
Inactivation of Human Coronavirus by Titania Nanoparticle Coatings and UVC Radiation: Throwing Light on SARS-CoV-2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Human Coronavirus
2.3. TiO2 Nanoparticles (TNPs)-Coated Glass Coverslips
2.4. UV Photocatalysis
2.5. Virus Infectivity and Immunofluorescence Assay (IFA)
2.6. RNA Extraction and qPCR
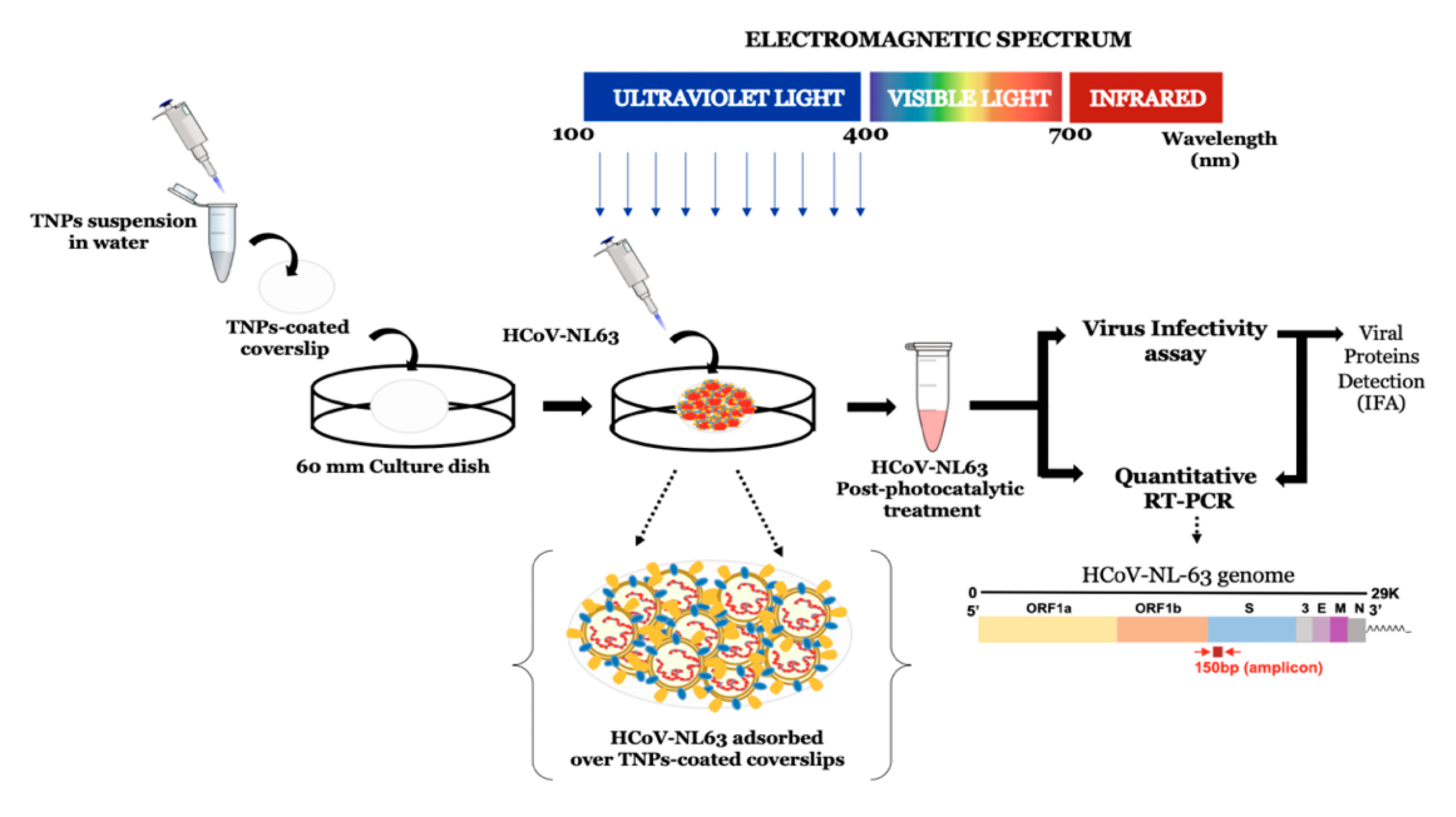
2.7. Experimental Setup
2.8. Statistical Analysis
3. Results
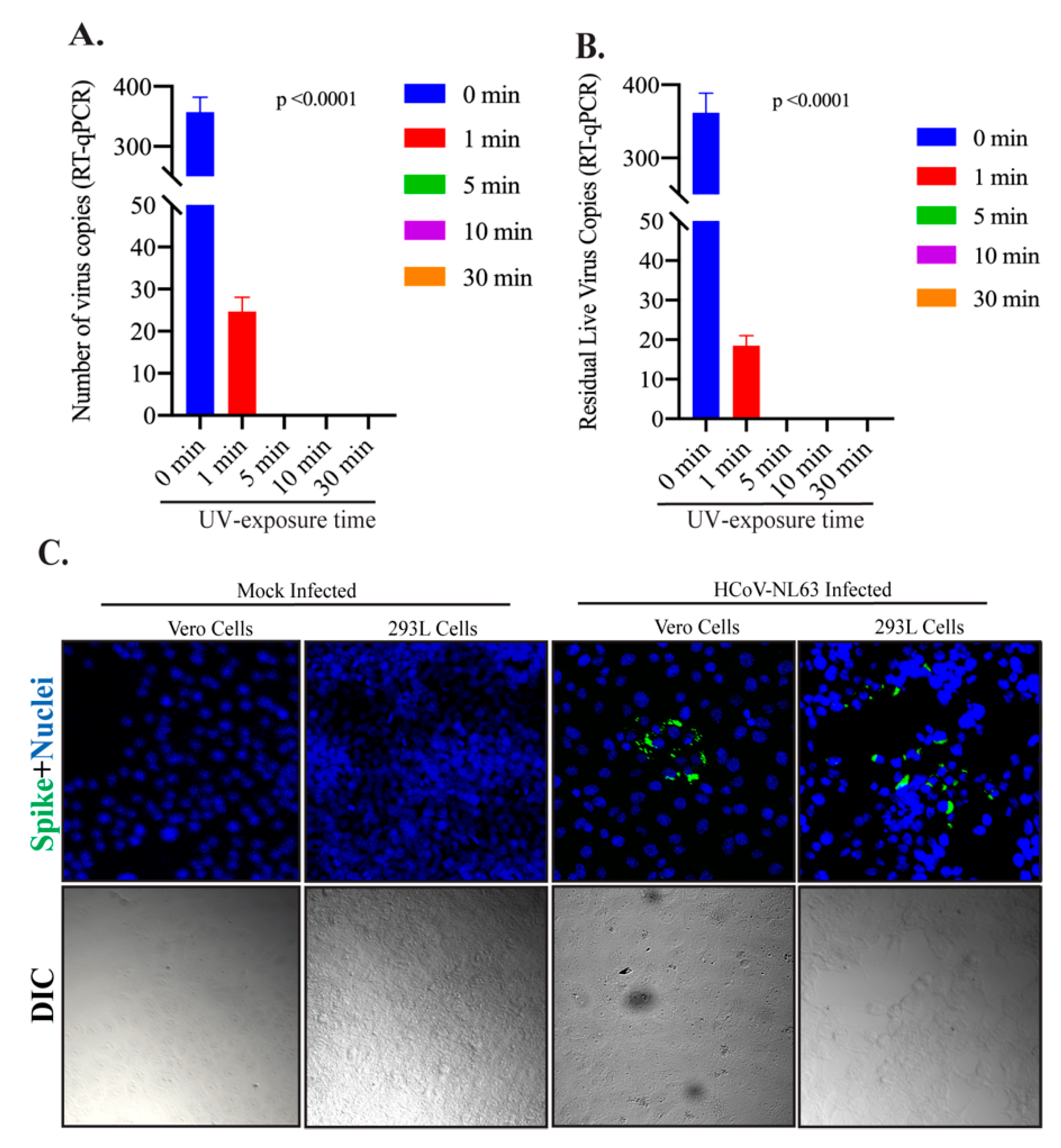
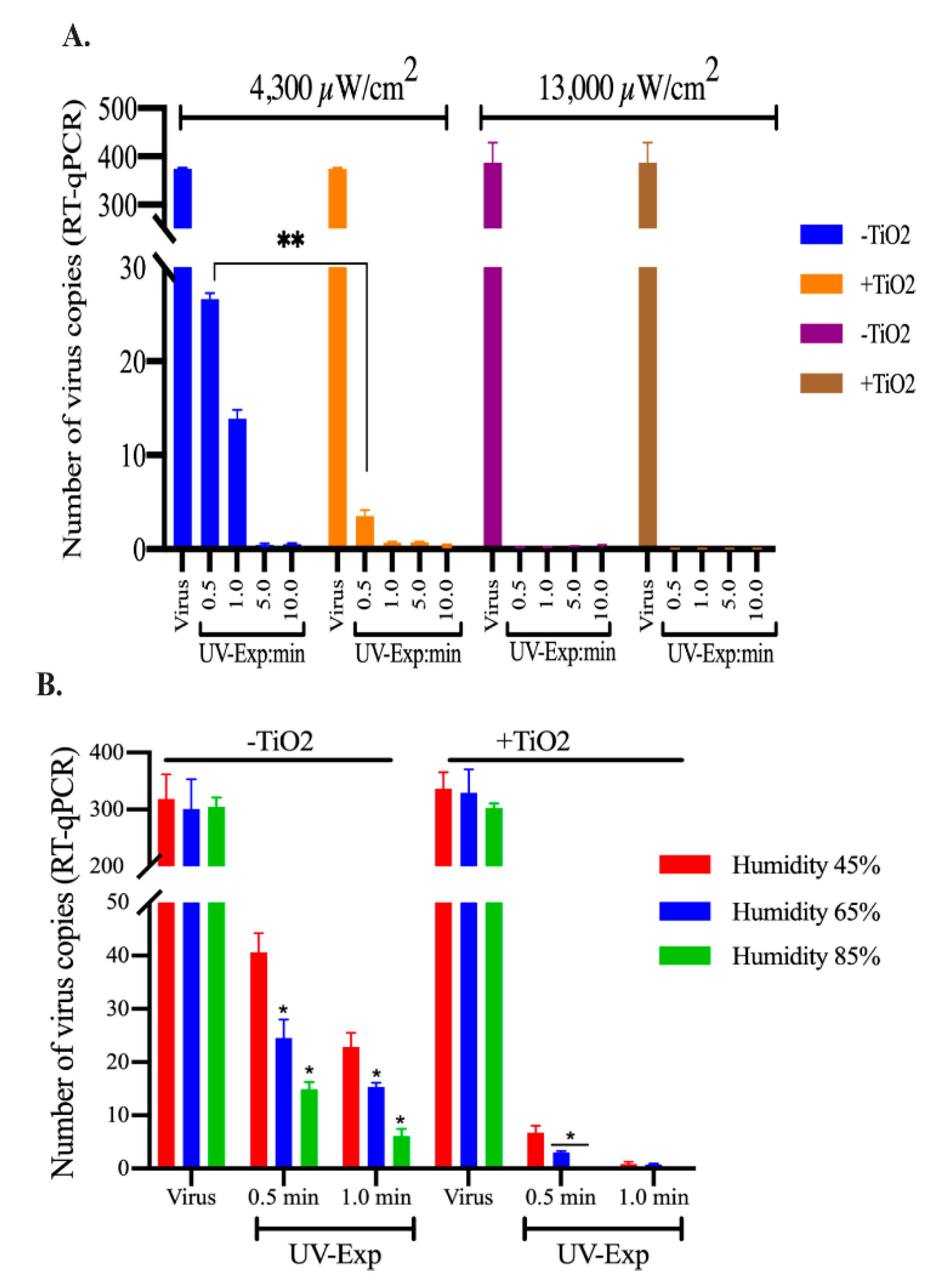
3.1. The Effect of UV Light Exposure on HCoV-NL63 Virus Stability
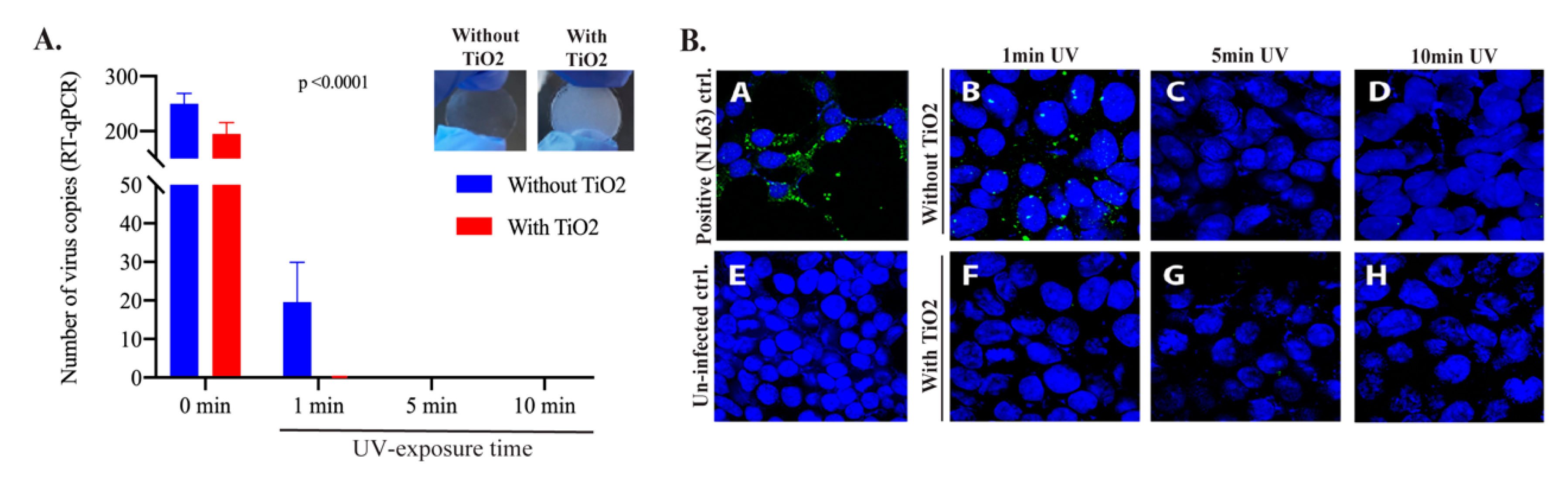
3.2. TNP Coating Efficiently Inactivated HCoV-NL63 Even at Brief Exposures
3.3. TiO2 Coatings Effectively Inactivated HCoV-NL63 Virus Present in Wet or Dried Form
3.4. Reducing the Exposure Distance and Increasing the Relative Humidity Enhanced HCoV-NL63 Inactivation
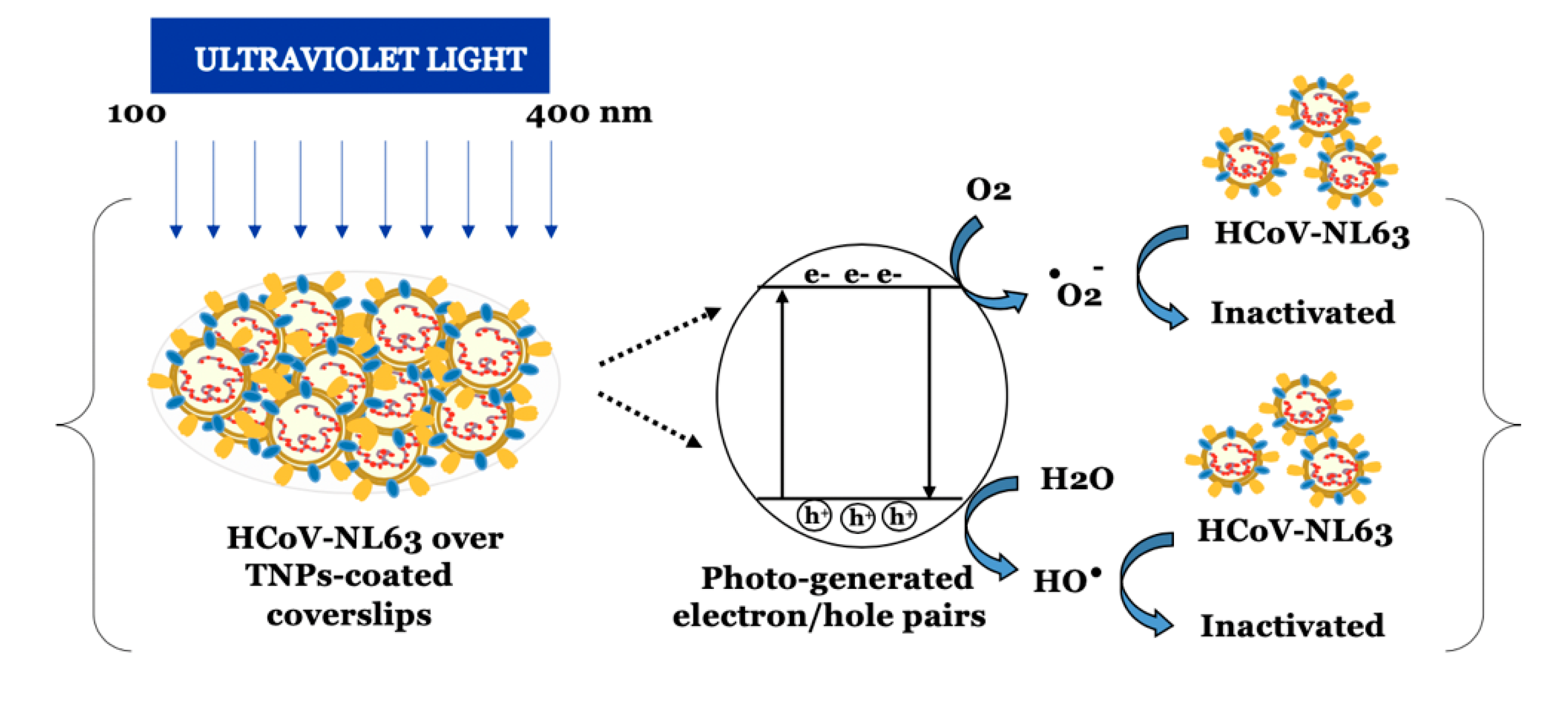
4. Discussion
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CENTER:, C.R. COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). Available online: https://coronavirus.jhu.edu/map.html (accessed on 20 December 2020).
- Organization:, W.H. Coronavirus Disease 2019 (COVID-19). Situation Report, 30. 2020. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 20 December 2020).
- CDC. Provisional Death Counts for Coronavirus Disease (Covid-19). Available online: https://www.cdc.gov/nchs/nvss/vsrr/covid19/ (accessed on 20 December 2020).
- Shao, L.; Li, X.; Zhou, Y.; Yu, Y.; Liu, Y.; Liu, M.; Zhang, R.; Zhang, H.; Wang, X.; Zhou, F. Novel Insights Into Illness Progression and Risk Profiles for Mortality in Non-survivors of COVID-19. Front Med. (Lausanne) 2020, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.; Kakara, R. Deaths from Falls Among Persons Aged ≥65 Years-United States, 2007–2016. Mmwr. Morb. Mortal. Wkly. Rep. 2018, 67, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I. Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, S.; Yang, Z.; Guan, W.; Liu, D.; Lin, Z.; Zhang, Y.; Xu, Z.; Liu, X.; Li, Y. SARS-CoV-2 viral load in clinical samples of critically ill patients. Am. J. Respir. Crit. Care Med. 2020, 201, 1435–1438. [Google Scholar] [CrossRef] [Green Version]
- Morawska, L. Droplet fate in indoor environments, or can we prevent the spread of infection? In Proceedings of the Indoor Air 2005: The 10th International Conference on Indoor Air Quality and Climate, Beijing, China, 4–9 September 2005; pp. 9–23. [Google Scholar]
- Wang, J.; Du, G. COVID-19 may transmit through aerosol. Ir. J. Med Sci. 2020, 189, 1143–1144. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Coronavirus Disease (COVID-19) Advice for the Public; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Ong, S.W.X.; Tan, Y.K.; Chia, P.Y.; Lee, T.H.; Ng, O.T.; Wong, M.S.Y.; Marimuthu, K. Air, surface environmental, and personal protective equipment contamination by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) from a symptomatic patient. JAMA 2020, 323, 1610–1612. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ning, Z.; Chen, Y.; Guo, M.; Liu, Y.; Gali, N.K.; Sun, L.; Duan, Y.; Cai, J.; Westerdahl, D. Aerodynamic analysis of SARS-CoV-2 in two Wuhan hospitals. Nature 2020, 582, 557–560. [Google Scholar] [CrossRef]
- Cai, J.; Sun, W.; Huang, J.; Gamber, M.; Wu, J.; He, G. Indirect Virus Transmission in Cluster of COVID-19 Cases, Wenzhou, China, 2020. Emerg. Infect. Dis. 2020, 26, 1–3. [Google Scholar] [CrossRef]
- Bogdan, J.; Zarzyńska, J.; Pławińska-Czarnak, J. Comparison of infectious agents susceptibility to photocatalytic effects of nanosized titanium and zinc oxides: A practical approach. Nanoscale Res. Lett. 2015, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nakano, R.; Ishiguro, H.; Yao, Y.; Kajioka, J.; Fujishima, A.; Sunada, K.; Minoshima, M.; Hashimoto, K.; Kubota, Y. Photocatalytic inactivation of influenza virus by titanium dioxide thin film. Photochem. Photobiol. Sci. 2012, 11, 1293–1298. [Google Scholar] [CrossRef]
- Mazurkova, N.; Spitsyna, Y.E.; Shikina, N.; Ismagilov, Z.; Zagrebel’nyi, S.; Ryabchikova, E. Interaction of titanium dioxide nanoparticles with influenza virus. Nanotechnol. Russ. 2010, 5, 417–420. [Google Scholar] [CrossRef]
- Abdul-Rasool, S.; Fielding, B.C. Understanding human coronavirus HCoV-NL63. Open Virol. J. 2010, 4, 76. [Google Scholar] [CrossRef]
- ATCC. Human Coronavirus 229E (ATCC® VR-740™). Available online: https://www.lgcstandards-atcc.org/products/all/VR-740.aspx?geo_country=ru (accessed on 20 December 2020).
- Current, R. Annual Average Relative Humidity by US State. Available online: https://www.currentresults.com/Weather/US/annual-average-humidity-by-state.php (accessed on 20 December 2020).
- CDC. Indoor Environmental Quality. Available online: https://www.cdc.gov/niosh/topics/indoorenv/temperature.html#:~:text=The%20difference%20in%20temperature%20ranges,mold%20growth%20%5BEPA%202012%5D (accessed on 20 December 2020).
- Zhou, J.; Otter, J.; Price, J.R.; Cimpeanu, C.; Garcia, D.M.; Kinross, J.; Boshier, P.; Mason, S.; Bolt, F.; Holmes, A.H. Investigating SARS-CoV-2 surface and air contamination in an acute healthcare setting during the peak of the COVID-19 pandemic in London. Clin. Infect. Dis. 2020, 1–22. [Google Scholar] [CrossRef]
- Guo, Z.-D.; Wang, Z.-Y.; Zhang, S.-F.; Li, X.; Li, L.; Li, C.; Cui, Y.; Fu, R.-B.; Dong, Y.-Z.; Chi, X.-Y. Early Release-Aerosol and Surface Distribution of Severe Acute Respiratory Syndrome Coronavirus 2 in Hospital Wards, Wuhan, China, 2020. Emerg. Infect. Dis. 2020, 26, 1–6. [Google Scholar] [CrossRef]
- Suo, T.; Liu, X.; Feng, J.; Guo, M.; Hu, W.; Guo, D.; Ullah, H.; Yang, Y.; Zhang, Q.; Wang, X. ddPCR: A more accurate tool for SARS-CoV-2 detection in low viral load specimens. Emerg. Microbes Infect. 2020, 9, 1–30. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.-F.; Xu, W.; Liu, S.-W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1–9. [Google Scholar] [CrossRef]
- Desai, A.N.; Patel, P. Stopping the Spread of COVID-19. JAMA 2020, 323, 1516. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.; Chu, J.; Perera, M. Correspondence. Stability of SARS-CoV-2 in different environmental conditions. Lancet Microbe 2020, 1, 10. [Google Scholar] [CrossRef]
- Casanova, L.M.; Jeon, S.; Rutala, W.A.; Weber, D.J.; Sobsey, M.D. Effects of air temperature and relative humidity on coronavirus survival on surfaces. Appl. Environ. Microbiol. 2010, 76, 2712–2717. [Google Scholar] [CrossRef] [Green Version]
- Gralton, J.; Tovey, E.; McLaws, M.-L.; Rawlinson, W.D. The role of particle size in aerosolised pathogen transmission: A review. J. Infect. 2011, 62, 1–13. [Google Scholar] [CrossRef]
- Tatlıdil, İ.; Sökmen, M.; Breen, C.; Clegg, F.; Buruk, C.K.; Bacaksız, E. Degradation of Candida albicans on TiO2 and Ag-TiO2 thin films prepared by sol–gel and nanosuspensions. J. Sol.-Gel. Sci. Technol. 2011, 60, 23. [Google Scholar] [CrossRef]
- Panayotov, D.A.; Burrows, S.P.; Morris, J.R. Photooxidation mechanism of methanol on rutile TiO2 nanoparticles. J. Phys. Chem. C 2012, 116, 6623–6635. [Google Scholar] [CrossRef]
- Vatansever, F.; de Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R. Antimicrobial strategies centered around reactive oxygen species–bactericidal antibiotics, photodynamic therapy, and beyond. Fems Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [Green Version]
- Yates, H.; Brook, L.; Ditta, I.; Evans, P.; Foster, H.; Sheel, D.; Steele, A. Photo-induced self-cleaning and biocidal behaviour of titania and copper oxide multilayers. J. Photochem. Photobiol. A Chem. 2008, 197, 197–205. [Google Scholar] [CrossRef]
- van der Molen, R.G.; Garssen, J.; de Klerk, A.; Claas, F.H.; Norval, M.; van Loveren, H.; Koerten, H.K.; Mommaas, A.M. Application of a systemic herpes simplex virus type 1 infection in the rat as a tool for sunscreen photoimmunoprotection studies. Photochem. Photobiol. Sci. 2002, 1, 592–596. [Google Scholar] [CrossRef]
- Zan, L.; Fa, W.; Peng, T.; Gong, Z.-K. Photocatalysis effect of nanometer TiO2 and TiO2-coated ceramic plate on Hepatitis B virus. J. Photochem. Photobiol. B Biol. 2007, 86, 165–169. [Google Scholar] [CrossRef]
- Monmaturapoj, N.; Sri-on, A.; Klinsukhon, W.; Boonnak, K.; Prahsarn, C. Antiviral activity of multifunctional composite based on TiO2-modified hydroxyapatite. Mater. Sci. Eng. C 2018, 92, 96–102. [Google Scholar] [CrossRef]
- Akhtar, S.; Shahzad, K.; Mushtaq, S.; Ali, I.; Rafe, M.H.; Fazal-ul-Karim, S.M. Antibacterial and antiviral potential of colloidal Titanium dioxide (TiO2) nanoparticles suitable for biological applications. Mater. Res. Express 2019, 6, 105409. [Google Scholar] [CrossRef]
- Han, W.; Zhang, B.; Cao, W.; Yang, D.; TAIRA, I.; Okamoto, Y.; Arai, J.; Yan, X. The inactivation effect of photocatalytic titanium apatite filter on SARS virus. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Jin Zhan 2004, 31, 982–985. [Google Scholar]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDevitt, J.J.; Rudnick, S.N.; Radonovich, L.J. Aerosol susceptibility of influenza virus to UV-C light. Appl. Environ. Microbiol. 2012, 78, 1666–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.-C.; Li, C.-S. Inactivation of virus-containing aerosols by ultraviolet germicidal irradiation. Aerosol Sci. Technol. 2005, 39, 1136–1142. [Google Scholar] [CrossRef]
- Buonanno, M.; Welch, D.; Shuryak, I.; Brenner, D.J. Far-UVC light (222 nm) efficiently and safely inactivates airborne human coronaviruses. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Bianco, A.; Biasin, M.; Pareschi, G.; Cavalleri, A.; Cavatorta, C.; Fenizia, F.; Galli, P.; Lessio, L.; Lualdi, M.; Redaelli, E. UV-C irradiation is highly effective in inactivating and inhibiting SARS-CoV-2 replication. Inact. Inhib. SARS-CoV-2 Replication 2020, 1–9. [Google Scholar] [CrossRef]
- Wilson, B.D.; Moon, S.; Armstrong, F. Comprehensive review of ultraviolet radiation and the current status on sunscreens. J. Clin. Aesthetic Dermatol. 2012, 5, 18. [Google Scholar]
- Arena, L.B.; Karagiozis, A.; Mantha, P. Monitoring of Internal Moisture Loads in Residential Buildings—Research Findings in Three Different Climate Zones. In Proceedings of the Buildings XI: Thermal Performance of Exterior Envelopes of Whole Buildings International Conference, Clearwater Beach, FL, USA, 5–9 December 2010; Available online: http://web.ornl.gov/sci/buildings/2012/2010%20B11%20papers/75_Arena.pdf (accessed on 20 December 2020).
- Berry, A.; Fillaux, J.; Martin-Blondel, G.; Boissier, J.; Iriart, X.; Marchou, B.; Magnaval, J.F.; Delobel, P. Evidence for a permanent presence of schistosomiasis in Corsica, France, 2015. Eurosurveillance 2016, 21, 30100. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.-H.; Peiris, J.M.; Lam, S.; Poon, L.; Yuen, K.; Seto, W.H. The effects of temperature and relative humidity on the viability of the SARS coronavirus. Adv. Virol. 2011, 2011, 1–7. [Google Scholar] [CrossRef]
- Yang, W.; Marr, L.C. Dynamics of airborne influenza A viruses indoors and dependence on humidity. PLoS ONE 2011, 6, e21481. [Google Scholar] [CrossRef] [Green Version]
- Sooryanarain, H.; Elankumaran, S. Environmental role in influenza virus outbreaks. Annu. Rev. Anim. Biosci. 2015, 3, 347–373. [Google Scholar] [CrossRef]
- Weber, T.P.; Stilianakis, N.I. Inactivation of influenza A viruses in the environment and modes of transmission: A critical review. J. Infect. 2008, 57, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jing, W.; Liu, J.; Ma, Q.; Yuan, J.; Wang, Y.; Du, M.; Liu, M. Effects of temperature and humidity on the daily new cases and new deaths of COVID-19 in 166 countries. Sci. Total Environ. 2020, 729, 139051. [Google Scholar] [CrossRef] [PubMed]
- Matson, M.J.; Yinda, C.K.; Seifert, S.N.; Bushmaker, T.; Fischer, R.J.; van Doremalen, N.; Lloyd-Smith, J.O.; Munster, V.J. Early Release-Effect of Environmental Conditions on SARS-CoV-2 Stability in Human Nasal Mucus and Sputum. Emerg. Infect. Dis. 2020, 26, 2276–2278. [Google Scholar] [CrossRef] [PubMed]
- Sizun, J.; Yu, M.; Talbot, P. Survival of human coronaviruses 229E and OC43 in suspension and after drying onsurfaces: A possible source ofhospital-acquired infections. J. Hosp. Infect. 2000, 46, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; Von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosc, F.; Lacroix-Desmazes, P.; Ayral, A. TiO2 anatase-based membranes with hierarchical porosity and photocatalytic properties. J. Colloid Interface Sci. 2006, 304, 545–548. [Google Scholar] [CrossRef]
- De Jong, B.; Meeder, A.; Koekkoek, K.; Schouten, M.; Westers, P.; Van Zanten, A. Pre–post evaluation of effects of a titanium dioxide coating on environmental contamination of an intensive care unit: The TITANIC study. J. Hosp. Infect. 2018, 99, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Visai, L.; De Nardo, L.; Punta, C.; Melone, L.; Cigada, A.; Imbriani, M.; Arciola, C.R. Titanium oxide antibacterial surfaces in biomedical devices. Int. J. Artif. Organs 2011, 34, 929–946. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khaiboullina, S.; Uppal, T.; Dhabarde, N.; Subramanian, V.R.; Verma, S.C. Inactivation of Human Coronavirus by Titania Nanoparticle Coatings and UVC Radiation: Throwing Light on SARS-CoV-2. Viruses 2021, 13, 19. https://doi.org/10.3390/v13010019
Khaiboullina S, Uppal T, Dhabarde N, Subramanian VR, Verma SC. Inactivation of Human Coronavirus by Titania Nanoparticle Coatings and UVC Radiation: Throwing Light on SARS-CoV-2. Viruses. 2021; 13(1):19. https://doi.org/10.3390/v13010019
Chicago/Turabian StyleKhaiboullina, Svetlana, Timsy Uppal, Nikhil Dhabarde, Vaidyanathan Ravi Subramanian, and Subhash C. Verma. 2021. "Inactivation of Human Coronavirus by Titania Nanoparticle Coatings and UVC Radiation: Throwing Light on SARS-CoV-2" Viruses 13, no. 1: 19. https://doi.org/10.3390/v13010019
APA StyleKhaiboullina, S., Uppal, T., Dhabarde, N., Subramanian, V. R., & Verma, S. C. (2021). Inactivation of Human Coronavirus by Titania Nanoparticle Coatings and UVC Radiation: Throwing Light on SARS-CoV-2. Viruses, 13(1), 19. https://doi.org/10.3390/v13010019