
Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Diagnosis of CMV Infections
2.2. Flow Cytometry
2.3. Unsupervised t-SNE Clustering
2.4. TCR Sequencing
2.5. Statistical Analyses
3. Results
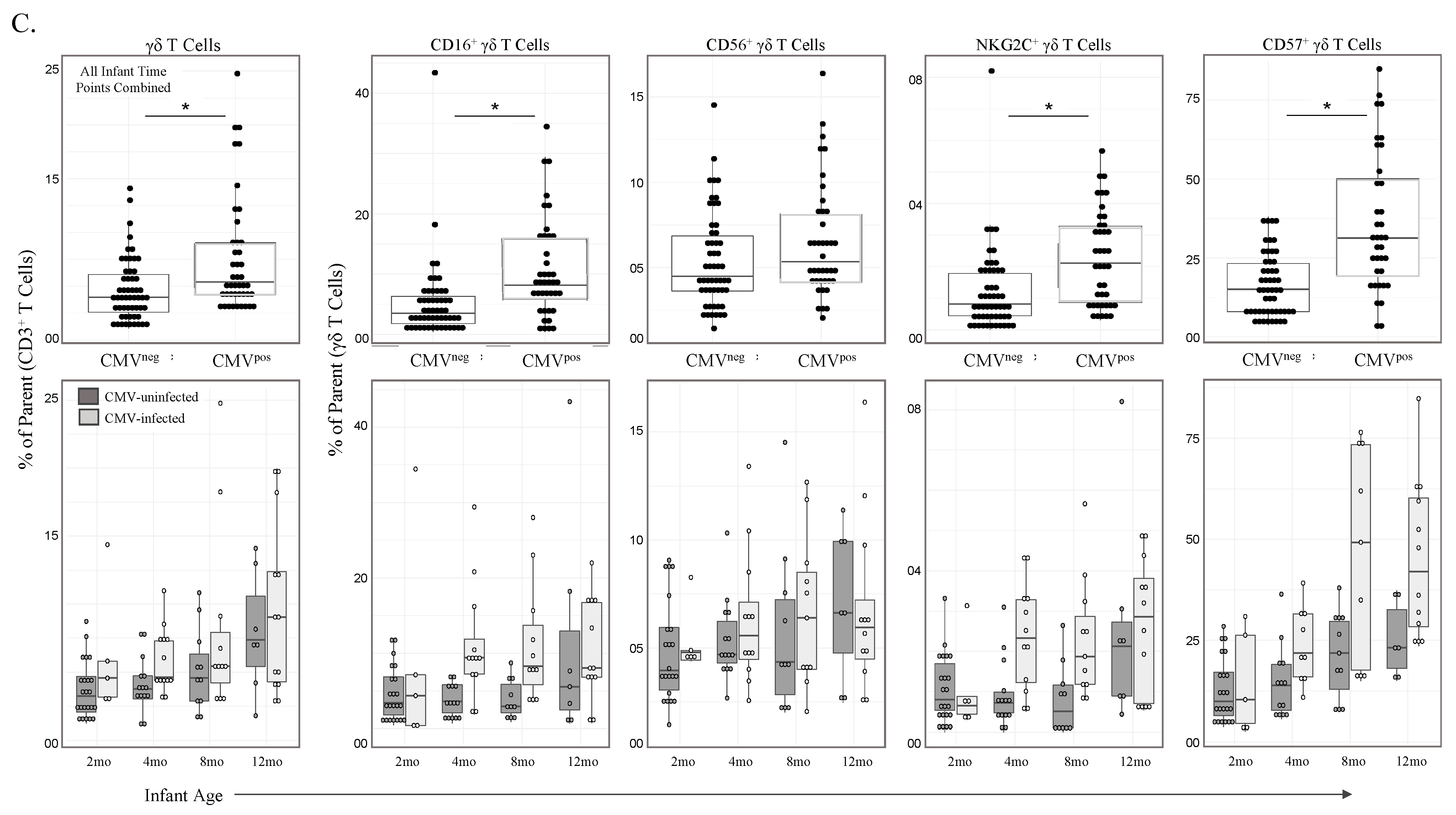
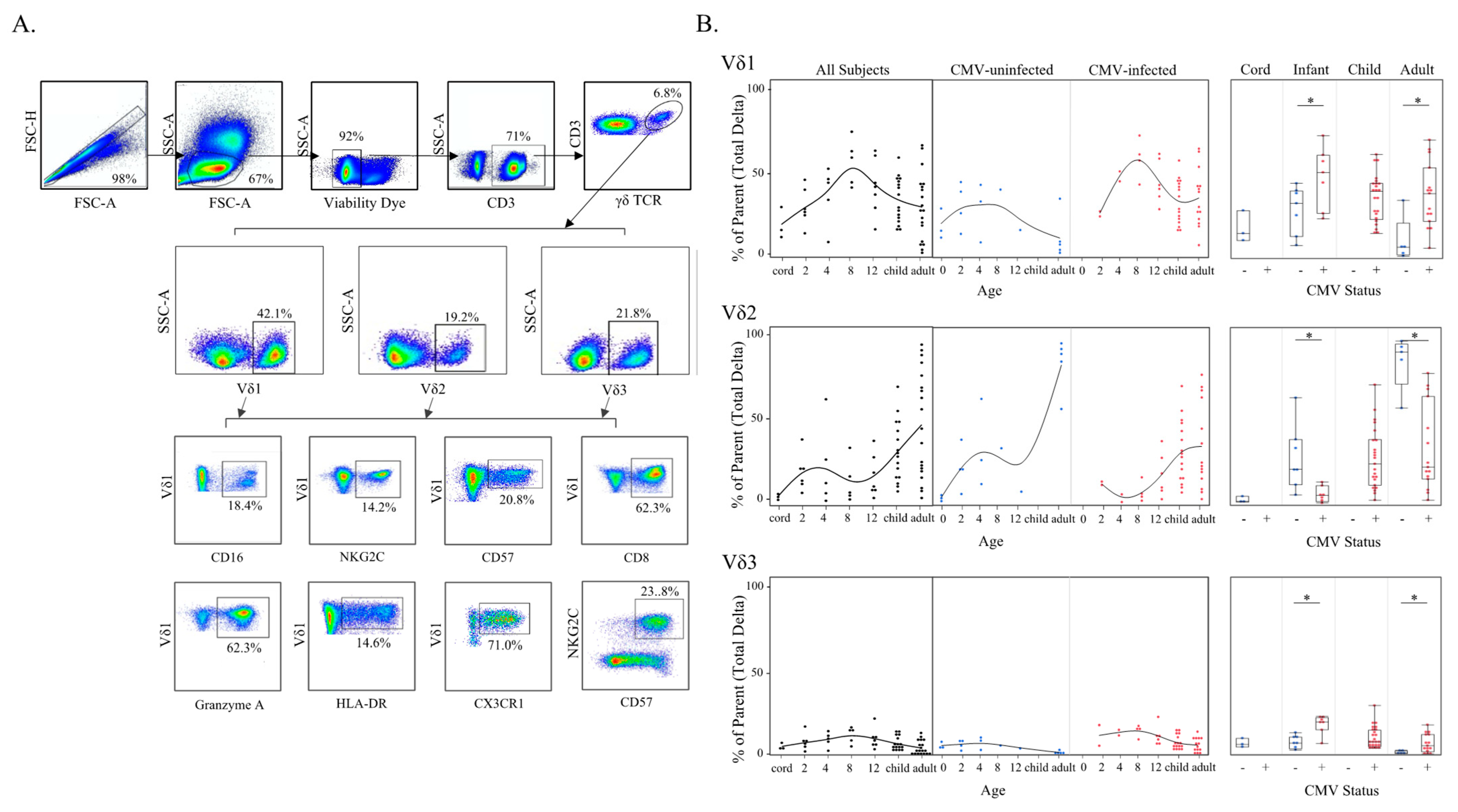
3.1. γδ T Cell Frequencies Change with Age and Are Expanded in Infants Experiencing Primary CMV Infection
3.2. A Public Vδ1-CALGELGDDKLIF TCR Clonotype Can Be Detected in Post-Natal CMV Infections Occuring Very Early in Life and Rapidly Decays
3.3. Adaptive-like Vδ1 and Vδ3 T Cells Have Similar Trajectories with Age and CMV Infection That Differ from the Innate-like Vδ2 T Cell Subset
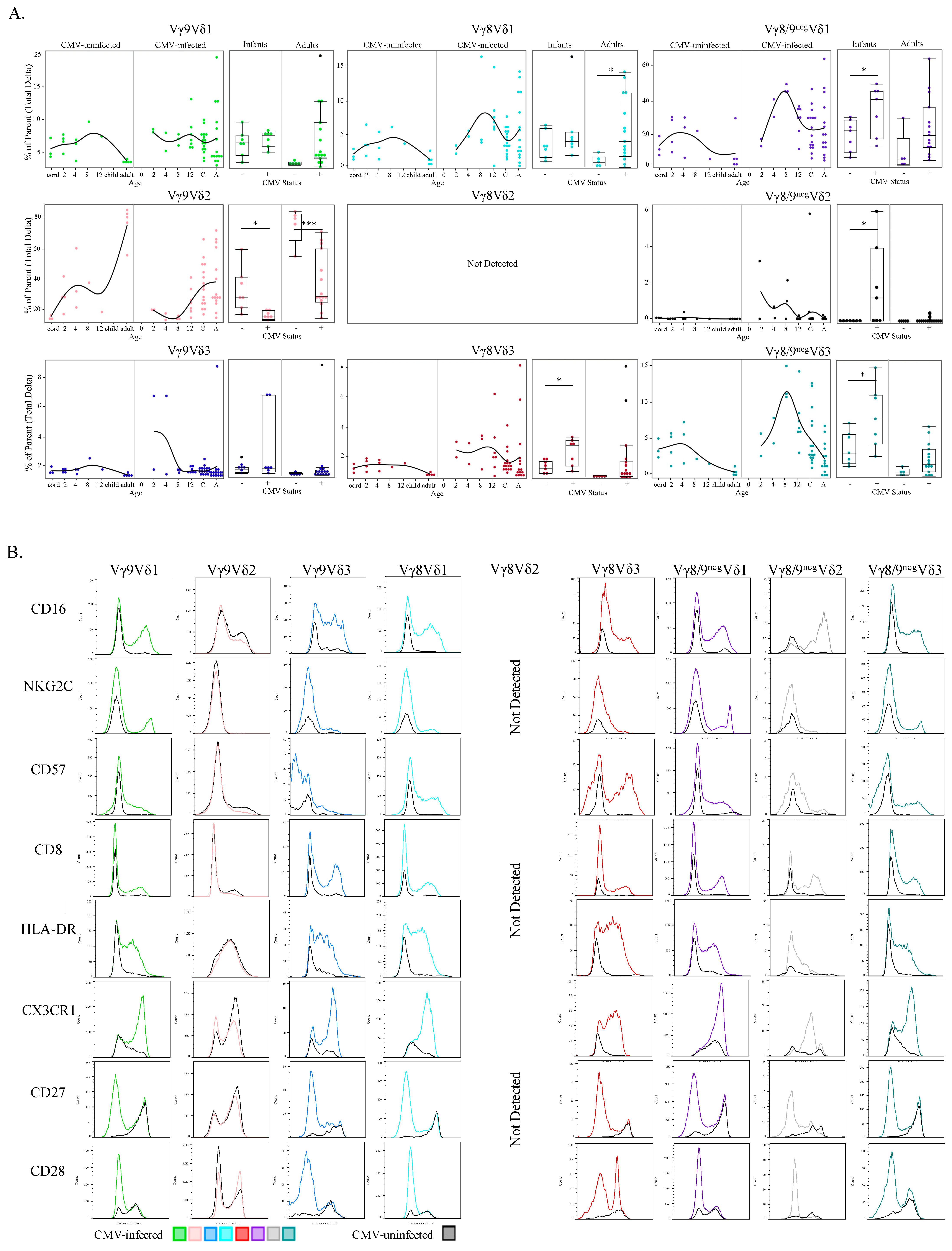
3.4. Phenotypic Differences of γδ T Cell δ-Subsets between CMV-Infected and CMV-Uninfected Infants and Adults
3.5. CMV-Associated γδ T Cell Subsets Have Unique Activated and Effector Phenotypes
3.6. NKG2C+CD57+ γδ T Cell Frequency Is Associated with NKG2C Genotype and CMV Shedding In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of Cytomegalovirus Seroprevalence and Demographic Characteristics Associated with Infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef]
- Savva, G.M.; Pachnio, A.; Kaul, B.; Morgan, K.; Huppert, F.A.; Brayne, C.; Moss, P.A.H. The Medical Research Council Cognitive Function and Ageing Study Cytomegalovirus Infection Is Associated with Increased Mortality in the Older Population. Aging Cell 2013, 12, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Pass, R.F.; Fowler, K.B.; Boppana, S.B.; Britt, W.J.; Stagno, S. Congenital Cytomegalovirus Infection Following First Trimester Maternal Infection: Symptoms at Birth and Outcome. J. Clin. Virol. 2006, 35, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.J.; Stowell, J.D.; Clark, R.; Dollard, P.R.; Johnson, D.; Mask, K.; Stover, C.; Wu, K.; Amin, M.; Hendley, W.; et al. Repeated Measures Study of Weekly and Daily Cytomegalovirus Shedding Patterns in Saliva and Urine of Healthy Cytomegalovirus-Seropositive Children. BMC Infect. Dis. 2014, 14, 569. [Google Scholar] [CrossRef]
- Gantt, S.; Marchant, A.; Boppana, S.B. Higher Expectations for a Vaccine to Prevent Congenital Cytomegalovirus Infection. J. Virol. 2018, 92, e00764-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, S.P. Prevention of Maternal–Fetal Transmission of Cytomegalovirus. EBioMedicine 2015, 2, 102–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, B.T.; Krantz, E.M.; Swan, D.; Ferrenberg, J.; Simmons, K.; Selke, S.; Huang, M.-L.; Casper, C.; Corey, L.; Wald, A.; et al. Transient Oral Human Cytomegalovirus Infections Indicate Inefficient Viral Spread from Very Few Initially Infected Cells. J. Virol. 2017, 91, e00380-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uldrich, A.P.; Le Nours, J.; Pellicci, D.G.; Gherardin, N.A.; McPherson, K.G.; Lim, R.T.; Patel, O.; Beddoe, T.; Gras, S.; Rossjohn, J.; et al. CD1d-Lipid Antigen Recognition by the Γδ TCR. Nat. Immunol. 2013, 14, 1137–1145. [Google Scholar] [CrossRef]
- Born, W.K.; Reardon, C.L.; O’Brien, R.L. The Function of Γδ T Cells in Innate Immunity. Curr. Opin. Immunol. 2006, 18, 31–38. [Google Scholar] [CrossRef]
- Vantourout, P.; Hayday, A. Six-of-the-Best: Unique Contributions of Γδ T Cells to Immunology. Nat. Rev. Immunol. 2013, 13, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Vermijlen, D.; Gatti, D.; Kouzeli, A.; Rus, T.; Eberl, M. Γδ T Cell Responses: How Many Ligands Will It Take till We Know? Semin. Cell Dev. Biol. 2018, 84, 75–86. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, N.E.; Eberl, M. Human Γδ T-Cell Control of Mucosal Immunity and Inflammation. Front. Immunol. 2018, 9, 985. [Google Scholar] [CrossRef]
- Hayday, A.C. Γδ T Cell Update: Adaptate Orchestrators of Immune Surveillance. J. Immunol. 2019, 203, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, Y.; Lau, Y.-L.; Tu, W. Γδ-T Cells: An Unpolished Sword in Human Anti-Infection Immunity. Cell. Mol. Immunol. 2013, 10, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Couzi, L.; Lafarge, X.; Pitard, V.; Neau-Cransac, M.; Dromer, C.; Billes, M.-A.; Lacaille, F.; Moreau, J.-F.; Merville, P.; Déchanet-Merville, J. Gamma-Delta T Cell Expansion Is Closely Associated with Cytomegalovirus Infection in All Solid Organ Transplant Recipients: Letter to the Editors. Transpl. Int. 2011, 24, e40–e42. [Google Scholar] [CrossRef]
- Déchanet, J.; Merville, P.; Lim, A.; Retière, C.; Pitard, V.; Lafarge, X.; Michelson, S.; Méric, C.; Hallet, M.-M.; Kourilsky, P.; et al. Implication of Γδ T Cells in the Human Immune Response to Cytomegalovirus. J. Clin. Investig. 1999, 103, 1437–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitard, V.; Roumanes, D.; Lafarge, X.; Couzi, L.; Garrigue, I.; Lafon, M.-E.; Merville, P.; Moreau, J.-F.; Dechanet-Merville, J. Long-Term Expansion of Effector/Memory V 2- T Cells Is a Specific Blood Signature of CMV Infection. Blood 2008, 112, 1317–1324. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.S.; Willcox, C.R.; Joyce, S.P.; Ladell, K.; Kasatskaya, S.A.; McLaren, J.E.; Hunter, S.; Salim, M.; Mohammed, F.; Price, D.A.; et al. Clonal Selection in the Human Vδ1 T Cell Repertoire Indicates Γδ TCR-Dependent Adaptive Immune Surveillance. Nat. Commun. 2017, 8, 14760. [Google Scholar] [CrossRef]
- Ravens, S.; Schultze-Florey, C.; Raha, S.; Sandrock, I.; Drenker, M.; Oberdörfer, L.; Reinhardt, A.; Ravens, I.; Beck, M.; Geffers, R.; et al. Human Γδ T Cells Are Quickly Reconstituted after Stem-Cell Transplantation and Show Adaptive Clonal Expansion in Response to Viral Infection. Nat. Immunol. 2017, 18, 393–401. [Google Scholar] [CrossRef]
- Vermijlen, D.; Brouwer, M.; Donner, C.; Liesnard, C.; Tackoen, M.; Van Rysselberge, M.; Twité, N.; Goldman, M.; Marchant, A.; Willems, F. Human Cytomegalovirus Elicits Fetal Γδ T Cell Responses in Utero. J. Exp. Med. 2010, 207, 807–821. [Google Scholar] [CrossRef] [Green Version]
- Mold, J.E.; Venkatasubrahmanyam, S.; Burt, T.D.; Michaelsson, J.; Rivera, J.M.; Galkina, S.A.; Weinberg, K.; Stoddart, C.A.; McCune, J.M. Fetal and Adult Hematopoietic Stem Cells Give Rise to Distinct T Cell Lineages in Humans. Science 2010, 330, 1695–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulou, M.; Sanchez Sanchez, G.; Vermijlen, D. Innate and Adaptive Γδ T Cells: How, When, and Why. Immunol. Rev. 2020, 298, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.S.; Willcox, C.R.; Hunter, S.; Kasatskaya, S.A.; Remmerswaal, E.B.M.; Salim, M.; Mohammed, F.; Bemelman, F.J.; Chudakov, D.M.; Oo, Y.H.; et al. The Human Vδ2+ T-Cell Compartment Comprises Distinct Innate-like Vγ9+ and Adaptive Vγ9- Subsets. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Monaco, G.; Wong, E.H.; Tan, W.L.W.; Kared, H.; Simoni, Y.; Tan, S.W.; How, W.Z.Y.; Tan, C.T.Y.; Lee, B.T.K.; et al. Mapping of γ/δ T Cells Reveals Vδ2+ T Cells Resistance to Senescence. EBioMedicine 2019, 39, 44–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieppo, P.; Papadopoulou, M.; Gatti, D.; McGovern, N.; Chan, J.K.Y.; Gosselin, F.; Goetgeluk, G.; Weening, K.; Ma, L.; Dauby, N.; et al. The Human Fetal Thymus Generates Invariant Effector Γδ T Cells. J. Exp. Med. 2020, 217, e20190580. [Google Scholar] [CrossRef]
- Gantt, S.; Orem, J.; Krantz, E.M.; Morrow, R.A.; Selke, S.; Huang, M.-L.; Schiffer, J.T.; Jerome, K.R.; Nakaganda, A.; Wald, A.; et al. Prospective Characterization of the Risk Factors for Transmission and Symptoms of Primary Human Herpesvirus Infections Among Ugandan Infants. J. Infect. Dis. 2016, 214, 36–44. [Google Scholar] [CrossRef]
- Johnston, C.; Orem, J.; Okuku, F.; Kalinaki, M.; Saracino, M.; Katongole-Mbidde, E.; Sande, M.; Ronald, A.; McAdam, K.; Huang, M.-L.; et al. Impact of HIV Infection and Kaposi Sarcoma on Human Herpesvirus-8 Mucosal Replication and Dissemination in Uganda. PLoS ONE 2009, 4, e4222. [Google Scholar] [CrossRef]
- Papadopoulou, M.; Tieppo, P.; McGovern, N.; Gosselin, F.; Chan, J.K.Y.; Goetgeluk, G.; Dauby, N.; Cogan, A.; Donner, C.; Ginhoux, F.; et al. TCR Sequencing Reveals the Distinct Development of Fetal and Adult Human Vγ9Vδ2 T Cells. J. Immunol. 2019, 203, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, S.C.; Andrus, J.P.; Perfetto, S.P.; Mantovani, J.J.; Herzenberg, L.A.; Herzenberg, L.A.; Roederer, M. Ontogeny of Γδ T Cells in Humans. J. Immunol. 2004, 172, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulou, M.; Dimova, T.; Shey, M.; Briel, L.; Veldtsman, H.; Khomba, N.; Africa, H.; Steyn, M.; Hanekom, W.A.; Scriba, T.J.; et al. Fetal Public Vγ9Vδ2 T Cells Expand and Gain Potent Cytotoxic Functions Early after Birth. Proc. Natl. Acad. Sci. USA 2020, 117, 18638–18648. [Google Scholar] [CrossRef]
- Ravens, S.; Fichtner, A.S.; Willers, M.; Torkornoo, D.; Pirr, S.; Schöning, J.; Deseke, M.; Sandrock, I.; Bubke, A.; Wilharm, A.; et al. Microbial Exposure Drives Polyclonal Expansion of Innate Γδ T Cells Immediately after Birth. Proc. Natl. Acad. Sci. USA 2020, 117, 18649–18660. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.; Björkander, S.; Qazi, K.R.; Bittmann, J.; Hell, L.; Jenmalm, M.C.; Marchini, G.; Vermijlen, D.; Abrahamsson, T.; Nilsson, C.; et al. Characterization of the Γδ T-cell Compartment during Infancy Reveals Clear Differences between the Early Neonatal Period and 2 Years of Age. Immunol. Cell Biol. 2019, 98, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Couzi, L.; Pitard, V.; Sicard, X.; Garrigue, I.; Hawchar, O.; Merville, P.; Moreau, J.-F.; Dechanet-Merville, J. Antibody-Dependent Anti-Cytomegalovirus Activity of Human T Cells Expressing CD16 (Fc RIIIa). Blood 2012, 119, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Gaballa, A.; Arruda, L.C.M.; Rådestad, E.; Uhlin, M. CD8 + Γδ T Cells Are More Frequent in CMV Seropositive Bone Marrow Grafts and Display Phenotype of an Adaptive Immune Response. Stem Cells Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadivar, M.; Petersson, J.; Svensson, L.; Marsal, J. CD8αβ + Γδ T Cells: A Novel T Cell Subset with a Potential Role in Inflammatory Bowel Disease. J. Immunol. 2016, 197, 4584–4592. [Google Scholar] [CrossRef] [Green Version]
- Scheper, W.; van Dorp, S.; Kersting, S.; Pietersma, F.; Lindemans, C.; Hol, S.; Heijhuurs, S.; Sebestyen, Z.; Gründer, C.; Marcu-Malina, V.; et al. ΓδT Cells Elicited by CMV Reactivation after Allo-SCT Cross-Recognize CMV and Leukemia. Leukemia 2013, 27, 1328–1338. [Google Scholar] [CrossRef]
- Lieberman, J. Granzyme A Activates Another Way to Die: Granzyme A-Mediated Cell Death. Immunol. Rev. 2010, 235, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Gumá, M.; Budt, M.; Sáez, A.; Brckalo, T.; Hengel, H.; Angulo, A.; López-Botet, M. Expansion of CD94/NKG2C+ NK Cells in Response to Human Cytomegalovirus-Infected Fibroblasts. Blood 2006, 107, 3624–3631. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Verges, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a Unique CD57+NKG2Chi Natural Killer Cell Subset during Acute Human Cytomegalovirus Infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vergès, S.; Milush, J.M.; Pandey, S.; York, V.A.; Arakawa-Hoyt, J.; Pircher, H.; Norris, P.J.; Nixon, D.F.; Lanier, L.L. CD57 Defines a Functionally Distinct Population of Mature NK Cells in the Human CD56dimCD16+ NK-Cell Subset. Blood 2010, 116, 3865–3874. [Google Scholar] [CrossRef] [Green Version]
- Newhook, N.; Fudge, N.; Grant, M. NK Cells Generate Memory-Type Responses to Human Cytomegalovirus-Infected Fibroblasts. Eur. J. Immunol. 2017, 47, 1032–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodier, M.R.; White, M.J.; Darboe, A.; Nielsen, C.M.; Goncalves, A.; Bottomley, C.; Moore, S.E.; Riley, E.M. Rapid NK Cell Differentiation in a Population with Near-Universal Human Cytomegalovirus Infection Is Attenuated by NKG2C Deletions. Blood 2014, 124, 2213–2222. [Google Scholar] [CrossRef] [Green Version]
- Moraru, M.; Cañizares, M.; Muntasell, A.; Pablo, R.; López-Botet, M.; Vilches, C. Assessment of Copy-Number Variation in the NKG2C Receptor Gene in a Single-Tube and Characterization of a Reference Cell Panel, Using Standard Polymerase Chain Reaction: NKG2C Copy-Number Variation—Genotyping and a Reference Panel. Tissue Antigens 2012, 80, 184–187. [Google Scholar] [CrossRef]
- Vietzen, H.; Pollak, K.; Honsig, C.; Jaksch, P.; Puchhammer-Stöckl, E. NKG2C Deletion Is a Risk Factor for Human Cytomegalovirus Viremia and Disease After Lung Transplantation. J. Infect. Dis. 2018, 217, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Lilleri, D.; Fornara, C.; Revello, M.G.; Gerna, G. Human Cytomegalovirus–Specific Memory CD8 + and CD4 + T Cell Differentiation after Primary Infection. J. Infect. Dis. 2008, 198, 536–543. [Google Scholar] [CrossRef]
- Klenerman, P.; Oxenius, A. T Cell Responses to Cytomegalovirus. Nat. Rev. Immunol. 2016, 16, 367–377. [Google Scholar] [CrossRef]
- Lidehall, A.K.; Sund, F.; Lundberg, T.; Eriksson, B.-M.; Tötterman, T.H.; Korsgren, O. T Cell Control of Primary and Latent Cytomegalovirus Infections in Healthy Subjects. J. Clin. Immunol. 2005, 25, 473–481. [Google Scholar] [CrossRef]
- Tomasec, P. Surface Expression of HLA-E, an Inhibitor of Natural Killer Cells, Enhanced by Human Cytomegalovirus GpUL40. Science 2000, 287, 1031–1033. [Google Scholar] [CrossRef] [PubMed]
- Prod’homme, V.; Tomasec, P.; Cunningham, C.; Lemberg, M.K.; Stanton, R.J.; McSharry, B.P.; Wang, E.C.Y.; Cuff, S.; Martoglio, B.; Davison, A.J.; et al. Human Cytomegalovirus UL40 Signal Peptide Regulates Cell Surface Expression of the NK Cell Ligands HLA-E and GpUL18. J. Immunol. 2012, 188, 2794–2804. [Google Scholar] [CrossRef]
- Hammer, Q.; Rückert, T.; Borst, E.M.; Dunst, J.; Haubner, A.; Durek, P.; Heinrich, F.; Gasparoni, G.; Babic, M.; Tomic, A.; et al. Peptide-Specific Recognition of Human Cytomegalovirus Strains Controls Adaptive Natural Killer Cells. Nat. Immunol. 2018, 19, 453–463. [Google Scholar] [CrossRef]
- Angelini, D.F.; Zambello, R.; Galandrini, R.; Diamantini, A.; Placido, R.; Micucci, F.; Poccia, F.; Semenzato, G.; Borsellino, G.; Santoni, A.; et al. NKG2A Inhibits NKG2C Effector Functions of Γδ T Cells: Implications in Health and Disease. J. Leukoc. Biol. 2011, 89, 75–84. [Google Scholar] [CrossRef]
- Fausther-Bovendo, H.; Wauquier, N.; Cherfils-Vicini, J.; Cremer, I.; Debre, P.; Vieillard, V. NKG2C Is a Major Triggering Receptor Involved in the Vd1 T Cell-Mediated Cytotoxicity against HIV-Infected CD4 T Cells. Aids 2008, 22, 217–226. [Google Scholar] [CrossRef]
- O’Sullivan, T.E.; Sun, J.C.; Lanier, L.L. Natural Killer Cell Memory. Immunity 2015, 43, 634–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumá, M.; Angulo, A.; Vilches, C.; Gómez-Lozano, N.; Malats, N.; López-Botet, M. Imprint of Human Cytomegalovirus Infection on the NK Cell Receptor Repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [Green Version]
- Hertoghs, K.M.L.; Moerland, P.D.; van Stijn, A.; Remmerswaal, E.B.M.; Yong, S.L.; van de Berg, P.J.E.J.; van Ham, S.M.; Baas, F.; ten Berge, I.J.M.; van Lier, R.A.W. Molecular Profiling of Cytomegalovirus-Induced Human CD8+ T Cell Differentiation. J. Clin. Investig. 2010, 120, 4077–4090. [Google Scholar] [CrossRef] [PubMed]
- Kallemeijn, M.J.; Boots, A.M.H.; van der Klift, M.Y.; Brouwer, E.; Abdulahad, W.H.; Verhaar, J.A.N.; van Dongen, J.J.M.; Langerak, A.W. Ageing and Latent CMV Infection Impact on Maturation, Differentiation and Exhaustion Profiles of T-Cell Receptor Gammadelta T-Cells. Sci. Rep. 2017, 7, 5509. [Google Scholar] [CrossRef]
- Verstichel, G.; Vermijlen, D.; Martens, L.; Goetgeluk, G.; Brouwer, M.; Thiault, N.; Caeneghem, Y.V.; Munter, S.D.; Weening, K.; Bonte, S.; et al. The Checkpoint for Agonist Selection Precedes Conventional Selection in Human Thymus. Sci. Immunol. 2017, 12, 860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbings, D.; Befus, A.D. CD4 and CD8: An inside-out Coreceptor Model for Innate Immune Cells. J. Leukoc. Biol. 2009, 86, 251–259. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMV-Infected Infants | CMV-Infected Adults | ||
|---|---|---|---|
| Expression levels compared to uninfected age controls | |||
| CD16 | Vδ1 | Higher (***) | Higher (*) |
| Vδ2 | Higher (*) | Lower (*) | |
| Vδ3 | Higher (***) | Higher (*) | |
| NKG2C | Vδ1 | Higher (*) | Higher (*) |
| Vδ2 | Not detected | Not detected | |
| Vδ3 | Higher (*) | No difference | |
| CD57 | Vδ1 | Higher (***) | No difference |
| Vδ2 | No difference | No difference | |
| Vδ3 | Higher (*) | Higher (*) | |
| CD8 | Vδ1 | Higher (*) | Higher (*) |
| Vδ2 | No difference | Lower (*) | |
| Vδ3 | Higher (*) | Higher (*) | |
| Granzyme A | Vδ1 | Higher (***) | Higher (*) |
| Vδ2 | No difference | No difference | |
| Vδ3 | Higher (***) | Higher (***) | |
| HLA-DR | Vδ1 | Higher (***) | Higher (*) |
| Vδ2 | Lower (*) | No difference | |
| Vδ3 | Higher (***) | Higher (***) | |
| CX3CR1 | Vδ1 | Higher (***) | No difference |
| Vδ2 | No difference | Lower (*) | |
| Vδ3 | Higher (***) | Higher (***) | |
| NKG2C+ CD57+ | Vδ1 | Higher (*) | Higher (*) |
| Not detected in CMV uninfected individuals | |||
| Vδ2 | Not detected | Not detected | |
| Vδ3 | No difference | Higher (***) | |
| Not detected in CMV uninfected individuals | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuengel, J.; Ranchal, S.; Maslova, A.; Aulakh, G.; Papadopoulou, M.; Drissler, S.; Cai, B.; Mohsenzadeh-Green, C.; Soudeyns, H.; Mostafavi, S.; et al. Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection. Viruses 2021, 13, 1987. https://doi.org/10.3390/v13101987
Tuengel J, Ranchal S, Maslova A, Aulakh G, Papadopoulou M, Drissler S, Cai B, Mohsenzadeh-Green C, Soudeyns H, Mostafavi S, et al. Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection. Viruses. 2021; 13(10):1987. https://doi.org/10.3390/v13101987
Chicago/Turabian StyleTuengel, Jessica, Sanya Ranchal, Alexandra Maslova, Gurpreet Aulakh, Maria Papadopoulou, Sibyl Drissler, Bing Cai, Cetare Mohsenzadeh-Green, Hugo Soudeyns, Sara Mostafavi, and et al. 2021. "Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection" Viruses 13, no. 10: 1987. https://doi.org/10.3390/v13101987
APA StyleTuengel, J., Ranchal, S., Maslova, A., Aulakh, G., Papadopoulou, M., Drissler, S., Cai, B., Mohsenzadeh-Green, C., Soudeyns, H., Mostafavi, S., van den Elzen, P., Vermijlen, D., Cook, L., & Gantt, S. (2021). Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection. Viruses, 13(10), 1987. https://doi.org/10.3390/v13101987