.6. The Americas
2.2. Hyper-Prevalence of H9N2 Viruses in Poultry
3. H9N2 Reassortment and Evolution
3.1. H9N2 Virus Pathogenesis
3.2. H9N2 Virus Transmission and Host Tropism in Poultry
4. H9N2 Reassortment and Evolution
4.1. H9N2 Viruses as Gene Donors
4.2. H9N2 Viruses asGene Recipients
4.3. H9N2 Intrasubtypic Reassortment
5. H9N2 Virus in Humans
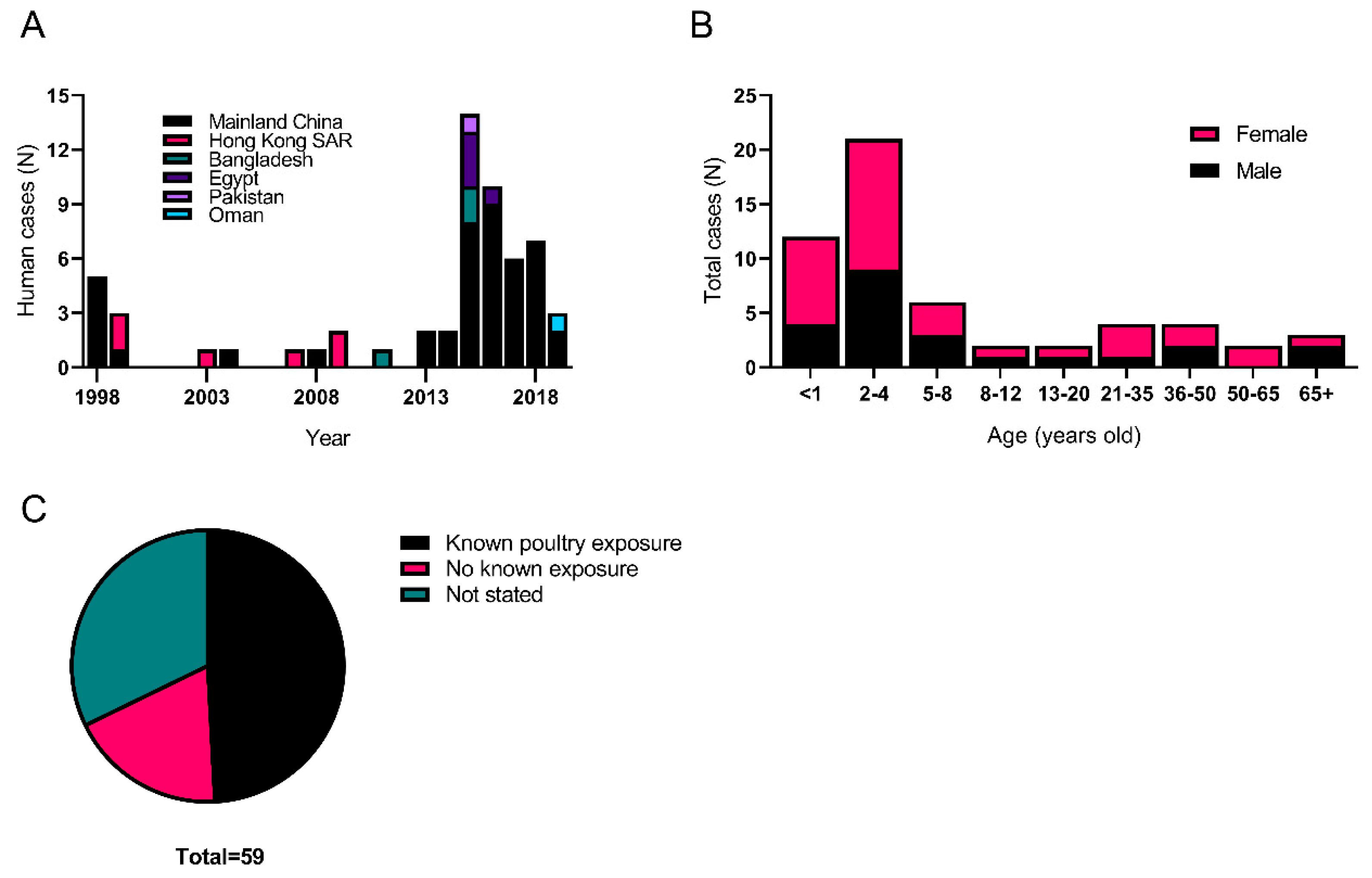
5.1. History of Human Infections with H9N2
5.2. Seropositivity Rates
5.3. Haemagglutinin and Receptor Binding
Molecular Basis of Receptor Binding
5.4. Ferret Experiments
5.5. Other Factors Involved in Zoonotic and Pandemic Potential in H9N2 Viruses
6. H9N2 Infection in Other Species
6.1. Minor Poultry Species
6.2. Swine
6.3. Canids
6.4. Horses
6.5. Mustelidae
6.6. Lagomorpha
6.7. Chiroptera
7. Vaccination and Control
8. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Wu, Y.; Tefsen, B.; Shi, Y.; Gao, G.F. Bat-derived influenza-like viruses H17N10 and H18N11. Trends Microbiol. 2014, 22, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Berhane, Y.; Hisanaga, T.; Kehler, H.; Neufeld, J.; Manning, L.; Argue, C.; Handel, K.; Hooper-McGrevy, K.; Jonas, M.; Robinson, J.; et al. Highly pathogenic avian influenza virus a (H7N3) in domestic poultry, saskatchewan, Canada, 2007. Emerg. Infect. Dis. 2009, 15, 1492–1495. [Google Scholar] [CrossRef]
- Capua, I.; Marangon, S. The avian influenza epidemic in Italy, 1999–2000: A review. Avian Pathol. 2000, 29, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Fouchier, R.A.; Schneeberger, P.M.; Rozendaal, F.W.; Broekman, J.M.; Kemink, S.A.; Munster, V.; Kuiken, T.; Rimmelzwaan, G.F.; Schutten, M.; Van Doornum, G.J.; et al. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc. Natl. Acad. Sci. USA 2004, 101, 1356–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.J.; Krauss, S.; Senne, D.A.; Mo, I.P.; Lo, K.S.; Xiong, X.P.; Norwood, M.; Shortridge, K.F.; Webster, R.G.; Guan, Y. Characterization of the pathogenicity of members of the newly established H9N2 influenza virus lineages in Asia. Virology 2000, 267, 279–288. [Google Scholar] [CrossRef]
- Suarez, D.L.; Perdue, M.L.; Cox, N.; Rowe, T.; Bender, C.; Huang, J.; Swayne, D.E. Comparisons of highly virulent H5N1 influenza A viruses isolated from humans and chickens from Hong Kong. J. Virol. 1998, 72, 6678–6688. [Google Scholar] [PubMed]
- Lam, T.T.; Wang, J.; Shen, Y.; Zhou, B.; Duan, L.; Cheung, C.L.; Ma, C.; Lycett, S.J.; Leung, C.Y.; Chen, X.; et al. The genesis and source of the H7n9 influenza viruses causing human infections in China. Nature 2013, 502, 241–244. [Google Scholar] [CrossRef]
- Homme, P.J.; Easterday, B.C. Avian influenza virus infections. I. Characteristics of influenza A-turkey-Wisconsin-1966 virus. Avian Dis. 1970, 14, 66–74. [Google Scholar] [CrossRef]
- Shortridge, K.F. Pandemic influenza: A zoonosis? Semin Respir Infect 1992, 7, 11–25. [Google Scholar]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef]
- Khalenkov, A.; Perk, S.; Panshin, A.; Golender, N.; Webster, R.G. Modulation of the severity of highly pathogenic H5N1 influenza in chickens previously inoculated with Israeli H9N2 influenza viruses. Virology 2009, 383, 32–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naguib, M.M.; Grund, C.; Arafa, A.S.; Abdelwhab, E.M.; Beer, M.; Harder, T.C. Heterologous post-infection immunity against egyptian avian influenza virus (AIV) H9N2 modulates the course of subsequent infection by highly pathogenic AIV H5N1, but vaccination immunity does not. J. Gen. Virol. 2017, 98, 1169–1173. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Luo, J.; Zhang, H.; Wang, C.; Duan, M.; Deliberto, T.J.; Nolte, D.L.; Ji, G.; He, H. Phylogenetic diversity and genotypical complexity of H9N2 influenza A viruses revealed by genomic sequence analysis. PLoS ONE 2011, 6, e17212. [Google Scholar] [CrossRef]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A brief summary of the epidemiology and genetic relatedness of avian influenza H9N2 virus in birds and mammals in the middle east and North Africa. Epidemiol Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, A.; Monne, I.; Salviato, A.; Valastro, V.; Schivo, A.; Amarin, N.M.; Gonzalez, C.; Ismail, M.M.; Al-Ankari, A.R.; Al-Blowi, M.H.; et al. Phylogeography and evolutionary history of reassortant H9N2 viruses with potential human health implications. J. Virol. 2011, 85, 8413–8421. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2015, 112, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Byarugaba, D.K.; Erima, B.; Ukuli, Q.A.; Atim, A.; Tugume, T.; Millard, M.; Kibuuka, K.; Mimbe, M.; Mworozi, E.A.; Danner, A.; et al. Hemagglutinin [Influenza A Virus]. Accession no. Avk87156.1. GenBank. 2018. Available online: https://www.ncbi.nlm.nih.gov/protein/Avk87156.1 (accessed on 3 July 2019).
- Kim, K.I.; Choi, J.G.; Kang, H.M.; To, T.L.; Nguyen, T.D.; Song, B.M.; Hong, M.S.; Choi, K.S.; Kye, S.J.; Kim, J.Y.; et al. Geographical distribution of low pathogenic avian influenza viruses of domestic poultry in Vietnam and their genetic relevance with Asian isolates. Poult. Sci. 2013, 92, 2012–2023. [Google Scholar] [CrossRef]
- Kim, J.A.; Cho, S.H.; Kim, H.S.; Seo, S.H. H9N2 influenza viruses isolated from poultry in Korean live bird markets continuously evolve and cause the severe clinical signs in layers. Vet. Microbiol. 2006, 118, 169–176. [Google Scholar] [CrossRef]
- Alexander, D.J. Report on avian influenza in the eastern hemisphere during 1997–2002. Avian Dis. 2003, 47, 792–797. [Google Scholar] [CrossRef]
- Horm, S.V.; Tarantola, A.; Rith, S.; Ly, S.; Gambaretti, J.; Duong, V.; Y, P.; Sorn, S.; Holl, D.; Allal, L.; et al. Intense circulation of A/H5N1 and other avian influenza viruses in cambodian live-bird markets with serological evidence of sub-clinical human infections. Emerg. Microbes. Infect. 2016, 5, e70. [Google Scholar] [CrossRef]
- Lin, T.N.; Nonthabenjawan, N.; Chaiyawong, S.; Bunpapong, N.; Boonyapisitsopa, S.; Janetanakit, T.; Mon, P.P.; Mon, H.H.; Oo, K.N.; Oo, S.M.; et al. Influenza A(H9N2) virus, Myanmar, 2014–2015. Emerg. Infect. Dis. 2017, 23, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Sonnberg, S.; Phommachanh, P.; Naipospos, T.S.; McKenzie, J.; Chanthavisouk, C.; Pathammavong, S.; Darnell, D.; Meeduangchanh, P.; Rubrum, A.M.; Souriya, M.; et al. Multiple introductions of avian influenza viruses (H5N1), Laos, 2009–2010. Emerg. Infect. Dis. 2012, 18, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Krueger, W.S.; Khuntirat, B.; Yoon, I.K.; Blair, P.J.; Chittagarnpitch, M.; Putnam, S.D.; Supawat, K.; Gibbons, R.V.; Bhuddari, D.; Pattamadilok, S.; et al. Prospective study of avian influenza virus infections among rural thai villagers. PLoS ONE 2013, 8, e72196. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.; Sahesti, A.; Murwijati, T.; Lestariningsih, C.L.; Irine, I.; Ayesda, C.S.; Prihartini, W.; Mahardika, G.N. Identification of avian influenza virus subtype H9N2 in chicken farms in Indonesia. Prev. Vet. Med. 2018, 159, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.R. Should we be concerned about the H9N2 virus? New Straits Times, 19 December 2018; 15. [Google Scholar]
- Marchenko, V.Y.; Goncharova, N.I.; Evseenko, V.A.; Susloparov, I.M.; Gavrilova, E.V.; Maksyutov, R.A.; Ryzhikov, A.B. Overview of the epidemiological situation on highly pathogenic avian influenza virus in Russia in 2018. Problemy Osobo Opasnykh Infektsii [Problems of Particularly Dangerous Infections] 2019, 1, 42–49. [Google Scholar] [CrossRef]
- Karlsson, E.A.; Horm, S.V.; Tok, S.; Tum, S.; Kalpravidh, W.; Claes, F.; Osbjer, K.; Dussart, P. Avian influenza virus detection, temporality and co-infection in poultry in Cambodian border provinces, 2017–2018. Emerg. Microbes. Infect. 2019, 8, 637–639. [Google Scholar] [CrossRef] [PubMed]
- Park, K.J.; Kwon, H.I.; Song, M.S.; Pascua, P.N.; Baek, Y.H.; Lee, J.H.; Jang, H.L.; Lim, J.Y.; Mo, I.P.; Moon, H.J.; et al. Rapid evolution of low-pathogenic H9N2 avian influenza viruses following poultry vaccination programmes. J. Gen. Virol. 2011, 92, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Tang, Y.; Liu, X.; Peng, D.; Liu, W.; Liu, H.; Lu, S.; Liu, X. Characterization of H9N2 influenza viruses isolated from vaccinated flocks in an integrated broiler chicken operation in eastern China during a 5 year period (1998–2002). J. Gen. Virol. 2008, 89, 3102–3112. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; McCauley, J. Gisaid: Global initiative on sharing all influenza data-from vision to reality. Euro. Surveill. 2017, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Bolotov, P.; Dernovoy, D.; Kiryutin, B.; Zaslavsky, L.; Tatusova, T.; Ostell, J.; Lipman, D. The influenza virus resource at the national center for biotechnology information. J. Virol. 2008, 82, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; et al. Influenza research database: An integrated bioinformatics resource for influenza virus research. Nucleic. Acids Res. 2017, 45, D466–D474. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.R.; Gregory, V.; Banks, J.; Brown, I.H.; Alexander, D.J.; Hay, A.J.; Lin, Y.P. H9N2 subtype influenza A viruses in poultry in pakistan are closely related to the H9N2 viruses responsible for human infection in Hong Kong. Virology 2000, 278, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Negovetich, N.J.; Feeroz, M.M.; Jones-Engel, L.; Walker, D.; Alam, S.M.; Hasan, K.; Seiler, P.; Ferguson, A.; Friedman, K.; Barman, S.; et al. Live bird markets of Bangladesh: H9N2 viruses and the near absence of highly pathogenic H5N1 influenza. PLoS ONE 2011, 6, e19311. [Google Scholar] [CrossRef] [PubMed]
- Tosh, C.; Nagarajan, S.; Behera, P.; Rajukumar, K.; Purohit, K.; Kamal, R.P.; Murugkar, H.V.; Gounalan, S.; Pattnaik, B.; Vanamayya, P.R.; et al. Genetic analysis of H9N2 avian influenza viruses isolated from India. Arch. Virol. 2008, 153, 1433–1439. [Google Scholar] [CrossRef]
- Hosseini, H.; Ghalyanchilangeroudi, A.; Fallah Mehrabadi, M.H.; Sediqian, M.S.; Shayeganmehr, A.; Ghafouri, S.A.; Maghsoudloo, H.; Abdollahi, H.; Farahani, R.K. Phylogenetic analysis of H9N2 avian influenza viruses in Afghanistan (2016–2017). Arch. Virol. 2017, 162, 3161–3165. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J. Summary of avian influenza Activity in Europe, Asia, Africa, and Australasia, 2002–2006. Avian Dis. 2007, 51, 161–166. [Google Scholar] [CrossRef]
- Brown, I.H. Summary of avian influenza Activity in Europe, Asia, and Africa, 2006–2009. Avian Dis. 2010, 54, 187–193. [Google Scholar] [CrossRef]
- Iqbal, M.; Yaqub, T.; Reddy, K.; McCauley, J.W. Novel genotypes of H9N2 influenza A viruses isolated from poultry in Pakistan containing ns genes similar to highly pathogenic H7N3 and H5N1 viruses. PLoS ONE 2009, 4, e5788. [Google Scholar] [CrossRef]
- Parvin, R.; Heenemann, K.; Halami, M.Y.; Chowdhury, E.H.; Islam, M.R.; Vahlenkamp, T.W. Full-genome analysis of avian influenza virus H9N2 from Bangladesh reveals internal gene reassortments with two distinct highly pathogenic avian influenza viruses. Arch. Virol. 2014, 159, 1651–1661. [Google Scholar] [CrossRef]
- Monne, I.; Hussein, H.A.; Fusaro, A.; Valastro, V.; Hamoud, M.M.; Khalefa, R.A.; Dardir, S.N.; Radwan, M.I.; Capua, I.; Cattoli, G. H9N2 influenza A virus circulates in H5N1 endemically infected poultry population in Egypt. Influenza Other Respi. Viruses 2013, 7, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Aamir, U.B.; Wernery, U.; Ilyushina, N.; Webster, R.G. Characterization of avian H9N2 influenza viruses from United Arab Emirates 2000 to 2003. Virology 2007, 361, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banet-Noach, C.; Perk, S.; Simanov, L.; Grebenyuk, N.; Rozenblut, E.; Pokamunski, S.; Pirak, M.; Tendler, Y.; Panshin, A. H9N2 influenza viruses from Israeli poultry: A five-year outbreak. Avian Dis. 2007, 51, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Body, M.H.; Alrarawahi, A.H.; Alhubsy, S.S.; Saravanan, N.; Rajmony, S.; Mansoor, M.K. Characterization of low pathogenic avian influenza virus subtype H9N2 isolated from free-living mynah birds (acridotheres tristis) in the sultanate of Oman. Avian Dis. 2015, 59, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Barbour, E.K.; Sagherian, V.K.; Sagherian, N.K.; Dankar, S.K.; Jaber, L.S.; Usayran, N.N.; Farran, M.T. Avian influenza outbreak in poultry in the Lebanon and transmission to neighbouring farmers and swine. Vet. Ital. 2006, 42, 77–85. [Google Scholar] [PubMed]
- Monne, I.; Cattoli, G.; Mazzacan, E.; Amarin, N.M.; Al Maaitah, H.M.; Al-Natour, M.Q.; Capua, I. Genetic comparison of H9N2 AI viruses isolated in Jordan in 2003. Avian Dis. 2007, 51, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Kraidi, Q.A.; Madadgar, O.; Ghalyanchi Langeroudi, A.; Karimi, V. Genetic analysis of H9N2 avian influenza viruses circulated in broiler flocks: A case study in Iraq in 2014–2015. Virus Genes 2017, 53, 205–214. [Google Scholar] [CrossRef]
- Davidson, I.; Shkoda, I.; Golender, N.; Perk, S.; Lapin, K.; Khinich, Y.; Panshin, A. Genetic characterization of ha gene of low pathogenic H9N2 influenza viruses isolated in israel during 2006–2012 periods. Virus Genes 2013, 46, 255–263. [Google Scholar] [CrossRef]
- Tombari, W.; Nsiri, J.; Larbi, I.; Guerin, J.L.; Ghram, A. Genetic evolution of low pathogenecity H9N2 avian influenza viruses in Tunisia: Acquisition of new mutations. Virol. J. 2011, 8, 467. [Google Scholar] [CrossRef]
- Kammon, A.; Heidari, A.; Dayhum, A.; Eldaghayes, I.; Sharif, M.; Monne, I.; Cattoli, G.; Asheg, A.; Farhat, M.; Kraim, E. Characterization of avian influenza And newcastle disease viruses from poultry in Libya. Avian Dis. 2015, 59, 422–430. [Google Scholar] [CrossRef]
- El Houadfi, M.; Fellahi, S.; Nassik, S.; Guerin, J.L.; Ducatez, M.F. First outbreaks and phylogenetic analyses of avian influenza H9N2 viruses isolated from poultry flocks in Morocco. Virol. J. 2016, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- Zecchin, B.; Minoungou, G.; Fusaro, A.; Moctar, S.; Ouedraogo-Kabore, A.; Schivo, A.; Salviato, A.; Marciano, S.; Monne, I. Influenza A(H9N2) virus, Burkina Faso. Emerg. Infect. Dis. 2017, 23, 2118–2119. [Google Scholar] [CrossRef] [PubMed]
- Rubrum, A.; Jeevan, T.; Darnell, D.; Webby, R.; Derrar, F.; Gradi, E.-A. Hemagglutinin [influenza A virus]. Accession no. Azf86190.1. GenBank. 2018. Available online: https://www.ncbi.nlm.nih.gov/protein/Azf86190.1 (accessed on 3 July 2019).
- Awuni, J.A.; Bianco, A.; Dogbey, O.J.; Fusaro, A.; Yingar, D.T.; Salviato, A.; Ababio, P.T.; Milani, A.; Bonfante, F.; Monne, I. Avian influenza H9N2 subtype in Ghana: Virus characterization and evidence of co-infection. Avian Pathol. 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Abolnik, C.; Cornelius, E.; Bisschop, S.P.; Romito, M.; Verwoerd, D. Phylogenetic analyses of genes from South African LPAI viruses isolated in 2004 from wild aquatic birds suggests introduction by Eurasian migrants. Dev. Biol. 2006, 124, 189–199. [Google Scholar] [PubMed]
- Oluwayelu, D.O.; Omolanwa, A.; Adebiyi, A.I.; Aiki-Raji, C.O. Flock-based surveillance for low pathogenic avian influenza virus in commercial breeders and layers, southwest nigeria. Afr. J. Infect. Dis. 2016, 11, 44–49. [Google Scholar] [CrossRef]
- Okoye, J.; Eze, D.; Krueger, W.S.; Heil, G.L.; Friary, J.A.; Gray, G.C. Serologic evidence of avian influenza virus infections among Nigerian agricultural workers. J. Med. Virol. 2013, 85, 670–676. [Google Scholar] [CrossRef]
- Reid, S.M.; Banks, J.; Ceeraz, V.; Seekings, A.; Howard, W.A.; Puranik, A.; Collins, S.; Manvell, R.; Irvine, R.M.; Brown, I.H. The detection of a low pathogenicity avian influenza virus subtype H9 infection in a turkey breeder flock in the United Kingdom. Avian Dis. 2016, 60, 126–131. [Google Scholar] [CrossRef]
- Swieton, E.; Jozwiak, M.; Minta, Z.; Smietanka, K. Genetic characterization of H9N2 avian influenza viruses isolated from poultry in Poland during 2013/2014. Virus Genes 2018, 54, 67–76. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Lexmond, P.; Vuong, O.; Schutten, M.; Guldemeester, J.; Osterhaus, A.D.; Elbers, A.R.; Slaterus, R.; Hornman, M.; Koch, G.; et al. Discordant detection of avian influenza virus subtypes in time and space between poultry and wild birds; towards improvement of surveillance programs. PLoS ONE 2017, 12, e0173470. [Google Scholar] [CrossRef]
- Harder, T. Endemic non-notifiable avian influenza virus infections in poultry, Joint 24th Annual Meetings of the National Laboratories for Avian Influenza And Newcastle Disease of European Union Member States 2018; European Commission. Available online: https://ec.europa.eu/food/sites/food/files/animals/docs/ad_cm_20180710_24-annual-meetings_ai-pres-08.pdf (accessed on 3 July 2019).
- Coman, A.; Maftei, D.N.; Krueger, W.S.; Heil, G.L.; Friary, J.A.; Chereches, R.M.; Sirlincan, E.; Bria, P.; Dragnea, C.; Kasler, I.; et al. Serological evidence for avian H9N2 influenza virus infections among Romanian agriculture workers. J. Infect. Public Health 2013, 6, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Panigrahy, B.; Senne, D.A.; Pedersen, J.C. Avian influenza virus subtypes inside and outside the live bird markets, 1993–2000: A spatial and temporal relationship. Avian Dis. 2002, 46, 298–307. [Google Scholar] [CrossRef]
- Senne, D.A. Avian influenza in north and South America, the Caribbean, and Australia, 2006–2008. Avian Dis. 2010, 54, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A. Avian influenza in north and South America, 2002–2005. Avian Dis. 2007, 51, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A. Avian influenza in the western hemisphere including the pacific islands and Australia. Avian Dis. 2003, 47, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Pasick, J.; Pedersen, J.; Hernandez, M.S. Avian influenza in north America, 2009–2011. Avian Dis. 2012, 56, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.F.; Dong, L.; Lan, Y.; Long, L.P.; Xu, C.; Zou, S.; Li, Z.; Wen, L.; Cai, Z.; Wang, W.; et al. Indications that live poultry markets are a major source of human H5N1 influenza virus infection in China. J. Virol. 2011, 85, 13432–13438. [Google Scholar] [CrossRef] [PubMed]
- Fournie, G.; Guitian, J.; Desvaux, S.; Cuong, V.C.; Dung do, H.; Pfeiffer, D.U.; Mangtani, P.; Ghani, A.C. Interventions for avian influenza A (H5N1) risk management in live bird market networks. Proc. Natl. Acad. Sci. USA 2013, 110, 9177–9182. [Google Scholar] [CrossRef] [Green Version]
- Thuy, D.M.; Peacock, T.P.; Bich, V.T.; Fabrizio, T.; Hoang, D.N.; Tho, N.D.; Diep, N.T.; Nguyen, M.; Hoa, L.N.; Trang, H.T.; et al. Prevalence and diversity of H9N2 avian influenza in chickens of northern Vietnam, 2014. Infect. Genet. Evol. 2016, 44, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.H.; Okamatsu, M.; Matsuno, K.; Hiono, T.; Ogasawara, K.; Nguyen, L.T.; Van Nguyen, L.; Nguyen, T.N.; Nguyen, T.T.; Van Pham, D.; et al. Genetic and antigenic characterization of H5, H6 and H9 avian influenza viruses circulating in live bird markets with intervention in the center part of Vietnam. Vet. Microbiol. 2016, 192, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.J.; Lin, X.D.; Guo, W.P.; Tian, J.H.; Wang, W.; Ying, X.H.; Wang, M.R.; Yu, B.; Yang, Z.Q.; Shi, M.; et al. Diversity and evolution of avian influenza viruses in live poultry markets, free-range poultry and wild wetland birds in China. J. Gen. Virol. 2016, 97, 844–854. [Google Scholar] [CrossRef]
- Huang, Y.; Li, X.; Zhang, H.; Chen, B.; Jiang, Y.; Yang, L.; Zhu, W.; Hu, S.; Zhou, S.; Tang, Y.; et al. Human infection with an avian influenza A (H9N2) virus in the middle region of China. J. Med. Virol. 2015, 87, 1641–1648. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhao, B.; Li, Y.; Zhang, X.; Chen, S.; Chen, T. Clinical and epidemiological characteristics of a young child infected with avian influenza A (H9N2) virus in China. J. Int. Med. Res. 2018, 46, 3462–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Chin, P.S.; Dyrting, K.C.; Ellis, T.M.; Webster, R.G.; Peiris, M. H9N2 influenza viruses possessing H5N1-like internal genomes continue to circulate in poultry in southeastern China. J. Virol. 2000, 74, 9372–9380. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.C.; Feeroz, M.M.; Hasan, M.K.; Akhtar, S.; Walker, D.; Seiler, P.; Barman, S.; Franks, J.; Jones-Engel, L.; McKenzie, P.; et al. Insight into live bird markets of Bangladesh: An overview of the dynamics of transmission of H5N1 and H9N2 avian influenza viruses. Emerg. Microbes Infect. 2017, 6, e12. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.; Ahmad, M.; Rashid, H.B.; Sultan, B.; Chaudhry, H.R.; Riaz, A.; Shaheen, M.S. Prospective study of avian influenza H9 infection in commercial poultry farms of punjab province and islamabad capital territory, Pakistan. Trop. Anim. Health Prod. 2017, 49, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.; Rashid, H.B.; Angot, A.; Thrusfield, M.; Bronsvoort, B.M.D.; Capua, I.; Cattoli, G.; Welburn, S.C.; Eisler, M.C. Risk factors for avian influenza H9 infection of chickens in live bird retail stalls of lahore district, Pakistan 2009–2010. Sci. Rep. 2018, 8, 5634. [Google Scholar] [CrossRef] [PubMed]
- Nili, H.; Asasi, K. Avian influenza (H9N2) outbreak in Iran. Avian Dis. 2003, 47, 828–831. [Google Scholar] [CrossRef]
- Kishida, N.; Sakoda, Y.; Eto, M.; Sunaga, Y.; Kida, H. Co-infection of Staphylococcus aureus or Haemophilus paragallinarum exacerbates H9N2 influenza A virus infection in chickens. Arch. Virol. 2004, 149, 2095–2104. [Google Scholar] [CrossRef]
- Pan, Q.; Liu, A.; Zhang, F.; Ling, Y.; Ou, C.; Hou, N.; He, C. Co-infection of broilers with Ornithobacterium rhinotracheale and H9N2 avian influenza virus. BMC. Vet. Res. 2012, 8, 104. [Google Scholar] [CrossRef]
- Seifi, S.; Asasi, K.; Mohammadi, A. Natural co-infection caused by avian influenza H9 subtype and infectious bronchitis viruses in broiler chicken farms. Veterinarski Arch. 2010, 80, 269–281. [Google Scholar]
- James, J.; Howard, W.; Iqbal, M.; Nair, V.; Barclay, W.S.; Shelton, H. Influenza A virus PB1-F2 protein prolongs viral shedding in chickens lengthening the transmission window. J. Gen. Virol. 2016, 97, 2516–2527. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, Z.; Guo, X.; Zhang, Y.; Wang, D.; Xu, S.; Yin, Y. Cytokine expression in 3 chicken host systems infected with H9N2 influenza viruses with different pathogenicities. Avian Pathol. 2016, 45, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, B.; Chen, Q.; Chen, Z. Characterization of H9N2 influenza viruses isolated from Dongting Lake wetland in 2007. Arch. Virol. 2011, 156, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Lee, D.H.; Park, J.K.; Lim, T.H.; Youn, H.N.; Yuk, S.S.; Lee, Y.J.; Mo, I.P.; Sung, H.W.; Lee, J.B.; et al. Isolation and characterization of a novel H9N2 influenza virus in Korean native chicken farm. Avian Dis. 2011, 55, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Gohrbandt, S.; Veits, J.; Breithaupt, A.; Hundt, J.; Teifke, J.P.; Stech, O.; Mettenleiter, T.C.; Stech, J. H9 avian influenza reassortant with engineered polybasic cleavage site displays a highly pathogenic phenotype in chicken. J. Gen. Virol. 2011, 92, 1843–1853. [Google Scholar] [CrossRef] [PubMed]
- Killingley, B.; Nguyen-Van-Tam, J. Routes of influenza transmission. Influenza Other Respir. Viruses 2013, 7, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Shortridge, K.F.; Zhou, N.N.; Guan, Y.; Gao, P.; Ito, T.; Kawaoka, Y.; Kodihalli, S.; Krauss, S.; Markwell, D.; Murti, K.G.; et al. Characterization of avian H5N1 influenza viruses from poultry in Hong Kong. Virology 1998, 252, 331–342. [Google Scholar] [CrossRef]
- Spekreijse, D.; Bouma, A.; Koch, G.; Stegeman, J.A. Airborne transmission of a highly pathogenic avian influenza virus strain H5N1 between groups of chickens quantified in an experimental setting. Vet. Microbiol. 2011, 152, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wu, J.; Zeng, X.; Huang, G.; Zou, L.; Song, Y.; Gopinath, D.; Zhang, X.; Kang, M.; Lin, J.; et al. Isolation of H5N6, H7N9 and H9N2 avian influenza A viruses from air sampled at live poultry markets in China, 2014 and 2015. Euro. Surveill. 2016, 21, 35. [Google Scholar] [CrossRef]
- Claes, G.; Welby, S.; Van Den Berg, T.; Van Der Stede, Y.; Dewulf, J.; Lambrecht, B.; Marche, S. The impact of viral tropism and housing conditions on the transmission of three H5/H7 low pathogenic avian influenza viruses in chickens. Epidemiol Infect. 2013, 141, 2428–2443. [Google Scholar] [CrossRef]
- Banks, J.; Speidel, E.S.; Moore, E.; Plowright, L.; Piccirillo, A.; Capua, I.; Cordioli, P.; Fioretti, A.; Alexander, D.J. Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Arch. Virol. 2001, 146, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, E.M.; Song, H.; Pena, L.; Perez, D.R. A 27-amino-acid deletion in the neuraminidase stalk supports replication of an avian H2N2 influenza A virus in the respiratory tract of chickens. J. Virol. 2010, 84, 11831–11840. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wei, L.; Yang, Y.; Wang, B.; Liang, W.; Gao, Y.; Xia, X.; Gao, L.; Cai, Y.; Hou, P.; et al. Amino acid substitutions in the neuraminidase protein of an H9N2 avian influenza virus affect its airborne transmission in chickens. Vet. Res. 2015, 46, 44. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, F.; Mazzetto, E.; Zanardello, C.; Fortin, A.; Gobbo, F.; Maniero, S.; Bigolaro, M.; Davidson, I.; Haddas, R.; Cattoli, G.; et al. A G1-lineage H9N2 virus with oviduct tropism causes chronic pathological changes in the infundibulum and a long-lasting drop in egg production. Vet. Res. 2018, 49, 83. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.G.; Castro, A.E. Evaluation of the infectivity, length of infection, and immune response of a low-pathogenicity H7N2 avian influenza virus in specific-pathogen-free chickens. Avian Dis. 2004, 48, 263–270. [Google Scholar] [CrossRef]
- van der Goot, J.A.; de Jong, M.C.; Koch, G.; Van Boven, M. Comparison of the transmission characteristics of low and high pathogenicity avian influenza A virus (H5N2). Epidemiol. Infect. 2003, 131, 1003–1013. [Google Scholar] [CrossRef]
- Perez, D.R.; Lim, W.; Seiler, J.P.; Yi, G.; Peiris, M.; Shortridge, K.F.; Webster, R.G. Role of quail in the interspecies transmission of H9 influenza A viruses: Molecular changes on ha that correspond to adaptation from ducks to chickens. J. Virol. 2003, 77, 3148–3156. [Google Scholar] [CrossRef]
- Yao, M.; Lv, J.; Huang, R.; Yang, Y.; Chai, T. Determination of infective dose of H9N2 avian influenza virus in different routes: Aerosol, intranasal, and gastrointestinal. Intervirology 2014, 57, 369–374. [Google Scholar] [CrossRef]
- Seiler, P.; Kercher, L.; Feeroz, M.M.; Shanmuganatham, K.; Jones-Engel, L.; Turner, J.; Walker, D.; Alam, S.M.R.; Hasan, M.K.; Akhtar, S.; et al. H9N2 influenza viruses from Bangladesh: Transmission in chicken and new world quail. Influenza Other Respir. Viruses 2018, 12, 814–817. [Google Scholar] [CrossRef]
- Peacock, T.P.; Benton, D.J.; James, J.; Sadeyen, J.R.; Chang, P.; Sealy, J.E.; Bryant, J.E.; Martin, S.R.; Shelton, H.; Barclay, W.S.; et al. Immune escape variants of H9N2 influenza viruses containing deletions at the haemagglutinin receptor binding site retain fitness in vivo and display enhanced zoonotic characteristics. J. Virol. 2017, 91, e00218-17. [Google Scholar] [CrossRef]
- Liu, D.; Shi, W.; Gao, G.F. Poultry carrying H9N2 act as incubators for novel human avian influenza viruses. Lancet 2014, 383, 869–875. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; et al. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: A descriptive study. Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef]
- Tosh, C.; Nagarajan, S.; Kumar, M.; Murugkar, H.V.; Venkatesh, G.; Shukla, S.; Mishra, A.; Mishra, P.; Agarwal, S.; Singh, B.; et al. Multiple introductions of a reassortant H5N1 avian influenza virus of clade 2.3.2.1c with PB2 gene of H9N2 subtype into Indian poultry. Infect. Genet. Evol. 2016, 43, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Meng, F.; Huang, D.; Sheng, X.; Wang, Y.; Zhang, W.; Chang, W.; Wang, L.; Qin, Z. Genomic and phylogenetic characterization of novel, recombinant H5N2 avian influenza virus strains isolated from vaccinated chickens with clinical symptoms in China. Viruses 2015, 7, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.Y.; Ke, C.W.; Li, Q.; Yuan, R.Y.; Xiang, D.; Jia, W.X.; Yu, Y.D.; Liu, L.; Huang, C.; Qi, W.B.; et al. Novel reassortant avian influenza A(H5N6) viruses in humans, Guangdong, China, 2015. Emerg. Infect. Dis. 2016, 22, 1507. [Google Scholar] [CrossRef] [PubMed]
- Monne, I.; Meseko, C.; Joannis, T.; Shittu, I.; Ahmed, M.; Tassoni, L.; Fusaro, A.; Cattoli, G. Highly pathogenic avian influenza A(H5N1) virus in poultry, Nigeria, 2015. Emerg. Infect. Dis. 2015, 21, 1275–1277. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xie, Q.; Zhang, S.; Li, Y.; Xiao, H.; Jin, T.; Zheng, W.; Li, J.; Jia, X.; Sun, L.; et al. Assessment of the internal genes of influenza A (H7N9) virus contributing to the high pathogenicity in mice. J. Virol. 2014, 89, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Shanmuganatham, K.; Feeroz, M.M.; Jones-Engel, L.; Walker, D.; Alam, S.; Hasan, M.; McKenzie, P.; Krauss, S.; Webby, R.J.; Webster, R.G. Genesis of avian influenza H9N2 in Bangladesh. Emerg. Microbes Infect. 2014, 3, e88. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Xu, C.; Wang, C.; Wu, B.; Luo, J.; Zhang, H.; Nolte, D.L.; Deliberto, T.J.; Duan, M.; Ji, G.; et al. Reassortant H9N2 influenza viruses containing H5N1-like PB1 genes isolated from black-billed magpies in southern China. PLoS ONE 2011, 6, e25808. [Google Scholar] [CrossRef]
- Shanmuganatham, K.K.; Jones, J.C.; Marathe, B.M.; Feeroz, M.M.; Jones-Engel, L.; Walker, D.; Turner, J.; Rabiul Alam, S.M.; Kamrul Hasan, M.; Akhtar, S.; et al. The replication of Bangladeshi H9N2 avian influenza viruses carrying genes from H7N3 in mammals. Emerg. Microbes Infect. 2016, 5, e35. [Google Scholar] [CrossRef]
- Naguib, M.M.; Ulrich, R.; Kasbohm, E.; Eng, C.L.P.; Hoffmann, D.; Grund, C.; Beer, M.; Harder, T.C. Natural reassortants between potentially zoonotic avian influenza viruses H5N1 and H9N2 from Egypt display distinct pathogenic phenotypes in experimentally infected chickens and ferrets. J. Virol. 2017, 91, e01300-17. [Google Scholar] [CrossRef] [PubMed]
- Peiris, M.; Yuen, K.Y.; Leung, C.W.; Chan, K.H.; Ip, P.L.; Lai, R.W.; Orr, W.K.; Shortridge, K.F. Human infection with influenza H9N2. Lancet 1999, 354, 916–917. [Google Scholar] [CrossRef]
- Guo, Y.; Li, J.; Cheng, X. [Discovery of men infected by avian influenza A (H9N2) virus]. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 1999, 13, 105–108. [Google Scholar] [PubMed]
- International Centre for Diarrhoeal Disease Research (ICDDRB). Outbreak of mild respiratory disease caused by H5N1 and H9N2 infections among young children in Dhaka, Bangladesh; Health and Science Bulletin, ICDDRB: Dhaka, Bangladesh, 2011; Volume 9, pp. 5–12. [Google Scholar]
- Ali, M.; Yaqub, T.; Mukhtar, N.; Imran, M.; Ghafoor, A.; Shahid, M.F.; Naeem, M.; Iqbal, M.; Smith, G.J.D.; Su, Y.C.F. Avian influenza A(H9N2) virus in poultry worker, Pakistan, 2015. Emerg. Infect. Dis. 2019, 25, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Influenza At the human-animal interface, summary and assessment, 23 June 2015; World Health Organisation: Geneva, Switzerland, 2015; pp. 3–4.
- Influenza At the human-animal interface, summary and assessment, 10 April to 10 May 2019; World Health Organisation: Geneva, Switzerland, 2019; pp. 2–3.
- Influenza At the human-animal interface, summary and assessment, 20 July to 3 October 2016; World Health Organisation: Geneva, Switzerland, 2016; pp. 5–6.
- Li, Q.; Zhou, L.; Zhou, M.; Chen, Z.; Li, F.; Wu, H.; Xiang, N.; Chen, E.; Tang, F.; Wang, D.; et al. Epidemiology of human infections with avian influenza A(H7N9) virus in China. N. Engl. J. Med. 2014, 370, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Smallman-Raynor, M.; Cliff, A.D. Avian influenza A (H5N1) age distribution in humans. Emerg. Infect. Dis. 2007, 13, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, E.; Bao, C.; Xiang, N.; Wu, J.; Wu, S.; Shi, J.; Wang, X.; Zheng, Y.; Zhang, Y.; et al. Clusters of human infection and human-to-human transmission of avian influenza A(H7N9) virus, 2013–2017. Emerg. Infect. Dis. 2018, 24, 397. [Google Scholar] [CrossRef] [PubMed]
- Gou, Y.; Xie, J.; Wang, M. [A strain of influenza A H9N2 virus repeatedly isolated from human population in China]. Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi = Chinese journal of experimental and clinical virology 2000, 14, 209–212. [Google Scholar]
- Butt, K.M.; Smith, G.J.; Chen, H.; Zhang, L.J.; Leung, Y.H.; Xu, K.M.; Lim, W.; Webster, R.G.; Yuen, K.Y.; Peiris, J.S.; et al. Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. J. Clin. Microbiol. 2005, 43, 5760–5767. [Google Scholar] [CrossRef]
- Wang, K.; Chen, J.H. Influenza A virus (a/Guangdong/w1/2004(h9n2)) segment 4 hemagglutinin (ha) gene, complete cds, genbank: Kx867849.1. GenBank. 2016. Available online: https://www.ncbi.nlm.nih.gov/nuccore/Kx867849.1 (accessed on 3 July 2019).
- Hong Kong Department of Health. Gene sequencing of H9N2 virus shows avian origin. Hong Kong Department of Health. 2009. Available online: http://www.dh.gov.hk/english/press/2009/090107–2.html (accessed on 3 July 2019).
- Hong Kong Department of Health. CHP investigating case of influenza A (H9N2) infection. Hong Kong Department of Health. 2009. Available online: http://www.dh.gov.hk/english/press/2009/091223–3.html (accessed on 3 July 2019).
- Hong Kong Department of Health. CHP investigating a case of influenza A (H9) infection. Hong Kong Department of Health. 2009. Available online: http://www.dh.gov.hk/english/press/2009/091103–2.html (accessed on 3 July 2019).
- Influenza at the human-animal interface, Summary and assessment, 24 January 2014; World Health Organisation: Geneva, Switzerland, 2014; pp. 2–3.
- Xu, J.; Li, S.; Yang, Y.; Liu, B.; Yang, H.; Li, T.; Zhang, L.; Li, W.; Luo, X.; Zhang, L.; et al. Human infection with a further evolved avian H9N2 influenza A virus in Sichuan, China. Sci. China Life Sci. 2018, 61, 604–606. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, W.; Xie, Y.; Lin, J. Influenza A virus (a/zhongshan/201501/2015(h9n2)) segment 4 hemagglutinin (ha) gene, complete cds, genbank: Ku217316.1. GenBank. 2016. Available online: https://www.ncbi.nlm.nih.gov/nuccore/Ku217316.1 (accessed on 3 July 2019).
- Influenza at the human-animal interface, Summary and assessment, 4 September 2015; World Health Organisation: Geneva, Switzerland, 2015; p. 2.
- Influenza At the Human-Animal Interface, Summary and Assessment, 14 December 2015; World Health Organisation: Geneva, Switzerland, 2015; p. 3.
- He, J.; Liu, L.P.; Hou, S.; Gong, L.; Wu, J.B.; Hu, W.F.; Wang, J.J. genomic characteristics of 2 strains of influenza A(H9N2) virus isolated from human infection cases in Anhui province. Zhonghua Liu Xing Bing Xue Za Zhi 2016, 37, 708–713. [Google Scholar] [PubMed]
- Influenza at the Human-Animal Interface, Summary and Assessment, 20 January 2016; World Health Organisation: Geneva, Switzerland, 2016; p. 5.
- Jie, Y.; Zheng, H.; Xiaolei, L.; Zinhua, O.; Dong, Y.; Yingchun, S.; Lingzhi, L.; Rengui, Y. Full-length genome analysis of an avian influenza A virus (H9N2) from a human infection in Changsha city. Futur. Med. 2018, 13, 323–330. [Google Scholar] [CrossRef]
- Influenza at the Human-Animal Interface, Summary and Assessment, 21 January to 25 February 2016; World Health Organisation: Geneva, Switzerland, 2016; pp. 5–6.
- Influenza at the Human-Animal Interface, Summary and Assessment, 5 April to 9 May 2016; World Health Organisation: Geneva, Switzerland, 2016; pp. 5–6.
- Influenza At the Human-Animal Interface, Summary and Assessment, 13 June to 19 July 2016; World Health Organisation: Geneva, Switzerland, 2016; p. 5.
- Yuan, R.; Liang, L.; Wu, J.; Kang, Y.; Song, Y.; Zou, L.; Zhang, X.; Ni, H.; Ke, C. Human infection with an avian influenza A/H9N2 virus in Guangdong in 2016. J. Infect. 2017, 74, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Influenza at the Human-Animal Interface, Summary and Assessment, 20 December to 16 January 2017; World Health Organisation: Geneva, Switzerland, 2017; pp. 3–4.
- Pan, Y.; Cui, S.; Sun, Y.; Zhang, X.; Ma, C.; Shi, W.; Peng, X.; Lu, G.; Zhang, D.; Liu, Y.; et al. Human infection with H9N2 avian influenza in northern China. Clin. Microbiol. Infect. 2018, 24, 321–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Influenza at the Human-Animal Interface, Summary and Assessment, 16 March to 20 April 2017; World Health Organisation: Geneva, Switzerland, 2017; p. 3.
- Influenza At the Human-Animal Interface, Summary and Assessment, 21 April to 16 May 2017; World Health Organisation: Geneva, Switzerland, 2017; pp. 4–5.
- Influenza at the Human-Animal Interface, Summary and Assessment, 16 June 2017 to 25 July 2017; World Health Organisation: Geneva, Switzerland, 2017; pp. 3–4.
- Avian Influenza Report: Reporting Period: January 7, 2018–January 13, 2018 (Week 2); Hong Kong Centre for Health Protection. Available online: http://www.chp.gov.hk (accessed on 3 July 2019).
- Influenza at the Human-Animal Interface, Summary and Assessment, 26 January to 2 March 2018; World Health Organisation: Geneva, Switzerland, 2018; pp. 3–4.
- Influenza at the Human-Animal Interface, Summary and Assessment, 21 July to 21 September 2018; World Health Organisation: Geneva, Switzerland, 2018; p. 2.
- Influenza at the Human-Animal Interface, Summary and Assessment, 2 November to 13 December 2018; World Health Organisation: Geneva, Switzerland, 2018; p. 2.
- Influenza at the Human-Animal Interface, Summary and Assessment, 14 December 2018 to 21 January 2019; World Health Organisation: Geneva, Switzerland, 2019; pp. 1–2.
- Influenza at the Human-Animal Interface, Summary and Assessment, 22 January to 12 February 2019; World Health Organisation: Geneva, Switzerland, 2019; p. 2.
- Influenza at the Human-animal Interface, Summary and Assessment, 13 February to 9 April 2019; World Health Organisation: Geneva, Switzerland, 2019; pp. 2–3.
- Khan, S.U.; Anderson, B.D.; Heil, G.L.; Liang, S.; Gray, G.C. A systematic review and meta-analysis of the seroprevalence of influenza A(H9N2) infection among humans. J. Infect. Dis. 2015, 212, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Hoa, L.N.M.; Tuan, N.A.; My, P.H.; Huong, T.T.K.; Chi, N.T.Y.; Hau Thu, T.T.; Carrique-Mas, J.; Duong, M.T.; Tho, N.D.; Hoang, N.D.; et al. Assessing evidence for avian-to-human transmission of influenza A/H9N2 virus in rural farming communities in northern vietnam. J. Gen. Virol. 2017, 98, 2011–2016. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Watanabe, T.; Hatta, M.; Das, S.C.; Ozawa, M.; Shinya, K.; Zhong, G.; Hanson, A.; Katsura, H.; Watanabe, S.; et al. Experimental adaptation of an influenza h5 ha confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature 2012, 486, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Linster, M.; van Boheemen, S.; de Graaf, M.; Schrauwen, E.J.; Lexmond, P.; Manz, B.; Bestebroer, T.M.; Baumann, J.; van Riel, D.; Rimmelzwaan, G.F.; et al. Identification, characterization, and natural selection of mutations driving airborne transmission of A/H5N1 virus. Cell 2014, 157, 329–339. [Google Scholar] [CrossRef]
- Gambaryan, A.S.; Tuzikov, A.B.; Pazynina, G.V.; Desheva, J.A.; Bovin, N.V.; Matrosovich, M.N.; Klimov, A.I. 6-sulfo sialyl lewis x is the common receptor determinant recognized by H5, H6, H7 and H9 influenza viruses of terrestrial poultry. Virol. J. 2008, 5, 85. [Google Scholar] [CrossRef]
- Peacock, T.P.; Benton, D.J.; Sadeyen, J.R.; Chang, P.; Sealy, J.E.; Bryant, J.E.; Martin, S.R.; Shelton, H.; McCauley, J.W.; Barclay, W.S.; et al. Variability in H9N2 haemagglutinin receptor-binding preference and the ph of fusion. Emerg. Microbes. Infect. 2017, 6, e11. [Google Scholar] [CrossRef]
- Sauer, A.K.; Liang, C.H.; Stech, J.; Peeters, B.; Quere, P.; Schwegmann-Wessels, C.; Wu, C.Y.; Wong, C.H.; Herrler, G. Characterization of the sialic acid binding activity of influenza A viruses using soluble variants of the H7 and H9 hemagglutinins. PLoS ONE 2014, 9, e89529. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Xu, D.; Shen, W.; Liu, Q.; Rong, G.; Li, X.; Yan, L.; Yang, J.; Chen, H.; Yu, H.; et al. A single mutation at position 190 in hemagglutinin enhances binding affinity for human type sialic acid receptor and replication of H9N2 avian influenza virus in mice. J. Virol. 2016, 90, 9806–9825. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shi, J.; Guo, J.; Deng, G.; Zhang, Q.; Wang, J.; He, X.; Wang, K.; Chen, J.; Li, Y.; et al. Genetics, receptor binding property, and transmissibility in mammals of naturally isolated H9N2 avian influenza viruses. PLoS Pathog 2014, 10, e1004508. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Xu, L.; Bao, L.; Yao, Y.; Deng, W.; Li, F.; Lv, Q.; Gu, S.; Wei, Q.; Qin, C. Characterization of an H9N2 avian influenza virus from a fringilla montifringilla brambling in northern China. Virology 2015, 476, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Kaverin, N.V.; Rudneva, I.A.; Ilyushina, N.A.; Lipatov, A.S.; Krauss, S.; Webster, R.G. Structural differences among hemagglutinins of influenza A virus subtypes are reflected in their antigenic architecture: Analysis of H9 escape mutants. J. Virol. 2004, 78, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Perez, D.R. Amino acid 226 in the hemagglutinin of H9N2 influenza viruses determines cell tropism and replication in human airway epithelial cells. J. Virol. 2007, 81, 5181–5191. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Wang, A.; Ding, J.; Kong, H.; Gao, X.; Li, L.; Chai, T.; Li, Y.; Zhang, K.; Wang, C.; et al. Adaptation of H9N2 aiv in guinea pigs enables efficient transmission by direct contact and inefficient transmission by respiratory droplets. Sci. Rep. 2015, 5, 15928. [Google Scholar] [CrossRef]
- Peacock, T.P.; Harvey, W.T.; Sadeyen, J.R.; Reeve, R.; Iqbal, M. The molecular basis of antigenic variation among a(H9N2) avian influenza viruses. Emerg. Microbes Infect. 2018, 7, 176. [Google Scholar] [CrossRef]
- Thangavel, R.R.; Bouvier, N.M. Animal models for influenza virus pathogenesis, transmission, and immunology. J. Immunol. Methods 2014, 410, 60–79. [Google Scholar] [CrossRef]
- Wan, H.; Sorrell, E.M.; Song, H.; Hossain, M.J.; Ramirez-Nieto, G.; Monne, I.; Stevens, J.; Cattoli, G.; Capua, I.; Chen, L.M.; et al. Replication and transmission of H9N2 influenza viruses in ferrets: Evaluation of pandemic potential. PLoS ONE 2008, 3, e2923. [Google Scholar] [CrossRef]
- SJCEIRS H9N2 Working Group. Assessing the fitness of distinct clades of influenza A (H9N2) viruses. Emerg. Microbes Infect. 2013, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, E.M.; Wan, H.; Araya, Y.; Song, H.; Perez, D.R. Minimal molecular constraints for respiratory droplet transmission of an avian-human H9N2 influenza A virus. Proc. Natl. Acad. Sci. USA 2009, 106, 7565–7570. [Google Scholar] [CrossRef] [PubMed]
- Kimble, J.B.; Sorrell, E.; Shao, H.; Martin, P.L.; Perez, D.R. Compatibility of H9N2 avian influenza surface genes and 2009 pandemic H1N1 internal genes for transmission in the ferret model. Proc. Natl. Acad. Sci. USA 2011, 108, 12084–12088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russier, M.; Yang, G.; Rehg, J.E.; Wong, S.S.; Mostafa, H.H.; Fabrizio, T.P.; Barman, S.; Krauss, S.; Webster, R.G.; Webby, R.J.; et al. Molecular requirements for a pandemic influenza virus: An acid-stable hemagglutinin protein. Proc. Natl. Acad. Sci. USA 2016, 113, 1636–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Wang, X.; Li, Q.; Liu, D.; Chen, H.; Zhao, M.; Gu, X.; He, L.; Liu, X.; Gu, M.; et al. Molecular mechanism of the airborne transmissibility of H9N2 avian influenza A viruses in chickens. J. Virol. 2014, 88, 9568–9578. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.M.; Smith, G.J.; Bahl, J.; Duan, L.; Tai, H.; Vijaykrishna, D.; Wang, J.; Zhang, J.X.; Li, K.S.; Fan, X.H.; et al. The genesis and evolution of H9N2 influenza viruses in poultry from southern China, 2000 to 2005. J. Virol. 2007, 81, 10389–10401. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Perez, D.R. Quail carry sialic acid receptors compatible with binding of avian and human influenza viruses. Virology 2006, 346, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kimble, B.; Nieto, G.R.; Perez, D.R. Characterization of influenza virus sialic acid receptors in minor poultry species. Virol. J. 2010, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.N.; Krauss, S.; Webster, R.G. H9N2 influenza A viruses from poultry in Asia have human virus-like receptor specificity. Virology 2001, 281, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.J.; Hickman, D.; Perez, D.R. Evidence of expanded host range and mammalian-associated genetic changes in a duck H9N2 influenza virus following adaptation in quail and chickens. PLoS ONE 2008, 3, e3170. [Google Scholar] [CrossRef]
- Peiris, J.S.; Guan, Y.; Markwell, D.; Ghose, P.; Webster, R.G.; Shortridge, K.F. Cocirculation of avian H9N2 and contemporary “human” H3N2 influenza A viruses in pigs in southeastern china: Potential for genetic reassortment? J. Virol. 2001, 75, 9679–9686. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.L.; Wang, C.F.; Yan, C.M.; Peng, J.S.; Jiang, Z.L.; Liu, J.H. Swine infection with H9N2 influenza viruses in China in 2004. Virus Genes 2008, 36, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Ge, F.; Li, X.; Ju, H.; Yang, D.; Liu, J.; Qi, X.; Wang, J.; Yang, X.; Qiu, Y.; Liu, P.; et al. Genotypic evolution and antigenicity of H9N2 influenza viruses in Shanghai, China. Arch. Virol. 2016, 161, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, M.; Hong, W.; Fan, X.; Chen, R.; Zheng, Z.; Zeng, Y.; Huang, R.; Zhang, Y.; Lam, T.T.; et al. Infectivity and transmissibility of avian H9N2 influenza viruses in pigs. J. Virol. 2016, 90, 3506–3514. [Google Scholar] [CrossRef] [PubMed]
- Mancera Gracia, J.C.; Van den Hoecke, S.; Saelens, X.; Van Reeth, K. Effect of serial pig passages on the adaptation of an avian H9N2 influenza virus to swine. PLoS ONE 2017, 12, e0175267. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Kang, B.; Lee, C.; Jung, K.; Ha, G.; Kang, D.; Park, S.; Park, B.; Oh, J. Transmission of avian influenza virus (H3N2) to dogs. Emerg. Infect. Dis. 2008, 14, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, X.; Liu, Q.; Liang, D.; Li, C.; He, Q.; Jiang, J.; Cui, Y.; Li, J.; Zheng, L.; et al. Evidence of avian-like H9N2 influenza A virus among dogs in Guangxi, China. Infect. Genet. Evol. 2013, 20, 471–475. [Google Scholar] [CrossRef]
- Zhou, H.; He, S.Y.; Sun, L.; He, H.; Ji, F.; Sun, Y.; Jia, K.; Ning, Z.; Wang, H.; Yuan, L.; et al. Serological evidence of avian influenza virus and canine influenza virus infections among stray cats in live poultry markets, china. Vet. Microbiol. 2015, 175, 369–373. [Google Scholar] [CrossRef]
- Su, S.; Zhou, P.; Fu, X.; Wang, L.; Hong, M.; Lu, G.; Sun, L.; Qi, W.; Ning, Z.; Jia, K.; et al. Virological and epidemiological evidence of avian influenza virus infections among feral dogs in live poultry markets, China: A threat to human health? Clin. Infect. Dis. 2014, 58, 1644–1646. [Google Scholar] [CrossRef]
- Lee, I.H.; Le, T.B.; Kim, H.S.; Seo, S.H. Isolation of a novel H3N2 influenza virus containing a gene of H9N2 avian influenza in a dog in South Korea in 2015. Virus Genes 2016, 52, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Yong-Feng, Z.; Fei-Fei, D.; Jia-Yu, Y.; Feng-Xia, Z.; Chang-Qing, J.; Jian-Li, W.; Shou-Yu, G.; Kai, C.; Chuan-Yi, L.; Xue-Hua, W.; et al. Intraspecies and interspecies transmission of mink H9N2 influenza virus. Sci. Rep. 2017, 7, 7429. [Google Scholar] [CrossRef] [PubMed]
- He, Q. Isolation and whole genome sequence analysis of equine H9N2 influenza virus in Guangxi. Master’s Thesis, Guangxi University, Nanning, China, 2012. [Google Scholar]
- Akerstedt, J.; Valheim, M.; Germundsson, A.; Moldal, T.; Lie, K.I.; Falk, M.; Hungnes, O. Pneumonia caused by influenza A H1N1 2009 virus in farmed American mink (neovison vison). Vet. Rec. 2012, 170, 362. [Google Scholar] [CrossRef] [PubMed]
- Englund, L. Studies on influenza viruses H10N4 and H10N7 of avian origin in mink. Vet. Microbiol. 2000, 74, 101–107. [Google Scholar] [CrossRef]
- Peng, L.; Chen, C.; Kai-yi, H.; Feng-xia, Z.; Yan-li, Z.; Zong-shuai, L.; Xing-xiao, Z.; Shi-jin, J.; Zhi-jing, X. Molecular characterization of H9N2 influenza virus isolated from mink and its pathogenesis in mink. Vet. Microbiol. 2015, 176, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xuan, Y.; Shan, H.; Yang, H.; Wang, J.; Wang, K.; Li, G.; Qiao, J. Avian influenza virus H9N2 infections in farmed minks. Virol. J. 2015, 12, 180. [Google Scholar] [CrossRef] [PubMed]
- Capuano, A.M.; Miller, M.; Stallknecht, D.E.; Moriarty, M.; Plancarte, M.; Dodd, E.; Batac, F.; Boyce, W.M. Serologic detection of subtype-specific antibodies to influenza A viruses in southern sea otters (enhydra lutris nereis). J. Wildl. Dis. 2017, 53, 906–910. [Google Scholar] [CrossRef]
- Yu, Z.; Cheng, K.; Sun, W.; Xin, Y.; Cai, J.; Ma, R.; Zhao, Q.; Li, L.; Huang, J.; Sang, X.; et al. Lowly pathogenic avian influenza (H9N2) infection in plateau pika (ochotona curzoniae), Qinghai lake, China. Vet. Microbiol. 2014, 173, 132–135. [Google Scholar] [CrossRef]
- Yan, Y.; Gu, J.Y.; Yuan, Z.C.; Chen, X.Y.; Li, Z.K.; Lei, J.; Hu, B.L.; Yan, L.P.; Xing, G.; Liao, M.; et al. Genetic characterization of H9N2 avian influenza virus in plateau pikas in the Qinghai lake region of China. Arch. Virol. 2017, 162, 1025–1029. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, H.; Huang, C.; Sun, H.; Li, L.; Su, J.; Ma, J.; Liu, D.; Wang, H.; Liu, W.; et al. Distribution of sialic acid receptors and experimental infections with different subtypes of influenza A viruses in Qinghai-Tibet plateau wild pika. Virol. J. 2015, 12, 63. [Google Scholar] [CrossRef]
- Kandeil, A.; Gomaa, M.R.; Shehata, M.M.; El Taweel, A.N.; Mahmoud, S.H.; Bagato, O.; Moatasim, Y.; Kutkat, O.; Kayed, A.S.; Dawson, P.; et al. Isolation and characterization of a distinct influenza A virus from Egyptian bats. J. Virol. 2019, 93, e01059-18. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Song, C.S. H9N2 avian influenza virus in Korea: Evolution and vaccination. Clin. Exp. Vaccine Res. 2013, 2, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Naeem, K.; Siddique, N. Use of strategic vaccination for the control of avian influenza in Pakistan. Dev. Biol. 2006, 124, 145–150. [Google Scholar] [PubMed]
- Kilany, W.H.; Ali, A.; Bazid, A.H.; El-Deeb, A.H.; El-Abideen, M.A.; Sayed, M.E.; El-Kady, M.F. A dose-response study of inactivated low pathogenic avian influenza H9N2 virus in specific-pathogen-free and commercial broiler chickens. Avian Dis. 2016, 60, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Bahari, P.; Pourbakhsh, S.A.; Shoushtari, H.; Bahmaninejad, M.A. Molecular characterization of H9N2 avian influenza viruses isolated from vaccinated broiler chickens in northeast Iran. Trop. Anim. Health Prod. 2015, 47, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.Y.; Joseph, S.; Chan, K.H.; Chen, H.; Patteril, N.A.; Elizabeth, S.K.; Muhammed, R.; Baskar, V.; Lau, S.K.; Kinne, J.; et al. Complete genome sequence of influenza virus H9N2 associated with a fatal outbreak among chickens in Dubai. Genome Announc. 2016, 4, e00752-16. [Google Scholar] [CrossRef]
- Sealy, J.E.; Yaqub, T.; Peacock, T.P.; Chang, P.; Ermetal, B.; Clements, A.; Sadeyen, J.R.; Mehboob, A.; Shelton, H.; Bryant, J.E.; et al. Association of increased receptor-binding avidity of influenza A(H9N2) viruses with escape from antibody-based immunity and enhanced zoonotic potential. Emerg. Infect. Dis. 2018, 25, 63–72. [Google Scholar] [CrossRef]
- Offeddu, V.; Cowling, B.J.; Malik Peiris, J.S. Interventions in live poultry markets for the control of avian influenza: A systematic review. One Health 2016, 2, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Fournie, G.; Hog, E.; Barnett, T.; Pfeiffer, D.U.; Mangtani, P. A systematic review and meta-analysis of practices exposing humans to avian influenza viruses, their prevalence, and rationale. Am. J. Trop. Med. Hyg. 2017, 97, 376–388. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Country | Years of Poultry Isolates | Lineages | Species | Status | Recorded Human Cases/Serology |
|---|---|---|---|---|---|
| Afghanistan | 2008–2009, 2016–2017 | G1-W | Chicken | Potentially endemic | No |
| Algeria | 2017 | G1-W | Chicken | Potentially endemic | No |
| Bangladesh | 2006–2007, 2009–present | G1-W, Y439 | Chicken, Quail | Endemic | Virus isolated |
| Belgium | 19833 | Y439 | H9N2-free | No | |
| Burkina Faso | 2017 | G1-W | Chicken | Potentially endemic | No |
| Cambodia | 2013, 2015, 2017–present | BJ94, G1-E, Y439 | Chicken | Likely endemic | Serology only |
| Colombia | 2005 | n/a | Chicken | Unknown | No |
| China | 1994–present | BJ94, G1-E, Y439 | Chicken, Guinea Fowl, Quail, Partridge, | Endemic | Virus isolated and serology |
| Egypt | 2006, 2011–2018 | G1-W | Chicken, Quail, Turkey | Endemic | Virus isolated and serology |
| France | 1998, 2003 | Y439 | Turkey | H9N2-free | No |
| Germany | 1994–1996, 19983, 2004, 2012–2013, 2015–2017 | Y439, G1-W | Chicken, Turkey | Recurrent infections from wild birds | No |
| Ghana | 2017–present | G1-W | Chicken | Potentially endemic | No |
| Hong Kong SAR | 1988, 1992, 1994, 1997, 1999–2000 2003, 2005–2012, 2014–2015 | BJ94, G1-E, Y439 | Chicken, Quail, Guinea Fowl, Partridge | Potentially endemic | Virus isolated and serology |
| Hungary | 2001 | Y439 | Chicken | H9N2-free | No |
| India | 2003–2004, 2006–2013, 2015 | G1-W | Chicken | Potentially endemic | Serology only |
| Indonesia | 2002, 2016–present | BJ94, Y439 | Chicken | Likely endemic | No |
| Iran | 1998–2017 | G1-W | Chicken | Endemic | Serology only |
| Iraq | 2005, 2008, 2014–2016 | G1-W | Chicken | Potentially endemic | No |
| Israel 1 | 2000–2014, 2016–2017 | G1-W | Chicken, Turkey, Ostrich | Potentially endemic | No |
| Italy | 1983–1985, 1989, 1994, 1996 | Y439 | Chicken, Turkey | H9N2-free | No |
| Japan (Imported goods only) | 1997, 2001–2002, 2015–2016 | BJ94 | Imported meat | H9N2-free | No |
| Jordan | 2003–2007, 2010 | G1-W | Chicken | Likely endemic | No |
| Kuwait | 2003–2005, 2008 | G1-W | Chicken | Potentially endemic | No |
| Laos 2 | 2009 | n/a | Chickens | Potentially endemic | No |
| Lebanon | 2004, 2010, 2017–present | G1-W | Chicken, Quail | Potentially endemic | No |
| Libya | 2005, 2013 | G1-W | Chicken | Potentially endemic | No |
| Malaysia | 2018 | n/a | n/a | Potentially endemic | No |
| Morocco | 2016 | G1-W | Chicken | Potentially endemic | No |
| Myanmar | 2014–2015 | BJ94 | Chicken | Potentially endemic | No |
| Nepal | 2009–2011 | G1-W | Chicken | Potentially endemic | No |
| Netherlands | 2010–2011 | Y439 | Chicken, Turkey | H9N2-free | No |
| Nigeria 2 | 2013 | n/a | Chicken | Potentially endemic | Serology only |
| Oman | 2006, 2019 | G1-W | Chicken | Potentially endemic | Virus isolated |
| Pakistan | 1998–2000, 2003–2012, 2014–2017, 2019 | G1-W | Chicken, Partridge | Endemic | Virus isolated and serology |
| Poland | 2013–2014 | Y439 | Turkey | H9N2-free | No |
| Qatar | 2008 | G1 | Chicken | Potentially endemic | No |
| Romania 2 | 2009–2010 | n/a | n/a | Unknown | Serology only |
| Russia (Eastern) | 2018 | G1-W, BJ94 | Chicken | Unknown | No |
| Saudi Arabia | 1998–2000, 2002, 2005––2008, 2010–2011, 2013, 2015–2016, 2018 | G1-W | Chicken | Potentially endemic | No |
| South Africa | 1995, 2008–2009 | Y439 | Ostrich | H9N2-free | No |
| South Korea | 1996, 1999–2012 | Y439(Korean) | Chicken, Guinea Fowl | Potentially endemic | No |
| Thailand 2 | 2008 | n/a | n/a | Potentially endemic | Serology only |
| Tunisia | 2010–2012, 2014 | G1-W | Chicken, Turkey | Potentially endemic | No |
| USA | 1966, 1978, 1981, 1983, 1985, 1988–1989, 1993, 1995, 1997, 1999, 2001 | USA | Chicken, Quail, Turkey | H9N2-free | No |
| UAE | 1999–2003, 2005–2006, 2008, 2011, 2015 | G1-W | Chicken, Quail | Potentially endemic | No |
| Uganda | 2017 | G1-W | Chicken | Potentially endemic | No |
| UK | 1970, 2010, 2013 | Y439 | Chicken, Turkey | H9N2-free | No |
| Vietnam | 2009, 2012–2017 | BJ94, G1-E, Y439 | Chicken, Quail | Likely endemic | Serology only |
| Year | Location | Patient | Clinical Signs | Viral Lineage | Poultry Exposure? | Reference |
|---|---|---|---|---|---|---|
| 1998 | Guangdong province, China | 14-year-old, male | ARI a | BJ94 | Yes, live chickens in dwelling | [116] |
| 75-year-old, male | ARI | BJ94 | Yes, lived near farmers market | [116] | ||
| 4-year-old, male | ARI | BJ94 | Unknown | [116] | ||
| 1-year-old, female | ARI | BJ94 | Unknown | [116] | ||
| 36-year-old, female | ARI | BJ94 | Yes, exposure to live poultry | [116] | ||
| 1999 | Guangdong province, China | 22-month-old, female | Fever, cough | BJ94 | No | [125] |
| Hong Kong | 13-month-old, female | Fever | G1 ‘Eastern’ | Yes | [115] | |
| 4-year-old, female | Fever, malaise | G1 ‘Eastern’ | Unknown | [115] | ||
| 2003 | Hong Kong | 5-year-old, male | Fever, cough | BJ94 | No | [126] |
| 2004 | Guangdong Province, China | Unknown | Unknown | G1 ‘Eastern’ | Unknown | [127] |
| 2007 | Hong Kong | 9-month-old, female | Mild illness | ND c | Yes | [128] |
| 2008 | Guangdong province, China | 2-month-old, female | ILI b | ND | Unknown | [128] |
| 2009 | Hong Kong | 35-month-old, female | Fever, cough, | G1 ‘Eastern’ | Unknown | [129] |
| 47-year-old, female | Fever, cough | G1 ‘Eastern’ | No | [130] | ||
| 2011 | Dhaka, Bangladesh | 4-year-old, female | Fever, cough | G1 ‘Western’ | Yes, close exposure to sick poultry | [117] |
| 2013 | Guangdong province, China | 86-year-old, male | Cough | BJ94 | No | [131] |
| Hunan province, China | 7-year-old, male | Fever | BJ94 | Yes, close contact to poultry | [74,131] | |
| 2014 | Sichuan Province, China | 2.5-year-old, male | Mild illness | BJ94 | Unknown | [119,132] |
| Guangdong province, China | Unknown | Mild illness | BJ94 | Unknown | [119] | |
| 2015 | Aswan, Egypt | 3-year-old, male | Unknown | ND | Yes | [119] |
| Cairo, Egypt | 7-year-old, female | ILI | ND | Yes | [119] | |
| 9-month-old, female | ILI | ND | Yes | [119] | ||
| Guangdong province, China | male | Unknown | BJ94 | Unknown | [133] | |
| Bangladesh | 3.5-year-old, female | Mild illness | ND | Yes, close contact with sick poultry | [134] | |
| Anhui province, China | 4-year-old, female | Mild illness | BJ94 | Yes, live bird market exposure | [135] | |
| Hunan province, China | 2-year-old, male | Mild illness | BJ94 | Unknown | [75,135] | |
| Anhui province, China | 6-year-old, male | Unknown | BJ94 | Unknown | [136] | |
| Hunan province, China | 15-year-old, female | Mild illness | ND | No | [75,135] | |
| 11-month-old, female | Mild illness | ND | No | [135] | ||
| Dhaka, Bangladesh | 46-year-old, male | Fever | ND | Yes, poultry worker, exposure to sick birds | [137] | |
| Guangdong province, China | 84-year-old, female | Unknown | ND | Unknown | [121] | |
| Punjab district, Pakistan | 36-year-old, male | Non-symptomatic | G1 ‘Western’ | Yes | [118] | |
| Hunan province, China | 2-year-old, male | Mild illness | BJ94 | Yes, live bird market exposure | [138] | |
| 2016 | Sichuan Province, China | 57-year-old, female | ARI, Died d | ND | Unknown | [139] |
| Cairo, Egypt | 18-month-old, male | ILI | ND | Yes, exposure to live bird market | [140] | |
| Guangdong province, China | 4-year-old, female | ARI | BJ94 | Yes | [141,142] | |
| 29-year-old, female | ARI | ND | Unknown | [121] | ||
| Yunnan province, China | 10-month-old, male | ILI | ND | Yes | [121] | |
| Jiangxi province, China | 4-year-old, female | Mild illness | ND | Unknown | [121] | |
| Henan province, China | 5-year-old, female | Unknown | ND | No | [121] | |
| Guangdong province, China | 3-year-old, male | Unknown | ND | Yes | [121] | |
| Guangdong province, China | 7-month-old, female | Mild illness | ND | Yes | [143] | |
| Beijing, China | 4-month-old, male | Mild illness | BJ94 | Yes | [144] | |
| 2017 | Gansu province, China | 11-month-old, male | Mild illness | ND | Yes | [145] |
| Beijing, China | 32-year-old, male | Mild illness | BJ94 | No | [144,146] | |
| Guangdong province, China | 2-month-old, female | ILI | ND | Yes, poultry at home | [147] | |
| Hunan province, China | 20-month-old, female | n/a | BJ94 | Unknown | [148] | |
| 9-month-old, male | ILI | BJ94 | Unknown | [75,148] | ||
| Anhui province, China | 9-year-old, female | Mild illness | BJ94 | Unknown | [149] | |
| 2018 | Guangdong province, China | 3-year-old, female | Mild illness | ND | Yes, exposure to live bird market | [149] |
| Beijing, China | 51-year-old, female | Mild illness | ND | Yes, exposure to slaughtered poultry | [149] | |
| Guangdong province, China | 24-year-old, female (pregnant) | Mild illness | ND | Yes, exposure to farm | [150] | |
| Guangdong province, China | 10-month-old, female | Mild illness | ND | Yes, backyard poultry exposure | [151] | |
| Guangxi province, China | 3-year-old, male | n/a | BJ94 | No | [151] | |
| Guangdong province, China | 32-year-old, female | Pneumonia | ND | Unknown | [152] | |
| Hunan province, China | 2-year-old, male | Mild illness | BJ94 | No | [153] | |
| 2019 | Yunnan province, China | 8-year-old, female | Mild illness | ND | No | [153] |
| Jiangsu province, China | 9-year-old, male | Severe pneumonia | ND | Yes | [154] | |
| Oman | 13-month-old, female | ILI | G1 ‘Western’ | Yes | [120] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peacock, T.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. https://doi.org/10.3390/v11070620
Peacock TP, James J, Sealy JE, Iqbal M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses. 2019; 11(7):620. https://doi.org/10.3390/v11070620
Chicago/Turabian StylePeacock, T. (Thomas) P., Joe James, Joshua E. Sealy, and Munir Iqbal. 2019. "A Global Perspective on H9N2 Avian Influenza Virus" Viruses 11, no. 7: 620. https://doi.org/10.3390/v11070620
APA StylePeacock, T. P., James, J., Sealy, J. E., & Iqbal, M. (2019). A Global Perspective on H9N2 Avian Influenza Virus. Viruses, 11(7), 620. https://doi.org/10.3390/v11070620