
Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cercospora beticola Isolates and Growth Conditions
2.2. Total RNA Extraction
2.3. High-Throughput Sequencing and Sequence Analysis
2.4. DNA Extraction
2.5. Confirmation of the Putative Mycoviruses
2.6. Phylogenetic Analysis
3. Results and Discussion
3.1. Mycoviruses Identification from C. beticola
3.2. Sequences Related to (+)ssRNA Viruses
3.2.1. One Predicted Novel Virus in Family Hypoviridae
3.2.2. One Strain of Previously Reported Narnaviruses
3.2.3. Three Predicted Novel Viruses in Family Botourmiaviridae
3.3. Sequences Related to (−)ssRNA Viruses
3.4. Sequence Related to ssRNA-RT Viruses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holtschulte, B. Cercospora beticola-worldwide distribution and incidence. In Cercospora Beticola Sacc. Biology, Agronomic Influence and Control Measures in Sugar Beet; Asher, M.J.C., Holtschulte, B., Molard, M.R., Rosso, F., Steinrucken, G., Beckers, R., Eds.; International Institute for Beet Research: Brussels, Belgium, 2000; pp. 5–16. [Google Scholar]
- Rangel, L.I.; Spanner, R.E.; Ebert, M.K.; Pethybridge, S.J.; Stukenbrock, E.H.; de Jonge, R.; Secor, G.A.; Bolton, M.D. Cercospora beticola: The intoxicating lifestyle of the leaf spot pathogen of sugar beet. Mol. Plant Pathol. 2020, 21, 1020–1041. [Google Scholar] [CrossRef]
- Giannopolitis, C.N. Occurrence of strains of Cercospora beticola resistant to triphenyltin fungicides in Greece. Plant Dis. Report. 1978, 62, 205–208. [Google Scholar]
- Bolton, M.D.; Birla, K.; Rivera-Varas, V.; Rudolph, K.D.; Secor, G.A. Characterization of CbCyp51 from field isolates of Cercospora beticola. Phytopathology 2012, 102, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiland, J.J.; Halloin, J.M. Benzimidazole resistance in Cercospora beticola sampled from sugarbeet fields in Michigan, USA 1. Can. J. Plant Pathol. 2001, 23, 78–82. [Google Scholar] [CrossRef]
- Trueman, C.L.; Hanson, L.E.; Somohano, P.; Rosenzweig, N. First report of DMI-insensitive Cercospora beticola on sugar beet in Ontario, Canada. New Dis. Rep. 2017, 36, 20. [Google Scholar] [CrossRef] [Green Version]
- Housni, Z.E.; Ezrari, S.; Tahiri, A.; Ouijja, A.; Lahlali, R. First Report of Benzimidazole, DMI and QoI-insensitive Cercospora beticola in sugar beet in Morocco. New Dis. Rep. 2018, 38, 17. [Google Scholar] [CrossRef] [Green Version]
- Hudec, K.; Mihók, M.; Roháčik, T.; Mišľan, Ľ. Sensitivity of Cercospora beticola to fungicides in Slovakia. Acta Fytotech. Et Zootech. 2020, 23, 147–154. [Google Scholar] [CrossRef]
- Mu, F.; Xie, J.; Cheng, S.; You, M.P.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T.; et al. Virome characterization of a collection of S. sclerotiorum from Australia. Front. Microbiol. 2018, 8, 2540. [Google Scholar] [CrossRef] [Green Version]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef]
- Yao, Z.; Zou, C.; Peng, N.; Zhu, Y.; Bao, Y.; Zhou, Q.; Wu, Q.; Chen, B.; Zhang, M. Virome identification and characterization of Fusarium sacchari and F. Andiyazi: Causative agents of pokkah boeng disease in sugarcane. Front. Microbiol. 2020, 11, 240. [Google Scholar] [CrossRef] [Green Version]
- Chiapello, M.; Rodríguez-Romero, J.; Ayllón, M.A.; Turina, M. Analysis of the virome associated to grapevine downy mildew lesions reveals new mycovirus lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef]
- Shapira, R.; Choi, G.H.; Nuss, D.L. Virus-like genetic organization and expression strategy for a double-stranded RNA genetic element associated with biological control of chestnut blight. EMBO J. 1991, 10, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Yu, J.; Kim, K.-H. Five questions about mycoviruses. PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef]
- Li, H.; Bian, R.; Liu, Q.; Yang, L.; Pang, T.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L. Identification of a novel hypovirulence-inducing hypovirus from Alternaria Alternata. Front. Microbiol. 2019, 10, 1076. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; Fu, Y.; Li, G.; Wang, M.; Jin, H.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. USA 2016, 113, 12803–12808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Cheng, S.; Chen, T.; Li, B.; Wang, Q.; et al. A 2-Kb mycovirus converts a pathogenic fungus into a beneficial endophyte for brassica protection and yield enhancement. Mol. Plant 2020, 13, 1420–1433. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, Y.; Lu, R.; Lau, N.; Lai, E.C.; Li, W.-X.; Ding, S.-W. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 1606–1611. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Ding, S.-W.; Zhang, Y.; Zhu, S. Identification of viruses and viroids by next-generation sequencing and homology-dependent and homology-independent algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; Saracchi, M.; Farina, G. Morphological, physiological and genetic diversity within a small population of Cercospora Beticola Sacc. Ann. Microbiol. 2004, 54, 129–150. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Wang, Q.; Cheng, S.; Xiao, X.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Discovery of two mycoviruses by high-throughput sequencing and assembly of mycovirus-derived small silencing RNAs from a hypovirulent strain of Sclerotinia sclerotiorum. Front. Microbiol. 2019, 10, 1415. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Y.; Wan, X.; She, Y.; Li, M.; Xi, H.; Xie, J.; Wen, C. A novel ourmia-like mycovirus confers hypovirulence-associated traits on Fusarium oxysporum. Front. Microbiol. 2020, 11, 569869. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Sang, W.; Wu, M.-D.; Zhang, J.; Yang, L.; Zhou, Y.-J.; Chen, W.-D.; Li, G.-Q. Novel hypovirulence-associated RNA mycovirus in the plant-pathogenic fungus Botrytis cinerea: Molecular and biological characterization. Appl. Environ. Microbiol. 2015, 81, 2299–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Thekke-Veetil, T.; Lagos-Kutz, D.; McCoppin, N.K.; Hartman, G.L.; Ju, H.-K.; Lim, H.-S.; Domier, L.L. Soybean thrips (Thysanoptera: Thripidae) harbor highly diverse populations of arthropod, fungal and plant viruses. Viruses 2020, 12, 1376. [Google Scholar] [CrossRef]
- McHale, M.T.; Roberts, I.N.; Noble, S.M.; Beaumont, C.; Whitehead, M.P.; Seth, D.; Oliver, R.P. CfT-I: An LTR-retrotransposon in Cladosporium Fulvum, a fungal pathogen of tomato. Mol. Genet. Genom. 1992, 233, 337–347. [Google Scholar] [CrossRef]
- Suzuki, N.; Ghabrial, S.A.; Kim, K.-H.; Pearson, M.; Marzano, S.-Y.L.; Yaegashi, H.; Xie, J.; Guo, L.; Kondo, H.; Koloniuk, I.; et al. ICTV virus taxonomy profile: Hypoviridae. J. Gen. Virol. 2018, 99, 615–616. [Google Scholar] [CrossRef]
- Romon-Ochoa, P.; Gorton, C.; Lewis, A.; van der Linde, S.; Webber, J.; Pérez-Sierra, A. Hypovirulent effect of the Cryphonectria hypovirus 1 in british isolates of Cryphonectria parasitica. Pest Manag. Sci. 2020, 76, 1333–1343. [Google Scholar] [CrossRef]
- Bryner, S.F.; Rigling, D.; Brunner, P.C. Invasion history and demographic pattern of Cryphonectria hypovirus 1 across European populations of the chestnut blight fungus. Ecol. Evol. 2012, 2, 3227–3241. [Google Scholar] [CrossRef]
- Chiba, S.; Suzuki, N. Highly activated RNA silencing via strong induction of dicer by one virus can interfere with the replication of an unrelated virus. Proc. Natl. Acad. Sci. USA 2015, 112, E4911–E4918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eusebio-Cope, A.; Sun, L.; Tanaka, T.; Chiba, S.; Kasahara, S.; Suzuki, N. The Chestnut blight fungus for studies on virus/host and virus/virus interactions: From a natural to a model host. Virology 2015, 477, 164–175. [Google Scholar] [CrossRef] [Green Version]
- García-Cuéllar, M.P.; Esteban, R.; Fujimura, T. RNA-dependent RNA polymerase activity associated with the yeast viral P91/20S RNA ribonucleoprotein complex. RNA 1997, 3, 27–36. [Google Scholar] [PubMed]
- Fujimura, T.; Esteban, R. Interactions of the RNA polymerase with the viral genome at the 5′- and 3′-ends contribute to 20S RNA narnavirus persistence in yeast. J. Biol. Chem. 2007, 282, 19011–19019. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Fishel, R.; Wickner, R.B. Circular single-stranded RNA replicon in Saccharomyces Cerevisiae. Proc. Natl. Acad. Sci. USA 1990, 87, 7628–7632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Yuan, Y.; Mao, J.; Yang, Z.; Cao, Q.; Zhang, T.; Wang, S.; Liu, D. Characterization of two novel mycoviruses from Penicillium digitatum and the related fungicide resistance analysis. Sci. Rep. 2018, 8, 5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS One 2018, 13, e0200511. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Myers, K.; Fry, W.E.; Hillman, B.I. A member of the virus family Narnaviridae from the plant pathogenic oomycete Phytophthora infestans. Arch. Virol. 2012, 157, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Chiba, Y.; Oiki, S.; Yaguchi, T.; Urayama, S.-I.; Hagiwara, D. Discovery of Divided RdRp Sequences and a Hitherto Unknown Genomic Complexity in Fungal Viruses. Virus Evol. 2021, 7, veaa101. [Google Scholar] [CrossRef]
- Sutela, S.; Forgia, M.; Vainio, E.J.; Chiapello, M.; Daghino, S.; Vallino, M.; Martino, E.; Girlanda, M.; Perotto, S.; Turina, M. The virome from a collection of endomycorrhizal fungi reveals new viral taxa with unprecedented genome organization. Virus Evol. 2020, 6, veaa076. [Google Scholar] [CrossRef]
- Jia, J.; Fu, Y.; Jiang, D.; Mu, F.; Cheng, J.; Lin, Y.; Li, B.; Marzano, S.-Y.L.; Xie, J. Interannual dynamics, diversity and evolution of the virome in Sclerotinia sclerotiorum from a single crop field. Virus Evol. 2021, 7, veab032. [Google Scholar] [CrossRef]
- Ruiz-Padilla, A.; Rodríguez-Romero, J.; Gómez-Cid, I.; Pacifico, D.; Ayllón, M.A. Novel mycoviruses discovered in the mycovirome of a necrotrophic fungus. mBio 2021, 12, e03705-20. [Google Scholar] [CrossRef]
- Ayllón, M.A.; Turina, M.; Xie, J.; Nerva, L.; Marzano, S.-Y.L.; Donaire, L.; Jiang, D.; ICTV Report Consortium. ICTV virus taxonomy profile: Botourmiaviridae. J. Gen. Virol. 2020, 101, 454–455. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in Esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef]
- Wang, Q.; Mu, F.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. A single ssRNA segment encoding RdRp is sufficient for replication, infection, and transmission of ourmia-like virus in fungi. Front. Microbiol. 2020, 11, 379. [Google Scholar] [CrossRef] [Green Version]
- Marzano, S.-Y.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, L.; Esmael, A.; Duan, J.; Bian, X.; Jia, J.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D.; et al. Four novel botourmiaviruses co-infecting an isolate of the rice blast fungus Magnaporthe oryzae. Viruses 2020, 12, 1383. [Google Scholar] [CrossRef] [PubMed]
- Donaire, L.; Rozas, J.; Ayllón, M.A. Molecular characterization of Botrytis ourmia-like virus, a mycovirus close to the plant pathogenic genus Ourmiavirus. Virology 2016, 489, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Chiba, S.; Toyoda, K.; Suzuki, N. Evidence for negative-strand RNA virus infection in fungi. Virology 2013, 435, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Wu, M.; Li, G. Molecular Characterization and geographic distribution of a mymonavirus in the population of Botrytis cinerea. Viruses 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xie, J.; Cheng, J.; Fu, Y.; Li, G.; Yi, X.; Jiang, D. Fungal negative-stranded RNA virus that is related to bornaviruses and nyaviruses. Proc. Natl. Acad. Sci. USA 2014, 111, 12205–12210. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; He, H.; Wang, S.; Chen, X.; Qiu, D.; Kondo, H.; Guo, L. Evidence for a novel negative-stranded RNA mycovirus isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2018, 518, 232–240. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. (Eds.) Family-Bunyaviridae. In Virus Taxonomy; Elsevier: San Diego, CA, USA, 2012; pp. 725–741. [Google Scholar]
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Bào, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.-X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Jiāng, D.-H.; Ayllón, M.A.; Marzano, S.-Y.L.; ICTV Report Consortium. ICTV virus taxonomy profile: Mymonaviridae. J. Gen. Virol. 2019, 100, 1343–1344. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wu, J.; Jiang, D.; Xie, J.; Cheng, J.; Lin, Y. ORF Ι of mycovirus SsNSRV-1 is associated with debilitating symptoms of Sclerotinia sclerotiorum. Viruses 2020, 12, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorens, C.; Soriano, B.; Krupovic, M.; Ictv Report Consortium. ICTV virus taxonomy profile: Metaviridae. J. Gen. Virol. 2020, 101, 1131–1132. [Google Scholar] [CrossRef] [PubMed]
- Sandmeyer, S.; Patterson, K.; Bilanchone, V. Ty3, a position-specific retrotransposon in budding yeast. Microbiol. Spectr. 2015, 3, 965–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef] [Green Version]
- Barba, M.; Czosnek, H.; Hadidi, A. Historical perspective, development and applications of next-generation sequencing in plant virology. Viruses 2014, 6, 106–136. [Google Scholar] [CrossRef] [PubMed]
- Kwok, K.T.T.; Nieuwenhuijse, D.F.; Phan, M.V.T.; Koopmans, M.P.G. Virus metagenomics in farm animals: A systematic review. Viruses 2020, 12, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]


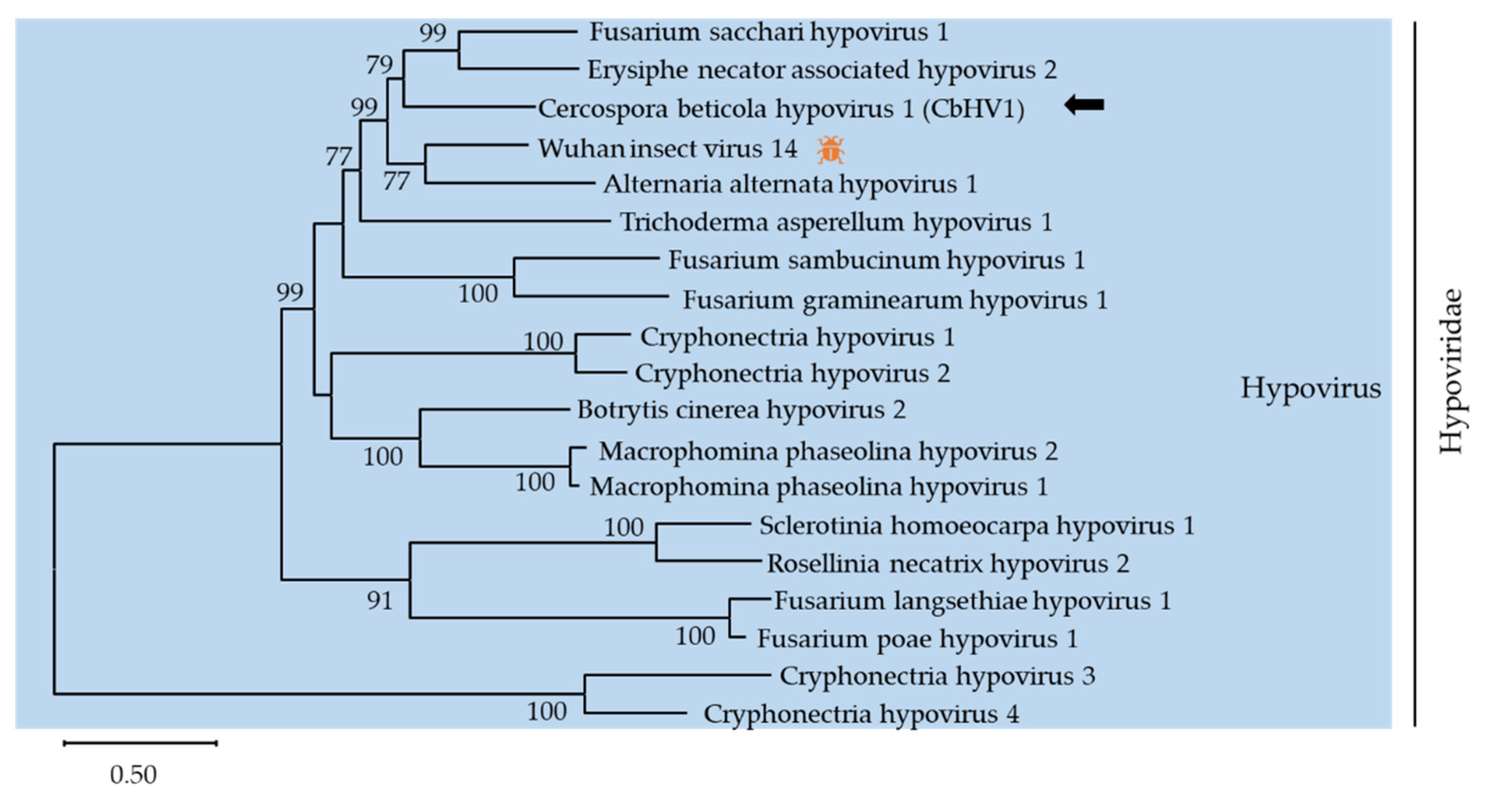
 ) represents viruses which can infect insects. Virus identified in this work is indicated by black arrows.
) represents viruses which can infect insects. Virus identified in this work is indicated by black arrows.
) represents viruses which can infect insects. Virus identified in this work is indicated by black arrows.
) represents viruses which can infect insects. Virus identified in this work is indicated by black arrows.

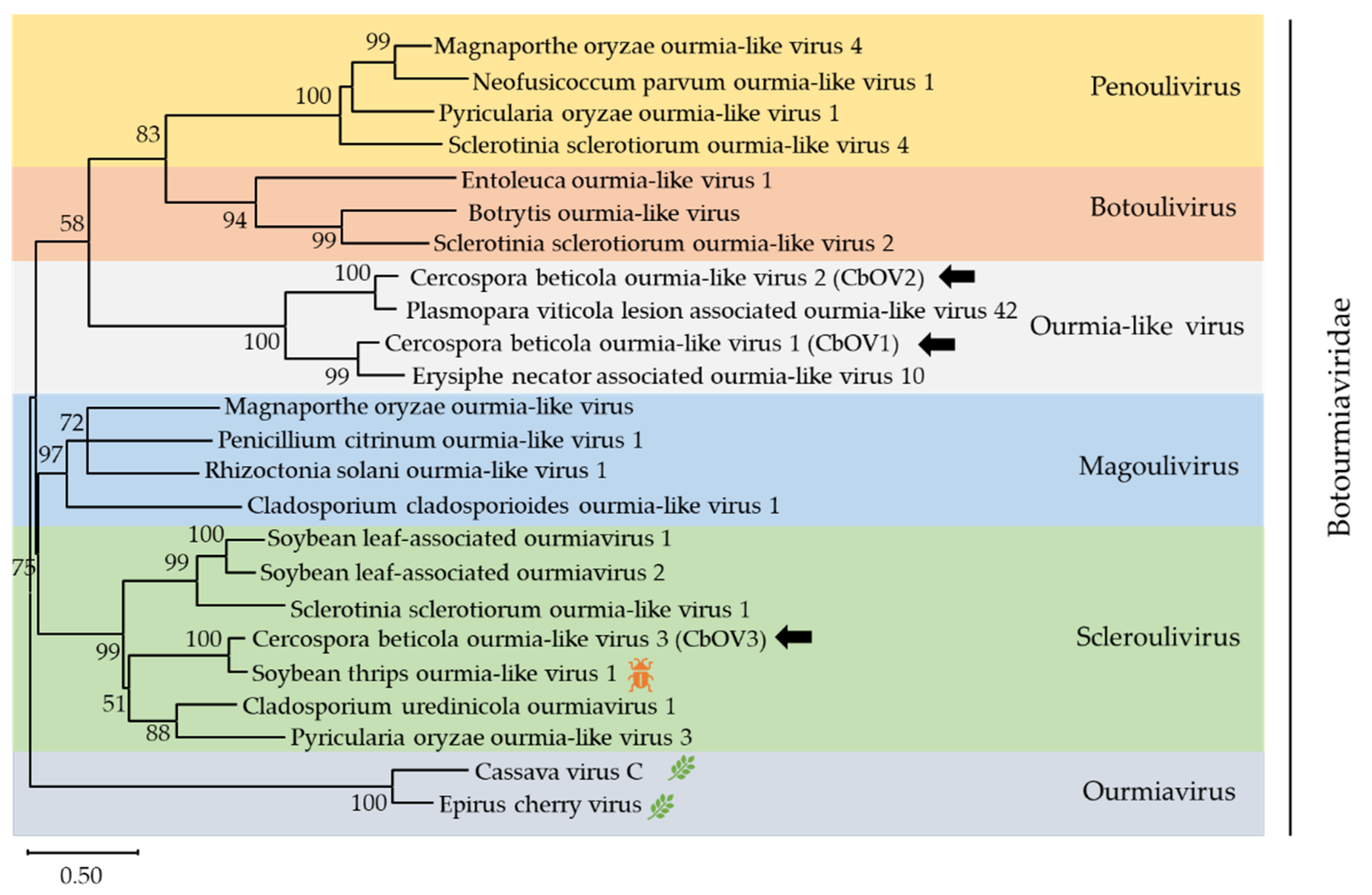
 ) represents viruses which can infect insects, and the leaf pattern (
) represents viruses which can infect insects, and the leaf pattern (  ) represents viruses which can infect plants. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect insects, and the leaf pattern ( ) represents viruses which can infect plants. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect plants. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect insects, and the leaf pattern ( ) represents viruses which can infect plants. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
 ) represents viruses which can infect insects. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect insects. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect insects. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.
) represents viruses which can infect insects. Only bootstrap values above 50% are indicated. Viruses identified in this work are indicated by black arrows.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Genome Type 1 | Family/Genus | Contig Number | Genbank Accession Number | Contig Length | Name of Putative Viruses | Best Match | aa Identity (%) | Query Coverage (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | (+)ssRNA | Hypoviridae | contig1357 | MZ546195 | 12624 | Cercospora beticola hypovirus 1 (CbHV1) | polyprotein [Wuhan insect virus 14] | 54.0 | 81 | [27] |
| 2 | (+)ssRNA | Narnaviridae | k141-72534 | MZ546196 | 2487 | Cercospora beticola narnavirus 1 (CbNV1) | RdRp [Erysiphe necator associated narnavirus 13] | 75.2 | 98 | Unpublished |
| 3 | (+)ssRNA | Botourmiaviridae | contig965 | MZ568927 | 2273 | Cercospora beticola ourmia-like virus 1 (CbOV1) | RdRp [Erysiphe necator associated ourmia-like virus 10] | 68.6 | 76 | Unpublished |
| 4 | (+)ssRNA | Botourmiaviridae | contig1024 | MZ568928 | 1440 | Cercospora beticola ourmia-like virus 2 (CbOV2) | RdRp [Plasmopara viticola lesion-associated ourmia-like virus 42] | 81.2 | 86 | [12] |
| 5 | (+)ssRNA | Botourmiaviridae | k141-5161 | MZ568929 | 2108 | Cercospora beticola ourmia-like virus 3 (CbOV3) | RdRp [Soybean thrips ourmia-like virus 1] | 83.4 | 82 | [28] |
| 6 | (−)ssRNA | - | contig154 | MZ599586 | 6604 | Cercospora beticola negative-stranded virus 1 (CbNSV1) | RdRp [Aspergillus fumigatus negative-stranded RNA virus 1] | 57.7 | 96 | Unpublished |
| 7 | (−)ssRNA | - | k141-54286 | MZ599593 | 566 | - | NS1 [Penicillium discovirus] | 49.2 | 63 | Unpublished |
| 8 | (−)ssRNA | - | k141-3378 | MZ599588 | 6191 | Cercospora beticola negative-stranded virus 3 (CbNSV3) | RdRp [Plasmopara viticola lesion-associated mononegaambi virus 5] | 62.8 | 94 | [12] |
| 9 | (−)ssRNA | - | k141-6617 | MZ599589 | 4197 | Cercospora beticola associated mycobunyavirales-like virus 1 segment L (CbBYV1) | RdRp [Plasmopara viticola lesion-associated mycobunyavirales-like virus 4] | 69.4 | 26 | [12] |
| 10 | (−)ssRNA | - | k141-67601 | MZ599592 | 1854 | Cercospora beticola associated mycobunyavirales-like virus 1 segment S (CbBYV1) | NS [Plasmopara viticola lesion-associated mycobunyavirales-like virus 4] | 46.0 | 76 | [12] |
| 11 | (−)ssRNA | - | k141-47347 | MZ599591 | 855 | Cercospora beticola associated mycobunyavirales-like virus 1 segment S (CbBYV1) | NP [Plasmopara viticola lesion-associated mycobunyavirales-like virus 4] | 65.9 | 92 | [12] |
| 12 | (−)ssRNA | - | k141-63059 | MZ599587 | 6457 | Cercospora beticola associated mycobunyavirales-like virus 2 (CbBYV2) | RdRp [Erysiphe necator associated negative-stranded RNA virus 10] | 63.2 | 84 | [12] |
| 13 | ssRNA-RT | Metaviridae | k141-31340 | MZ599594 | 1247 | - | env homologue [Cladosporium fulvum T-1 virus] | 64.3 | 33 | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhou, M.; Yang, Y.; Liu, Q.; Zhang, Z.; Han, C.; Wang, Y. Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola. Viruses 2021, 13, 1915. https://doi.org/10.3390/v13101915
Li Y, Zhou M, Yang Y, Liu Q, Zhang Z, Han C, Wang Y. Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola. Viruses. 2021; 13(10):1915. https://doi.org/10.3390/v13101915
Chicago/Turabian StyleLi, Yingxi, Mengke Zhou, Yizhou Yang, Qi Liu, Zongying Zhang, Chenggui Han, and Ying Wang. 2021. "Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola" Viruses 13, no. 10: 1915. https://doi.org/10.3390/v13101915
APA StyleLi, Y., Zhou, M., Yang, Y., Liu, Q., Zhang, Z., Han, C., & Wang, Y. (2021). Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola. Viruses, 13(10), 1915. https://doi.org/10.3390/v13101915