
Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patholomorphological, Histological and Ultrastructural Examination
2.2. Cells and Viruses
2.3. Detection of SWPV-Specific Sequences Using PCR and qPCR
2.4. Permissivity of Different Porcine Cell Lines to SWPV Infection
2.5. Whole Genome Sequencing and Analysis
2.6. Phylogenetic Analysis
3. Results
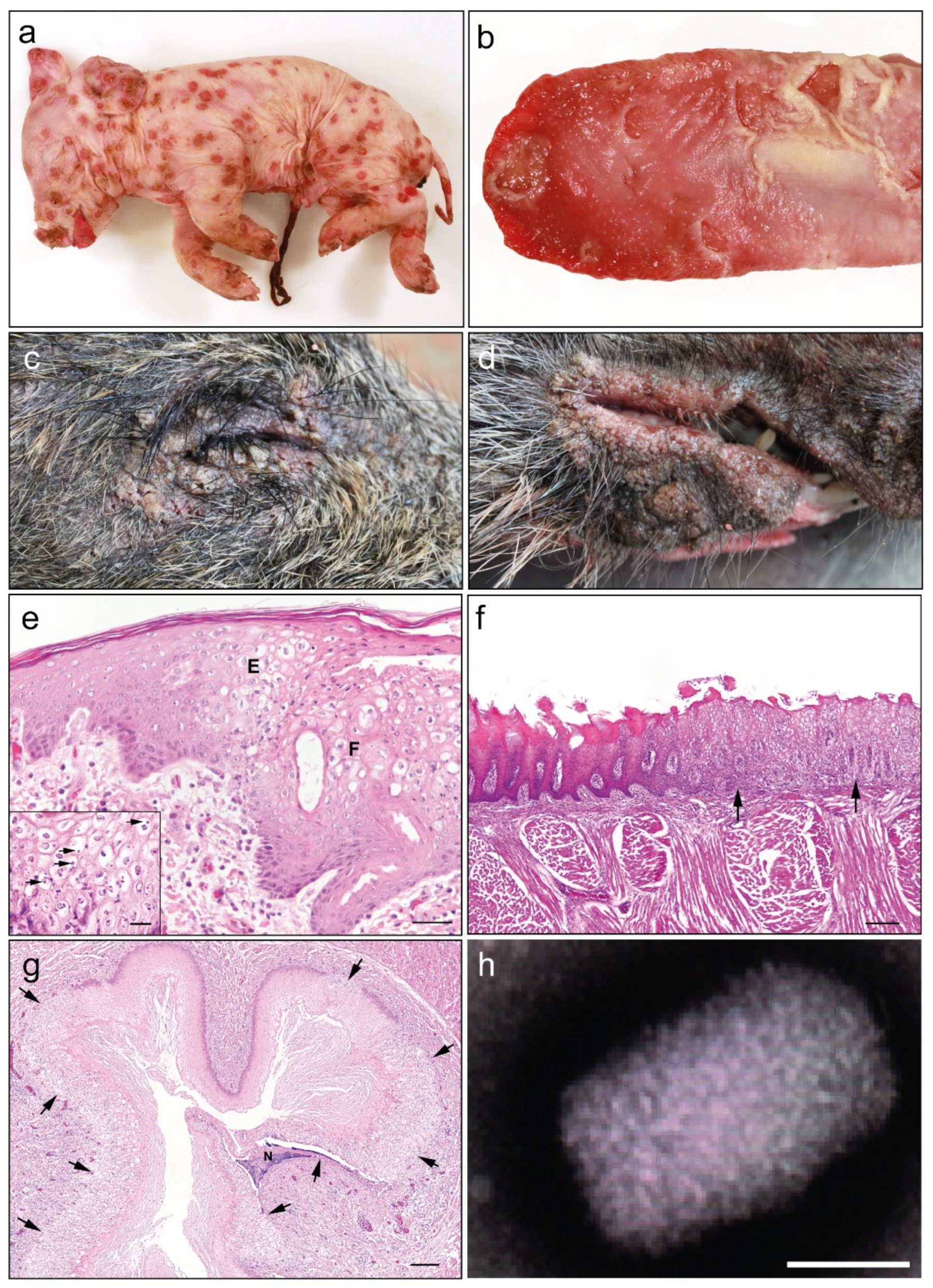
3.1. Clinical and Pathological Findings
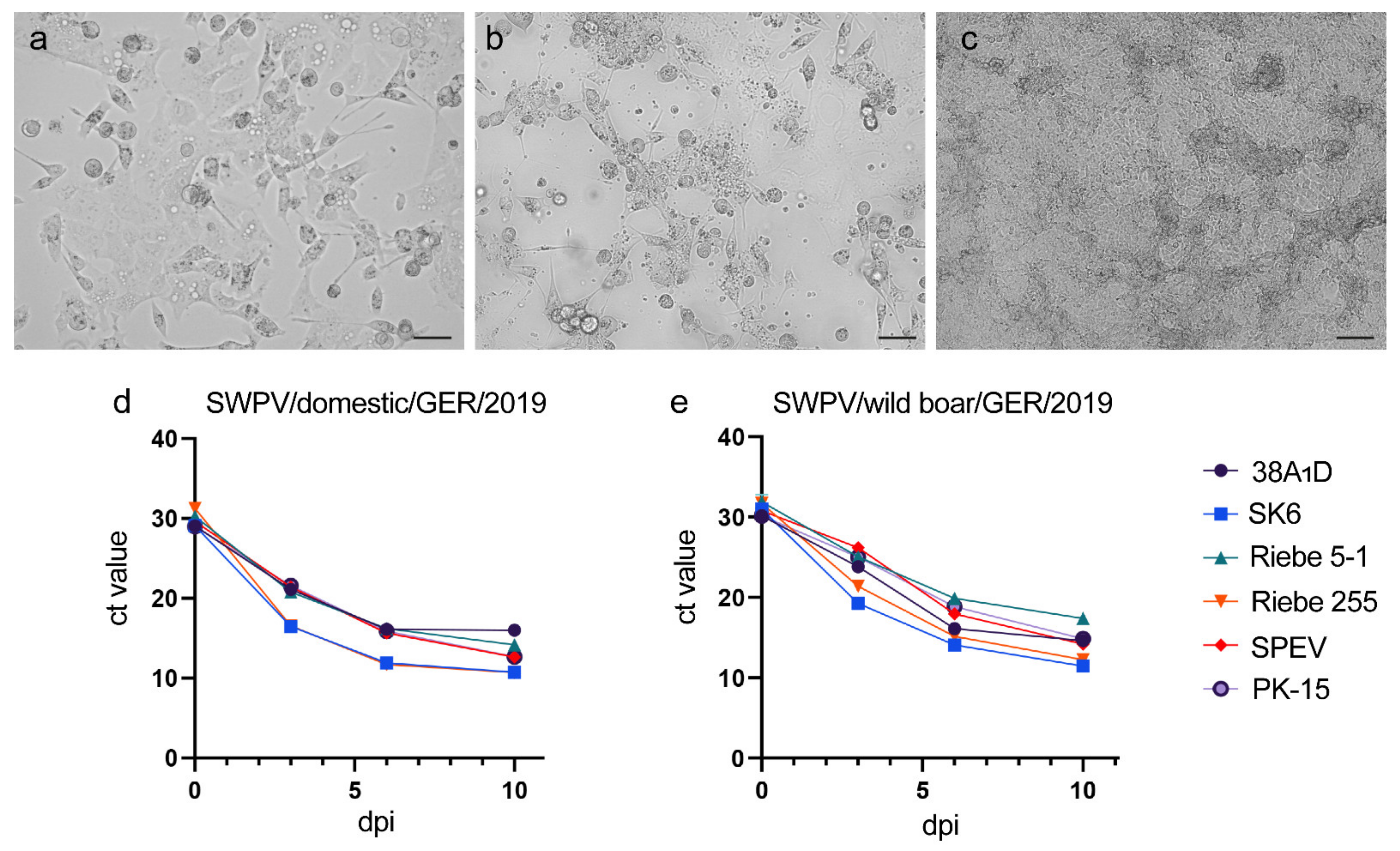
3.2. Assessment of The Susceptibility of Porcine Cell Lines to SWPV Infection
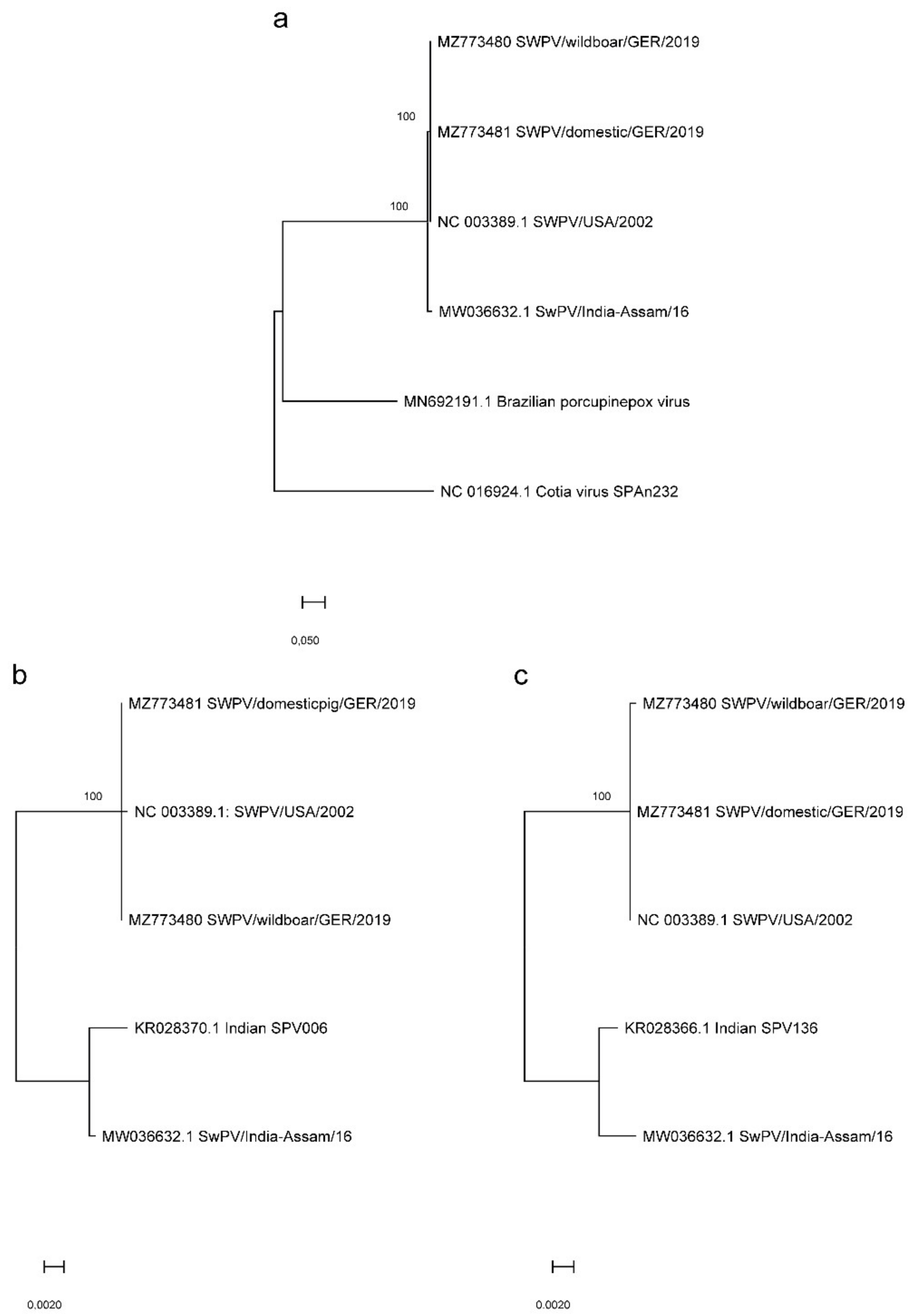
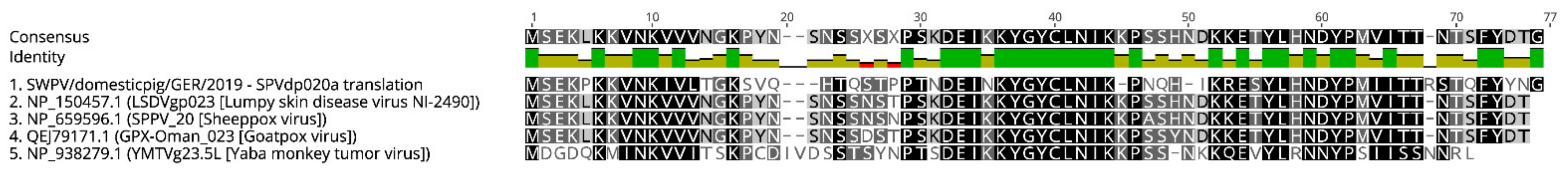
3.3. Phylogenetic and Whole Genome Analysis of SWPV Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spinola, W.T.J. Die Krankheiten der Schweine (1842); Hieschwald, A., Ed.; Kessinger Publishing: Berlin, Germany, 2010; p. 204. [Google Scholar]
- McNutt, S.; Murray, C.; Purwin, P. Swine Pox. J. Am. Vet. Med. Assoc. 1929, 74, 752–761. [Google Scholar]
- Olufemi, B.E.; Ayoade, G.O.; Ikede, B.O.; Akpavie, S.O.; Nwufoh, K.J. Swine pox in Nigeria. Vet. Rec. 1981, 109, 278–280. [Google Scholar] [CrossRef]
- Jubb, T.F.; Ellis, T.M.; Peet, R.L.; Parkinson, J. Swinepox in pigs in northern Western Australia. Aust. Vet. J. 1992, 69, 99. [Google Scholar] [CrossRef]
- Meyer, R.C.; Conroy, J.D. Experimental Swinepox in Gnotobiotic Piglets. Res. Vet. Sci. 1972, 13, 334–340. [Google Scholar] [CrossRef]
- Olinda, R.G.; Maia, L.A.; Cargnelutti, J.F.; Gois, R.C.S.; Batista, J.S.; Dantas, A.F.M.; Flores, E.F.; Riet-Correa, F. Swinepox dermatitis in backyard pigs in Northeastern Brazil. Pesqui. Vet. Bras. 2016, 36, 468–472. [Google Scholar] [CrossRef] [Green Version]
- Medaglia, M.L.G.; de Cássia Pereira, A.; Freitas, T.R.P.; Damaso, C.R. Swinepox virus outbreak, Brazil, 2011. Emerg. Infect. Dis. 2011, 17, 1976–1978. [Google Scholar] [CrossRef]
- Mech, P.; Bora, D.P.; Neher, S.; Barman, N.N.; Borah, P.; Tamuly, S.; Dutta, L.J.; Das, S.K. Identification of swinepox virus from natural outbreaks in pig population of Assam. Virusdisease 2018, 29, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Riyesh, T.; Barua, S.; Kumar, N.; Jindal, N.; Bera, B.C.; Narang, G.; Mahajan, N.K.; Arora, D.; Anand, T.; Vaid, R.K.; et al. Isolation and genetic characterization of swinepox virus from pigs in India. Comp. Immunol. Microbiol. Infect. Dis. 2016, 46, 60–65. [Google Scholar] [CrossRef] [PubMed]
- De Boer, G.F. Swinepox. Virus isolation, experimental infections and the differentiation from vaccinia virus infections. Arch. Virol. 1975, 49, 141–150. [Google Scholar] [CrossRef] [PubMed]
- House, J.A.; House, C.A. Chapter 23, Swinepox. In Diseases of Swine, 8th ed.; Straw, B.A., D’Allaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Iowa State University Press: Iowa City, IA, USA, 1993; pp. 291–295. [Google Scholar]
- Cheville, N.F. Immunofluorescent and Morphologic Studies on Swinepox. Pathol. Vet. 1966, 3, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Medaglia, M.L.G.; Pessoa, L.C.G.D.; Sales, E.R.C.; Freitas, T.R.P.; Damaso, C.R. Spread of cantagalo virus to northern Brazil. Emerg. Infect. Dis. 2009, 15, 1142–1143. [Google Scholar] [CrossRef]
- Moussatché, N.; Damaso, C.R.; McFadden, G. When good vaccines go wild: Feral Orthopoxvirus in developing countries and beyond. J. Infect. Dev. Ctries. 2008, 2, 156–173. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Balamurugan, V.; Bhanuprakash, V.; Venkatesan, G.; Hosamani, M. Emergence and reemergence of vaccinia-like viruses: Global scenario and perspectives. Indian J. Virol. 2012, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Medaglia, M.L.G.; Sá, N.M.B.; Correa, I.A.; Costa, L.J.; Damaso, C.R. One-step duplex polymerase chain reaction for the detection of swinepox and vaccinia viruses in skin lesions of swine with poxvirus-related disease. J. Virol. Methods 2015, 219, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Delhon, G.A.; Tulman, E.R.; Afonso, C.L.; Rock, D.L. Genus Suipoxvirus. In Poxviruses; Birkhäuser: Basel, Switzerland, 2007; pp. 203–215. [Google Scholar]
- Steineck, T.; Kolodziejek, J.; Nowotny, N.; Schilcher, F. A case of Swine Pox in a Wild Boar (Sus Scrofa) in Austria. In Proceedings of the 6th Conference of the European Wildlife Disease Association, Uppsala, Sweden, 8–12 September 2004. [Google Scholar]
- Datt, N.S. Comparative studies of pigpox and vaccinia viruses: I. Host range Pathogenicity. J. Comp. Pathol. 1964, 74, 62–69. [Google Scholar] [CrossRef]
- Yuan, X.; Lin, H.; Li, B.; He, K.; Fan, H. Swinepox virus vector-based vaccines: Attenuation and biosafety assessments following subcutaneous prick inoculation. Vet. Res. 2018, 49, 14. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.P.; Hall, M.R.; McFarland, M.D. Immunological responses of cross-bred and inbred miniature pigs to swine poxvirus. Vet. Immunol. Immunopathol. 1989, 23, 149–159. [Google Scholar] [CrossRef]
- Mittal, D.; Mahajan, V.B.; Pathak, D.; Filia, G. Differential diagnosis of swine pox during an outbreak. Indian Vet. J. 2011, 88, 9–11. [Google Scholar]
- Kasza, L.; Griesemer, R.A. Experimental swine pox. Am. J. Vet. Res. 1962, 23, 443–451. [Google Scholar] [PubMed]
- Teppema, J.S.; De Boer, G.F. Ultrastructural aspects of experimental swinepox with special reference to inclusion bodies. Arch. Virol. 1975, 49, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Huang, D.; Liu, S.; Lin, H.; Zhu, H.; Liu, B.; Lu, C. Immune responses and protective efficacy of a recombinant swinepox virus expressing HA1 against swine H1N1 influenza virus in mice and pigs. Vaccine 2012, 30, 3119–3125. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Ma, Z.; Fan, H.; Lu, C. Construction and immunogenicity of recombinant swinepox virus expressing capsid protein of PCV2. Vaccine 2012, 30, 6307–6313. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Lin, H.; Li, B.; He, K.; Fan, H. Efficacy and immunogenicity of recombinant swinepox virus expressing the truncated S protein of a novel isolate of porcine epidemic diarrhea virus. Arch. Virol. 2017, 162, 3779–3789. [Google Scholar] [CrossRef]
- Miller, R.B.; Olson, L.D. Experimental induction of cutaneous streptococcal abscesses in swine as a sequela to swinepox. Am. J. Vet. Res. 1980, 41, 341–347. [Google Scholar] [PubMed]
- Shope, R.E. Swine pox. Arch. Gesamte Virusforsch. 1940, 1, 457–467. [Google Scholar] [CrossRef]
- Borst, G.H.; Kimman, T.G.; Gielkens, A.L.; van der Kamp, J.S. Four sporadic cases of congenital swinepox. Vet. Rec. 1990, 127, 61–63. [Google Scholar]
- Thibault, S.; Drolet, R.; Alain, R.; Dea, S. Congenital swine pox: A sporadic skin disorder in nursing piglets. Swine Health Prod. 1998, 6, 276–278. [Google Scholar]
- Laber, K.E.; Whary, M.T.; Bingel, S.A.; Goodrich, J.A.; Smith, A.C.; Swindle, M.M. Chapter 15, Biology and diseases of swine. In Laboratory Animal Medicine, 2nd ed.; Fox, J.G., Anderson, L.C., Loew, F.M., Quimby, F.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 615–673. [Google Scholar]
- Massung, R.F.; Jayarama, V.; Moyer, R.W. DNA sequence analysis of conserved and unique regions of swinepox virus: Identification of genetic elements supporting phenotypic observations including a novel G protein-coupled receptor homologue. Virology 1993, 197, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; McFadden, G. Modulation of tumor necrosis factor by microbial pathogens. PLoS Pathog. 2006, 2, e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Osorio, F.A.; Balinsky, C.; Kutish, G.F.; Rock, D.L. The genome of swinepox virus. J. Virol. 2002, 76, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Moorkamp, L.; Beineke, A.; Kaim, U.; Diesterbeck, U.; Urstadt, S.; Czerny, C.P.; Rüberg, H.; Grosse Beilage, E. Swinepox-skin disease with sporadic occurrence. Dtsch. Tierarztl. Wochenschr. 2008, 115, 162–166. [Google Scholar]
- Strandström, H.; Veijalainen, P.; Moennig, V.; Hunsmann, G.; Schwarz, H.; Schäfer, W. C-type particles produced by a permanent cell line from a leukemic pig. I. Origin and properties of the host cells and some evidence for the occurrence of C-type-like particles. Virology 1974, 57, 175–178. [Google Scholar] [CrossRef]
- Cagatay, G.N.; Antos, A.; Suckstorff, O.; Isken, O.; Tautz, N.; Becher, P.; Postel, A. Porcine Complement Regulatory Protein CD46 Is a Major Receptor for Atypical Porcine Pestivirus but Not for Classical Swine Fever Virus. J. Virol. 2021, 95, e02186-20. [Google Scholar] [CrossRef]
- Hamel, A.L.; Lin, L.L.; Sachvie, C.; Grudeski, E.; Nayar, G.P. PCR detection and characterization of type-2 porcine circovirus. Can. J. Vet. Res. 2000, 64, 44–52. [Google Scholar]
- Yang, K.; Jiao, Z.; Zhou, D.; Guo, R.; Duan, Z.; Tian, Y. Development of a multiplex PCR to detect and discriminate porcine circoviruses in clinical specimens. BMC Infect. Dis. 2019, 19, 778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, F.A.; Dos Santos Júnior, E.M.; Laguardia-Nascimento, M.; Freitas, T.R.P.; Damaso, C.R.; Rivetti Júnior, A.V.; Camargos, M.F.; Fonseca Júnior, A.A. Validation of a real-time PCR assay for detection of swinepox virus. Arch. Virol. 2019, 164, 3059–3063. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Ervynck, A.; Dobney, K.; Hongo, H.; Meadow, R. Born free? New evidence for the status of “Sus scrofa” at Neolithic Çayönü Tepesi (southeastern Anatolia, Turkey). Paléorient 2001, 27, 47–73. [Google Scholar] [CrossRef]
- Frantz, L.A.F.; Haile, J.; Lin, A.T.; Scheu, A.; Geörg, C.; Benecke, N.; Alexander, M.; Linderholm, A.; Mullin, V.E.; Daly, K.G.; et al. Ancient pigs reveal a near-complete genomic turnover following their introduction to Europe. Proc. Natl. Acad. Sci. USA 2019, 116, 17231–17238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flad, R.K.; Yuan, J.; Li, S. Zooarcheological evidence for animal domestication in northwest China. Dev. Quat. Sci. 2007, 9, 167–203. [Google Scholar]
- Frantz, L.; Meijaard, E.; Gongora, J.; Haile, J.; Groenen, M.A.M.; Larson, G. The Evolution of Suidae. Annu. Rev. Anim. Biosci. 2016, 4, 61–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, J.J.; Brisbin, I.L. Wild Pigs in the United States: Their History, Comparative Morphology, and Current Status; University of Georgia Press: Athens, GA, USA, 2008; ISBN 9780820331379/0820331376. [Google Scholar]
- Aasdev, A.; Mishra, A.; Bora, D.P.; Kurkure, N.V.; Barman, N.N.; Raut, A.A. First complete genome characterization of swinepox virus directly from a clinical sample indicates divergence of a Eurasian-lineage virus. Arch. Virol. 2021, 166, 1217–1225. [Google Scholar] [CrossRef]
- Smithson, C.; Meyer, H.; Gigante, C.M.; Gao, J.; Zhao, H.; Batra, D.; Damon, I.; Upton, C.; Li, Y. Two novel poxviruses with unusual genome rearrangements: NY_014 and Murmansk. Virus Genes 2017, 53, 883–897. [Google Scholar] [CrossRef]
- Olitsky, P.K.; Long, P.H. Relation of Vaccinal immunity to the persistence of the virus in rabbits. J. Exp. Med. 1929, 50, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Maiboroda, A.D. Experimental infection of Norvegian rats (Rattus norvegicus) with ratpox virus. Acta Virol. 1982, 26, 288–291. [Google Scholar]
- Ginsberg, A.H.; Johnson, K.P. The effect of cyclophosphamide on intracerebral vaccinia virus infection in BalbC mice. Exp. Mol. Pathol. 1977, 27, 285–294. [Google Scholar] [CrossRef]
- Sakala, I.G.; Chaudhri, G.; Scalzo, A.A.; Eldi, P.; Newsome, T.P.; Buller, R.M.; Karupiah, G. Evidence for Persistence of Ectromelia Virus in Inbred Mice, Recrudescence Following Immunosuppression and Transmission to Naïve Mice. PLoS Pathog. 2015, 11, e1005342. [Google Scholar] [CrossRef] [PubMed]
- Paez, E.; Dallo, S.; Esteban, M. Generation of a dominant 8-MDa deletion at the left terminus of vaccinia virus DNA. Proc. Natl. Acad. Sci. USA 1985, 82, 3365–3369. [Google Scholar] [CrossRef] [Green Version]
- Pogo, B.G.; Friend, C. Persistent infection of Friend erythroleukemia cells with vaccinia virus. Proc. Natl. Acad. Sci. USA 1982, 79, 4805–4809. [Google Scholar] [CrossRef] [Green Version]
- Buller, R.M.; Weinblatt, A.C.; Hamburger, A.W.; Wallace, G.D. Observations on the replication of ectromelia virus in mouse-derived cell lines: Implications for epidemiology of mousepox. Lab. Anim. Sci. 1987, 37, 28–32. [Google Scholar] [PubMed]
- Harley, J.D.; Gillespie, A.M. A complicated case of congenital vaccinia. Pediatrics 1972, 50, 150–153. [Google Scholar] [PubMed]
- Aitkens, G.H.; Bowman, R.; Hansman, D. A case of foetal vaccinia. Med. J. Aust. 1968, 2, 173–174. [Google Scholar] [CrossRef]
- Green, D.M.; Reid, S.M.; Rhaney, K. Generalised vaccinia in the human foetus. Lancet 1966, 1, 1296–1298. [Google Scholar] [CrossRef]
- Mbala, P.K.; Huggins, J.W.; Riu-Rovira, T.; Ahuka, S.M.; Mulembakani, P.; Rimoin, A.W.; Martin, J.W.; Muyembe, J.-J.T. Maternal and fetal outcomes among pregnant women with human monkeypox infection in the Democratic Republic of Congo. J. Infect. Dis. 2017, 216, 824–828. [Google Scholar] [CrossRef]
- Wisser, J.; Pilaski, J.; Strauss, G.; Meyer, H.; Burck, G.; Truyen, U.; Rudolph, M.; Frölich, K. Cowpox virus infection causing stillbirth in an Asian elephant (Elphas maximus). Vet. Rec. 2001, 149, 244–246. [Google Scholar] [CrossRef]
- Kalter, S.S.; Heberling, R.L.; Panigel, M.; Brack, M.; Felsburg, P.J. Fetal infection of the baboon (Papio cynocephalus) with vaccinia virus. Proc. Soc. Exp. Biol. Med. 1973, 143, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.R. Smallpox, 1st ed.; The Kothari Book Depot: Bombay, India, 1972. [Google Scholar]
- Kasza, L.; Bohl, E.H.; Jones, D.O. Isolation and cultivation of swine pox virus in primary cell cultures of swine origin. Am. J. Vet. Res. 1960, 21, 269–273. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host-ID | Species | Age of Animals | Tissue | RT-PCR (Ct Value) |
|---|---|---|---|---|
| 101-200136 | Domestic pig | 1 day 1 | FFPE skin | 23.81 |
| 201-200045 | Domestic pig | 1 day 1 | skin | 15.78 |
| 201-200046 | Domestic pig | 1 day | skin | 18.96 |
| 201-200047 | Domestic pig | 1 day | tongue | 28.33 |
| 201-200047 | Domestic pig | 1 day | skin | 19.02 |
| 201-200047 | Domestic pig | 1 day | lung | 37.03 |
| 201-200047 | Domestic pig | 1 day | intestine | 31.37 |
| 201-200047 | Domestic pig | 1 day | umbilical cord | 21.43 |
| 201-200047 | Domestic pig | 1 day | esophagus | 28.24 |
| 201-200070 | Wild boar | 8 weeks 2 | skin | 15.95 |
| 201-200071 | Wild boar | 8 weeks 2 | skin | 14.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaiser, F.K.; Wiedemann, A.; Kühl, B.; Menke, L.; Beineke, A.; Baumgärtner, W.; Wohlsein, P.; Rigbers, K.; Becher, P.; Peters, M.; et al. Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes. Viruses 2021, 13, 2038. https://doi.org/10.3390/v13102038
Kaiser FK, Wiedemann A, Kühl B, Menke L, Beineke A, Baumgärtner W, Wohlsein P, Rigbers K, Becher P, Peters M, et al. Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes. Viruses. 2021; 13(10):2038. https://doi.org/10.3390/v13102038
Chicago/Turabian StyleKaiser, Franziska K., Anastasia Wiedemann, Bianca Kühl, Laura Menke, Andreas Beineke, Wolfgang Baumgärtner, Peter Wohlsein, Kerstin Rigbers, Paul Becher, Martin Peters, and et al. 2021. "Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes" Viruses 13, no. 10: 2038. https://doi.org/10.3390/v13102038
APA StyleKaiser, F. K., Wiedemann, A., Kühl, B., Menke, L., Beineke, A., Baumgärtner, W., Wohlsein, P., Rigbers, K., Becher, P., Peters, M., Osterhaus, A. D. M. E., & Ludlow, M. (2021). Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes. Viruses, 13(10), 2038. https://doi.org/10.3390/v13102038