Viral Genomic Characterization and Replication Pattern of Human Polyomaviruses in Kidney Transplant Recipients

 , ,
, ,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. DNA Isolation and Quantitative Real-Time PCR Assay
2.3. HPyVs Molecular Characterization
2.3.1. Molecular Characterization of JCPyV
2.3.2. Molecular Characterization of BKPyV
2.3.3. Molecular Characterization of MCPyV
2.4. DNA Automatic Sequencing
2.5. Immunosuppression and Concomitant Medications
2.6. Statistical Analysis
3. Results
3.1. HPyVs Viruria in the Study Cohort
3.2. HPyVs Viruria in Urinary Samples
3.3. HPyVs Urinary Load
3.4. HPyVs Molecular Characterization
3.4.1. JCPyV Molecular Characterization
3.4.2. BKPyV Molecular Characterization
3.4.3. MCPyV Molecular Characterization
3.5. HPyV-Associated Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feltkamp, M.C.W.; Kazem, S.; van der Meijden, E.; Lauber, C.; Gorbalenya, A.E. From Stockholm to Malawi: Recent developments in studying human polyomaviruses. J. Gen. Virol. 2013, 94, 482–496. [Google Scholar] [CrossRef]
- Cook, L. Polyomaviruses. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Wolfe, R.A.; Ashby, V.B.; Milford, E.L.; Ojo, A.O.; Ettenger, R.E.; Agodoa, L.Y.; Held, P.J.; Port, F.K. Comparison of mortality in all patients on dialysis, patients on dialysis awaiting transplantation, and recipients of a first cadaveric transplant. N. Engl. J. Med. 1999, 341, 1725–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, H.H.; Drachenberg, C.B.; Steiger, J.; Ramos, E. Polyomavirus-associated nephropathy in renal transplantation: Critical issues of screening and management. Adv. Exp. Med. Biol. 2006, 577, 160–173. [Google Scholar] [PubMed]
- Cukuranovic, J.; Ugrenovic, S.; Jovanovic, I.; Visnjic, M.; Stefanovic, V. Viral infection in renal transplant recipients. ScientificWorldJournal 2012, 2012, 820621. [Google Scholar] [CrossRef] [PubMed]
- Kartau, M.; Sipilä, J.O.; Auvinen, E.; Palomäki, M.; Verkkoniemi-Ahola, A. Progressive Multifocal Leukoencephalopathy: Current Insights. Degener. Neurol. Neuromuscul. Dis. 2019, 9, 109–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeCaprio, J.A.; Garcea, R.L. A cornucopia of human polyomaviruses. Nat. Rev. Microbiol. 2013, 11, 264–276. [Google Scholar] [CrossRef]
- Moens, U.; Calvignac-Spencer, S.; Lauber, C.; Ramqvist, T.; Feltkamp, M.C.W.; Daugherty, M.D.; Verschoor, E.J.; Ehlers, B.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Polyomaviridae. J. Gen. Virol. 2017, 98, 1159–1160. [Google Scholar]
- Schowalter, R.M.; Buck, C.B. The Merkel cell polyomavirus minor capsid protein. PLoS Pathog. 2013, 9, e1003558. [Google Scholar] [CrossRef] [Green Version]
- Spurgeon, M.E.; Lambert, P.F. Merkel cell polyomavirus: A newly discovered human virus with oncogenic potential. Virology 2013, 435, 118–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerits, N.; Moens, U. Agnoprotein of mammalian polyomaviruses. Virology 2012, 432, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Prado, J.C.M.; Monezi, T.A.; Amorim, A.T.; Lino, V.; Paladino, A.; Boccardo, E. Human polyomaviruses and cancer: An overview. Clinics (Sao Paulo) 2018, 73 (Suppl. S1), e558s. [Google Scholar] [CrossRef]
- Khalili, K.; Stoner, G.L. Human Polyomaviruses: Molecular and Clinical Perspectives; Wiley-Liss: Hoboken, NJ, USA, 2001; p. 688. [Google Scholar]
- Agostini, H.T.; Deckhut, A.; Jobes, D.V.; Girones, R.; Schlunck, G.; Prost, M.G.; Frias, C.; Pérez-Trallero, E.; Ryschkewitsch, C.F.; Stoner, G.L. Genotypes of JC virus in East, Central and Southwest Europe. J. Gen. Virol. 2001, 82, 1221–1331. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, C.; Kitamura, T.; Guo, J.; Al-Ahdal, M.N.; Shchelkunov, S.N.; Otova, B.; Ondrejka, P.; Chollet, J.Y.; El-Safi, S.; Ettayebi, M.; et al. Typing of urinary JC virus DNA offers a novel means of tracing human migrations. Proc. Natl. Acad. Sci. USA 1997, 94, 9191–9196. [Google Scholar] [CrossRef] [Green Version]
- Agostini, H.T.; Yanagihara, R.; Davis, V.; Ryschkewitsch, C.F.; Stoner, G.L. Asian genotypes of JC virus in Native Americans and in a Pacific Island population: Markers of viral evolution and human migration. Proc. Natl. Acad. Sci. USA 1997, 94, 14542–14546. [Google Scholar] [CrossRef] [Green Version]
- Jobe, D.V.; Friedlaender, J.S.; Mgone, C.S.; Agostini, H.T.; Koki, G.; Yanagihara, R.; Ng, T.C.N.; Chima, S.C.; Ryschkewitsch, C.F.; Stoner, G.L. New JC virus (JCV) genotypes from papua new guinea and micronesia (type 8 and type 2E) and evolutionary analysis of 32 complete JCV genomes. Arch. Virol. 2001, 146, 2097–2113. [Google Scholar] [CrossRef] [PubMed]
- Ferenczy, M.W.; Marshall, L.J.; Nelson, C.D.; Atwood, W.J.; Nath, A.; Khalili, K.; Major, E.O. Molecular biology, epidemiology, and pathogenesis of progressive multifocal leukoencephalopathy, the JC virus-induced demyelinating disease of the human brain. Clin. Microbiol. Rev. 2012, 25, 471–506. [Google Scholar] [CrossRef] [Green Version]
- Ault, G.S.; Stoner, G.L. Human polyomavirus JC promoter/enhancer rearrangement patterns from progressive multifocal leukoencephalopathy brain are unique derivatives of a single archetypal structure. J. Gen. Virol. 1993, 74, 1499–1507. [Google Scholar] [CrossRef]
- Agostini, H.T.; Ryschkewitsch, C.F.; Singer, E.J.; Stoner, G.L. JC virus regulatory region rearrangements and genotypes in progressive multifocal leukoencephalopathy: Two independent aspects of virus variation. J. Gen. Virol. 1997, 78, 659–664. [Google Scholar] [CrossRef] [Green Version]
- Jobes, D.V.; Chima, S.C.; Ryschkewitsch, C.F.; Stoner, G.L. Phylogenetic analysis of 22 complete genomes of the human polyomavirus JC virus. J. Gen. Virol. 1998, 79, 2491–2498. [Google Scholar] [CrossRef] [Green Version]
- Pagani, E.; Delbue, S.; Mancuso, R.; Borghi, E.; Tarantini, L.; Ferrante, P. Molecular analysis of JC virus genotypes circulating among the Italian healthy population. J. Neurovirol. 2003, 9, 559–566. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Nishimoto, Y.; Chen, Q.; Hasegawa, M.; Zhong, S.; Ikegaya, H.; Ohno, N.; Sugimoto, C.; Takasaka, T.; Kitamura, T.; et al. Relationships between BK virus lineages and human populations. Microbes Infect. 2007, 9, 204–213. [Google Scholar] [CrossRef]
- Martel-Jantin, C.; Pedergnana, V.; Nicol, J.T.; Leblond, V.; Trégouët, D.A.; Tortevoye, P.; Plancoulaine, S.; Coursaget, P.; Touzé, A.; Abel, L.; et al. Merkel cell polyomavirus infection occurs during early childhood and is transmitted between siblings. J. Clin. Virol. 2013, 58, 288–291. [Google Scholar] [CrossRef] [Green Version]
- Gosert, R.; Rinaldo, C.H.; Funk, G.A.; Egli, A.; Ramos, E.; Drachenberg, C.B.; Hirsch, H.H. Polyomavirus BK with rearranged noncoding control region emerge in vivo in renal transplant patients and increase viral replication and cytopathology. J. Exp. Med. 2008, 205, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Olsen, G.H.; Hirsch, H.H.; Rinaldo, C.H. Functional analysis of polyomavirus BK non-coding control region quasispecies from kidney transplant recipients. J. Med. Virol. 2009, 81, 1959–1967. [Google Scholar] [CrossRef]
- Gosert, R.; Kardas, P.; Major, E.O.; Hirsch, H.H. Rearranged JC virus noncoding control regions found in progressive multifocal leukoencephalopathy patient samples increase virus early gene expression and replication rate. J. Virol. 2010, 84, 10448–10456. [Google Scholar] [CrossRef] [Green Version]
- Ajuh, E.T.; Wu, Z.; Kraus, E.; Weissbach, F.H.; Bethge, T.; Gosert, R.; Fischer, N.; Hirsch, H.H. Novel Human Polyomavirus Noncoding Control Regions Differ in Bidirectional Gene Expression according to Host Cell, Large T-Antigen Expression, and Clinically Occurring Rearrangements. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotta, N.; Delbue, S.; Signorini, L.; Villani, S.; D’Alessandro, S.; Campisciano, G.; Colli, C.; De Seta, F.; Ferrante, P.; Comar, M. Merkel Cell Polyomavirus Is Associated with Anal Infections in Men Who Have Sex with Men. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Delbue, S.; Pagani, E.; Guerini, F.R.; Agliardi, C.; Mancuso, R.; Borghi, E.; Rossi, F.; Boldorini, R.; Veggiani, C.; Car, P.G.; et al. Distribution, characterization and significance of polyomavirus genomic sequences in tumors of the brain and its covering. J. Med. Virol. 2005, 77, 447–454. [Google Scholar] [CrossRef]
- Gorelik, L.; Reid, C.; Testa, M.; Brickelmaier, M.; Bossolasco, S.; Pazzi, A.; Bestetti, A.; Carmillo, P.; Wilson, E.; McAuliffe, M.; et al. Progressive multifocal leukoencephalopathy (PML) development is associated with mutations in JC virus capsid protein VP1 that change its receptor specificity. J. Infect. Dis. 2011, 204, 103–114. [Google Scholar] [CrossRef]
- Johnsen, J.I.; Seternes, O.M.; Johansen, T.; Moens, U.; Mäntyjärvi, R.; Traavik, T. Subpopulations of non-coding control region variants within a cell culture-passaged stock of BK virus: Sequence comparisons and biological characteristics. J. Gen. Virol. 1995, 76, 1571–1581. [Google Scholar] [CrossRef]
- Flaegstad, T.; Sundsfjord, A.; Arthur, R.R.; Pedersen, M.; Traavik, T.; Subramani, S. Amplification and sequencing of the control regions of BK and JC virus from human urine by polymerase chain reaction. Virology 1991, 180, 553–560. [Google Scholar] [CrossRef]
- Anzivino, E.; Bellizzi, A.; Mitterhofer, A.P.; Tinti, F.; Barile, M.; Colosimo, M.T.; Fioriti, D.; Mischitelli, M.; Chiarini, F.; Ferretti, G.; et al. Early monitoring of the human polyomavirus BK replication and sequencing analysis in a cohort of adult kidney transplant patients treated with basiliximab. Virol. J. 2011, 8, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashida, Y.; Higuchi, T.; Matsui, K.; Shibata, Y.; Nakajima, K.; Sano, S.; Daibata, M. Genetic Variability of the Noncoding Control Region of Cutaneous Merkel Cell Polyomavirus: Identification of Geographically Related Genotypes. J. Infect. Dis. 2018, 217, 1601–1611. [Google Scholar] [CrossRef] [Green Version]
- Bofill-Mas, S.; Rodriguez-Manzano, J.; Calgua, B.; Carratala, A.; Girones, R. Newly described human polyomaviruses Merkel cell, KI and WU are present in urban sewage and may represent potential environmental contaminants. Virol. J. 2010, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- Agostini, H.T.; Ryschkewitsch, C.F.; Stoner, G.L. Genotype profile of human polyomavirus JC excreted in urine of immunocompetent individuals. J. Clin. Microbiol. 1996, 34, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.N.; Major, E.O. A classification scheme for human polyomavirus JCV variants based on the nucleotide sequence of the noncoding regulatory region. J. Neurovirol. 2001, 7, 280–287. [Google Scholar]
- Jin, L.; Gibson, P.E.; Knowles, W.A.; Clewley, J.P. BK virus antigenic variants: Sequence analysis within the capsid VP1 epitope. J. Med. Virol. 1993, 39, 50–56. [Google Scholar] [CrossRef]
- Van Loon, E.; Bernards, J.; Van Craenenbroeck, A.H.; Naesens, M. The Causes of Kidney Allograft Failure: More Than Alloimmunity. A Viewpoint Article. Transplantation 2020, 104, e46–e56. [Google Scholar] [CrossRef]
- Favi, E.; James, A.; Puliatti, C.; Whatling, P.; Ferraresso, M.; Rui, C.; Cacciola, R. Utility and safety of early allograft biopsy in adult deceased donor kidney transplant recipients. Clin. Exp. Nephrol. 2020, 24, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Kotton, C.N.; Fishman, J.A. Viral infection in the renal transplant recipient. J. Am. Soc. Nephrol. 2005, 16, 1758–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishman, J.A. Infection in Organ Transplantation. Am. J. Transplant. 2017, 17, 856–879. [Google Scholar] [CrossRef] [Green Version]
- Moroni, G.; Binda, V.; Quaglini, S.; Sacchi, L.; Raffiotta, F.; Cosa, F.; Montagnino, G.; Favi, E.; Messa, P.; Ponticelli, C. Causes of late transplant failure in cyclosporine-treated kidney allograft recipients. Clin. Exp. Nephrol 2019, 23, 1076–1086. [Google Scholar] [CrossRef]
- Mishra, N.; Pereira, M.; Rhodes, R.H.; An, P.; Pipas, J.M.; Jain, K.; Kapoor, A.; Briese, T.; Faust, P.L.; Lipkin, W.I. Identification of a novel polyomavirus in a pancreatic transplant recipient with retinal blindness and vasculitic myopathy. J. Infect. Dis. 2014, 210, 1595–1599. [Google Scholar] [CrossRef] [Green Version]
- Kwak, E.J.; Vilchez, R.A.; Randhawa, P.; Shapiro, R.; Butel, J.S.; Kusne, S. Pathogenesis and management of polyomavirus infection in transplant recipients. Clin. Infect. Dis. 2002, 35, 1081–1087. [Google Scholar] [CrossRef]
- Knowles, W.A. Discovery and epidemiology of the human polyomaviruses BK virus (BKV) and JC virus (JCV). Adv. Exp. Med. Biol. 2006, 577, 19–45. [Google Scholar]
- Jiang, M.; Abend, J.R.; Johnson, S.F.; Imperiale, M.J. The role of polyomaviruses in human disease. Virology 2009, 384, 266–273. [Google Scholar] [CrossRef] [Green Version]
- Polo, C.; Pérez, J.L.; Mielnichuck, A.; Fedele, C.G.; Niubò, J.; Tenorio, A. Prevalence and patterns of polyomavirus urinary excretion in immunocompetent adults and children. Clin. Microbiol. Infect. 2004, 10, 640–644. [Google Scholar] [CrossRef] [Green Version]
- Jeong, B.H.; Lee, K.H.; Choi, E.K.; Kim, K.; Kim, Y.S. Genotyping of the JC virus in urine samples of healthy Korean individuals. J. Med. Virol. 2004, 72, 281–289. [Google Scholar] [CrossRef]
- Signorini, L.; Belingheri, M.; Ambrogi, F.; Pagani, E.; Binda, S.; Ticozzi, R.; Ferraresso, M.; Ghio, L.; Giacon, B.; Ferrante, P.; et al. High frequency of Merkel cell polyomavirus DNA in the urine of kidney transplant recipients and healthy controls. J. Clin. Virol. 2014, 61, 565–570. [Google Scholar] [CrossRef]
- Chehadeh, W.; Kurien, S.S.; Nampoory, M.R. Molecular characterization of BK and JC viruses circulating among potential kidney donors in Kuwait. Biomed. Res. Int. 2013, 2013, 683464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boukoum, H.; Nahdi, I.; Abid, N.; Foulongne, V.; Sahtout, W.; Zallema, D.; Skiri, H.; Aloui, S.; Achour, A.; Segondy, M.; et al. Distribution of JC polyomavirus genotypes in Tunisian renal transplant recipients between January 2008 and January 2011. J. Med. Virol. 2012, 84, 1818–1824. [Google Scholar] [CrossRef]
- Comerlato, J.; Campos, F.S.; Oliveira, M.T.; Cibulski, S.P.; Corrêa, L.; Kulmann, M.I.; Arantes, T.S.; Hentges, L.P.; Spilki, F.R.; Roehe, P.M.; et al. Molecular detection and characterization of BK and JC polyomaviruses in urine samples of renal transplant patients in Southern Brazil. J. Med. Virol. 2015, 87, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Husseiny, M.I.; Anastasi, B.; Singer, J.; Lacey, S.F. A comparative study of Merkel cell, BK and JC polyomavirus infections in renal transplant recipients and healthy subjects. J. Clin. Virol. 2010, 49, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Drachenberg, C.B.; Hirsch, H.H.; Papadimitriou, J.C.; Gosert, R.; Wali, R.K.; Munivenkatappa, R.; Nogueira, J.; Cangro, C.B.; Haririan, A.; Mendley, S.; et al. Polyomavirus BK versus JC replication and nephropathy in renal transplant recipients: A prospective evaluation. Transplantation 2007, 84, 323–330. [Google Scholar] [CrossRef]
- Bialasiewicz, S.; Rockett, R.J.; Barraclough, K.A.; Leary, D.; Dudley, K.J.; Isbel, N.M.; Sloots, T.P. Detection of Recently Discovered Human Polyomaviruses in a Longitudinal Kidney Transplant Cohort. Am. J. Transplant. 2016, 16, 2734–2740. [Google Scholar] [CrossRef] [Green Version]
- Favi, E.; Puliatti, C.; Sivaprakasam, R.; Ferraresso, M.; Ambrogi, F.; Delbue, S.; Gervasi, F.; Salzillo, I.; Raison, N.; Cacciola, R. Incidence, risk factors, and outcome of BK polyomavirus infection after kidney transplantation. World J. Clin. Cases 2019, 7, 270–290. [Google Scholar] [CrossRef]
- Seppälä, H.M.; Helanterä, I.T.; Laine, P.K.S.; Lautenschlager, I.T.; Paulín, L.G.; Jahnukainen, T.J.; Auvinen, P.O.V.; Auvinen, E. Archetype JC Polyomavirus (JCPyV) Prevails in a Rare Case of JCPyV Nephropathy and in Stable Renal Transplant Recipients With JCPyV Viruria. J. Infect. Dis. 2017, 216, 981–989. [Google Scholar] [CrossRef]
- Yin, W.Y.; Lu, M.C.; Lee, M.C.; Liu, S.C.; Lin, T.Y.; Lai, N.S. A correlation between polyomavirus JC virus quantification and genotypes in renal transplantation. Am. J. Surg. 2010, 200, 53–58. [Google Scholar] [CrossRef]
- Baksh, F.K.; Finkelstein, S.D.; Swalsky, P.A.; Stoner, G.L.; Ryschkewitsch, C.F.; Randhawa, P. Molecular genotyping of BK and JC viruses in human polyomavirus-associated interstitial nephritis after renal transplantation. Am. J. Kidney Dis. 2001, 38, 354–365. [Google Scholar] [CrossRef]
- Chehadeh, W.; Nampoory, M.R. Genotypic diversity of polyomaviruses circulating among kidney transplant recipients in Kuwait. J. Med. Virol. 2013, 85, 1624–1631. [Google Scholar] [CrossRef]
- Saundh, B.K.; Baker, R.; Harris, M.; Hale, A. A prospective study of renal transplant recipients reveals an absence of primary JC polyomavirus infections. J. Clin. Virol. 2016, 77, 101–105. [Google Scholar] [CrossRef]
- Cubitt, C.L. Molecular genetics of the BK virus. Adv. Exp. Med. Biol. 2006, 577, 85–95. [Google Scholar]
- Olsen, G.H.; Andresen, P.A.; Hilmarsen, H.T.; Bjørang, O.; Scott, H.; Midtvedt, K.; Rinaldo, C.H. Genetic variability in BK Virus regulatory regions in urine and kidney biopsies from renal-transplant patients. J. Med. Virol. 2006, 78, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Perets, T.T.; Silberstein, I.; Rubinov, J.; Sarid, R.; Mendelson, E.; Shulman, L.M. High frequency and diversity of rearrangements in polyomavirus bk noncoding regulatory regions cloned from urine and plasma of Israeli renal transplant patients and evidence for a new genetic subtype. J. Clin. Microbiol. 2009, 47, 1402–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalona, A.C.; Lopes, G.S.; Schrago, C.G.; Gonçalves, R.T.; Zalis, M.G.; Varella, R.B. Molecular characterization of BK polyomavirus subtypes in renal transplant recipients in Brazil. J. Med. Virol. 2011, 83, 1401–1405. [Google Scholar] [CrossRef] [PubMed]


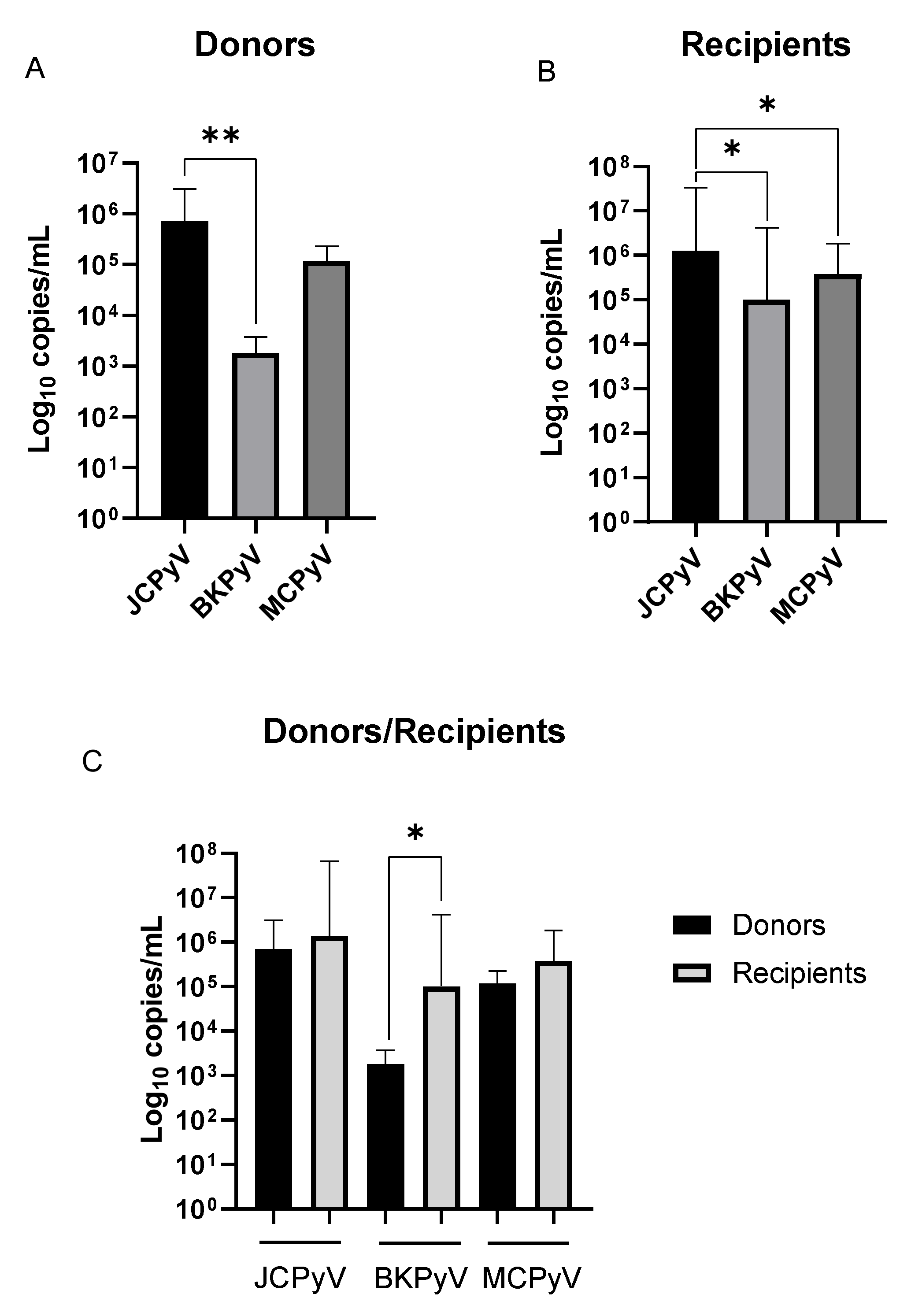{kind=link}
| Donors (n = 57) | Recipients (n = 57) | |
|---|---|---|
| Variables | Mean ± SD or n (%) | |
| Male:female | 26:24 * | 32:25 |
| Country of origin ≠ Italy | 4/53 ## (7.5) | 7/50 ### (14) |
| Age (years) | 55.2 ± 13.2 ** | 44.3 ± 17.9 *** |
| Living:deceased | 23:29 # | - |
| Basiliximab | - | 31/52 $ (59.6) |
| Tacrolimus | - | 51/52 $ (98.1) |
| Mycophenolate mofetil | - | 50/52 $ (96.1) |
| Days After Transplantation | 0 | 1 | 15 | 30 | 60 | 90 | 180 | 270 | 360 | 540 |
|---|---|---|---|---|---|---|---|---|---|---|
| Code | T0 | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 |
| JCPyV +/tot (%) | BKPyV +/tot (%) | MCPyV +/tot (%) | HPyV/tot (%) | |
|---|---|---|---|---|
| Donors * | 25/28 (89.3) #,$,& | 3/28 (10.7) ^,$ | 2/28 (7.1) °,& | 28/57 (49.1) + |
| Recipients ** | 25/44 (56.8) # | 21/44 (47.7) ^ | 17/44 (38.6) ° | 44/57 (77.2) + |
| Total | 50 α/72 (69.4) §,£ | 24 β/72 (33.3) § | 19 π/72 (26.4) £ | 72/114 (63.1) |
| JCPyV +/tot (%) | BKPyV +/tot (%) | MCPyV +/tot (%) | HPyV/tot (%) |
|---|---|---|---|
| 172/551 (31.2) *,°,# | 62/551 (11.2) *,^,#,$ | 38/551 (6.9) °,^,#,$ | 272/551 (49.4) |
| Median Viral Load Copies/mL (IQR *) | ||||
|---|---|---|---|---|
| JCPyV | BKPyV | MCPyV | HPyVs | |
| Donors | 7.11 × 105 ° (1.17 × 106) | 1.80 × 103 °,$ (1.58 × 103) | 1.18 × 105 (1.10 × 105) | 3.64 × 105 (1.15 × 106) |
| Recipients | 1.29 × 106 #,& (3.16 × 107) | 1.02 × 105 #,$ (3.67 × 106) | 3.76 × 105 & (1.47 × 106) | 5.65 × 105 (1.54 × 107) |
| Total | 1.01 × 106 (2.57 × 107) | 6.39 × 104 (2.60 × 106) | 3.67 × 105 (1.34 × 106) | 5.42 × 105 (1.16 × 107) |
| JCPyV Genotype | Positive Donors (n = 24) (%) | Positive Recipients (n = 24) (%) |
|---|---|---|
| 1a | 6 (25.0%) | 7* (25.0%) |
| 1b | 7 (29.2%) | 7 (29.2%) |
| 2a | 1 (4.2%) | 1 (4.2%) |
| 2c | 3 (12.5%) | 2 (8.3%) |
| 4 | 5 (20.8%) | 6 * (25.0%) |
| 6 | 1 (4.2%) | 1 (4.2%) |
| Without identification | 1 (4.2%) | - |
| BKPyV Genotype | Positive Donors (n = 3) (%) | Positive Recipients (n = 13) (%) |
|---|---|---|
| Ia | 1 (33.3%) | 1 (7.7%) |
| Ib-1 | 2 (66.7%) | 8 (61.5%) |
| Ib-2 | - | 4 (30.8%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Signorini, L.; Dolci, M.; Favi, E.; Colico, C.; Ferraresso, M.; Ticozzi, R.; Basile, G.; Ferrante, P.; Delbue, S. Viral Genomic Characterization and Replication Pattern of Human Polyomaviruses in Kidney Transplant Recipients. Viruses 2020, 12, 1280. https://doi.org/10.3390/v12111280
Signorini L, Dolci M, Favi E, Colico C, Ferraresso M, Ticozzi R, Basile G, Ferrante P, Delbue S. Viral Genomic Characterization and Replication Pattern of Human Polyomaviruses in Kidney Transplant Recipients. Viruses. 2020; 12(11):1280. https://doi.org/10.3390/v12111280
Chicago/Turabian StyleSignorini, Lucia, Maria Dolci, Evaldo Favi, Caterina Colico, Mariano Ferraresso, Rosalia Ticozzi, Giuseppe Basile, Pasquale Ferrante, and Serena Delbue. 2020. "Viral Genomic Characterization and Replication Pattern of Human Polyomaviruses in Kidney Transplant Recipients" Viruses 12, no. 11: 1280. https://doi.org/10.3390/v12111280
APA StyleSignorini, L., Dolci, M., Favi, E., Colico, C., Ferraresso, M., Ticozzi, R., Basile, G., Ferrante, P., & Delbue, S. (2020). Viral Genomic Characterization and Replication Pattern of Human Polyomaviruses in Kidney Transplant Recipients. Viruses, 12(11), 1280. https://doi.org/10.3390/v12111280