
Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Virus Isolation
2.3. Subtyping and Species Identification
2.4. Sequencing and Phylogenetic Analysis
2.5. Pathogenicity in Mice
2.6. Statistical Analysis
2.7. Ethical Approval
3. Results
3.1. Identification of H7NX AIV Isolates
3.2. Molecular Characterization of H7 AIV Isolates
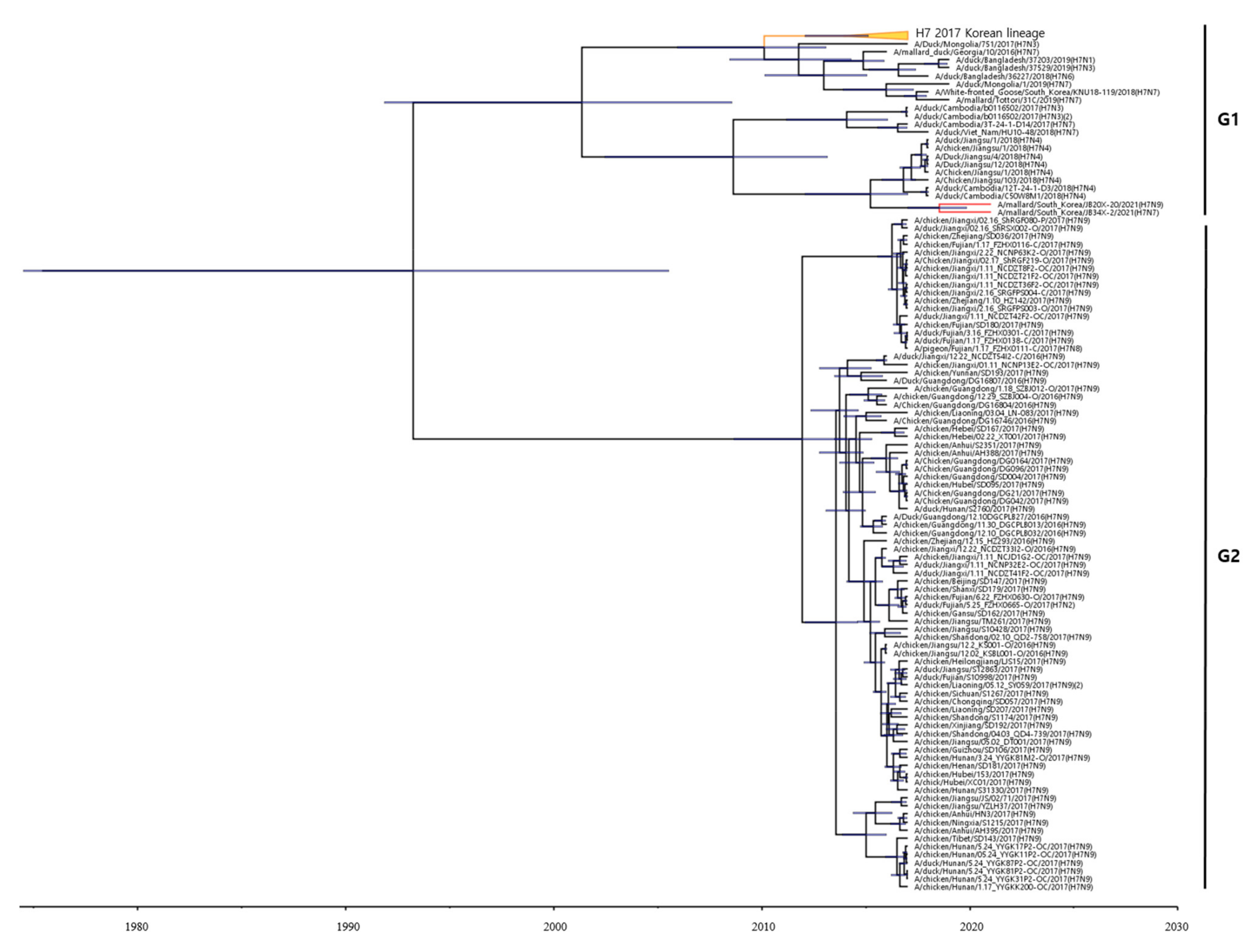
3.3. Phylogenetic Analysis of H7 AIVs Isolates
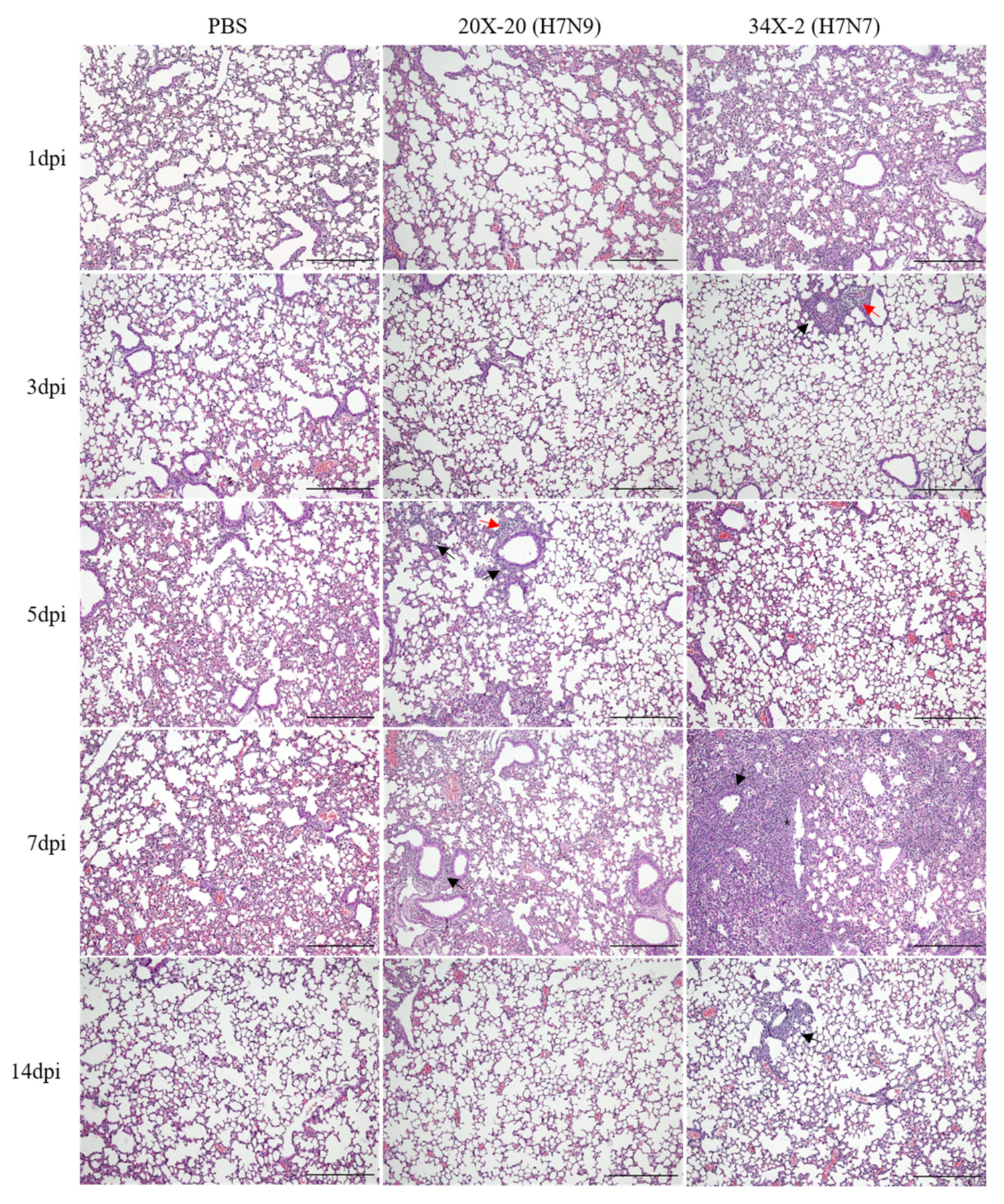
3.4. Pathogenicity in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamb, R.A.; Krug, R.M. Orthomyxoviridae: The viruses and their replication. Fields Virol. 2001, 1, 1487–1531. [Google Scholar]
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco-Cabrera, S.; Gómez, J.; et al. New world bats harbor diverse influenza a viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.A.; Chen, L.-M.; Recuenco-Cabrera, S.; Ellison, J.; Davis, C.T.; York, I.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, J.; Elbers, A.; Bouma, A.; Koch, G.; de Wit, J.; Stegeman, J. Transmission characteristics of low pathogenic avian influenza virus of H7N7 and H5N7 subtypes in layer chickens. Vet. Microbiol. 2012, 155, 207–213. [Google Scholar] [CrossRef]
- Shi, J.; Deng, G.; Kong, H.; Gu, C.; Ma, S.; Yin, X.; Zeng, X.; Cui, P.; Chen, Y.; Yang, H.; et al. H7N9 virulent mutants detected in chickens in China pose an increased threat to humans. Cell Res. 2017, 27, 1409–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horimoto, T.; Rivera, E.; Pearson, J.; Senne, D.; Krauss, S.; Kawaoka, Y.; Webster, R. Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexico. Virology 1995, 213, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Banks, J.; Speidel, E.S.; Moore, E.; Plowright, L.; Piccirillo, A.; Capua, I.; Cordioli, P.; Fioretti, A.; Alexander, D.J. Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Arch. Virol. 2001, 146, 963–973. [Google Scholar] [CrossRef]
- Berhane, Y.; Hisanaga, T.; Kehler, H.; Neufeld, J.; Manning, L.; Argue, C.; Handel, K.; Hooper-McGrevy, K.; Jonas, M.; Robinson, J.; et al. Highly pathogenic avian influenza virus A (H7N3) in Domestic Poultry, Saskatchewan, Canada, 2007. Emerg. Infect. Dis. 2009, 15, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Suarez, D.L.; Senne, D.A.; Banks, J.; Brown, I.H.; Essen, S.C.; Lee, C.-W.; Manvell, R.J.; Mathieu-Benson, C.; Moreno, V.; Pedersen, J.C.; et al. Recombination resulting in virulence shift in Avian Influenza Outbreak, Chile. Emerg. Infect. Dis. 2004, 10, 693–699. [Google Scholar] [CrossRef]
- Beerens, N.; Heutink, R.; Harders, F.; Bossers, A.; Koch, G.; Peeters, B. Emergence and selection of a highly pathogenic avian influenza H7N3 virus. J. Virol. 2020, 94, e01818-19. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Torchetti, M.K.; Killian, M.L.; Berhane, Y.; Swayne, D.E. Highly pathogenic avian influenza A (H7N9) virus, Tennessee, USA, March 2017. Emerg. Infect. Dis. 2017, 23, 1860. [Google Scholar] [CrossRef] [PubMed]
- Killian, M.L.; Kim-Torchetti, M.; Hines, N.; Yingst, S.; DeLiberto, T.; Lee, D.-H. Outbreak of h7n8 low pathogenic avian influenza in commercial turkeys with spontaneous mutation to highly pathogenic avian influenza. Genome Announc. 2016, 4, e00457-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youk, S.; Lee, D.-H.; Killian, M.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Torchetti, M.K. Highly pathogenic avian influenza A (H7N3) virus in Poultry, United States, 2020. Emerg. Infect. Dis. 2020, 26, 2966. [Google Scholar] [CrossRef]
- Byrne, A.; Reid, S.; Seekings, A.; Núñez, A.; Prieto, A.O.; Ridout, S.; Warren, C.; Puranik, A.; Ceeraz, V.; Essen, S.; et al. H7N7 avian influenza virus mutation from low to high pathogenicity on a layer chicken farm in the UK. Viruses 2021, 13, 259. [Google Scholar] [CrossRef]
- Naguib, M.M.; Verhagen, J.H.; Mostafa, A.; Wille, M.; Li, R.; Graaf, A.; Järhult, J.D.; Ellström, P.; Zohari, S.; Lundkvist, Å.; et al. Global patterns of avian influenza A (H7): Virus evolution and zoonotic threats. FEMS Microbiol. Rev. 2019, 43, 608–621. [Google Scholar] [CrossRef] [Green Version]
- Abdelwhab, E.M.; Veits, J.; Mettenleiter, T.C. Prevalence and control of H7 avian influenza viruses in birds and humans. Epidemiol. Infect. 2014, 142, 896–920. [Google Scholar] [CrossRef]
- Kim, M.-C.; Jeong, O.-M.; Kang, H.-M.; Paek, M.-R.; Kwon, J.-S.; Song, C.-S.; Kwon, Y.-K.; Lee, J.-G.; Kwon, J.-H.; Lee, Y.-J. Pathogenicity and transmission studies of H7N7 avian influenza virus isolated from feces of magpie origin in chickens and magpie. Vet. Microbiol. 2010, 141, 268–274. [Google Scholar] [CrossRef]
- Kim, H.-R.; Park, C.-K.; Lee, Y.-J.; Oem, J.-K.; Kang, H.-M.; Choi, J.-G.; Lee, O.-S.; Bae, Y.-C. Low pathogenic H7 subtype avian influenza viruses isolated from domestic ducks in South Korea and the close association with isolates of wild birds. J. Gen. Virol. 2012, 93, 1278–1287. [Google Scholar] [CrossRef]
- Lee, E.-K.; Lee, Y.-N.; Kye, S.-J.; Lewis, N.S.; Brown, I.H.; Sagong, M.; Heo, G.-B.; Kang, Y.-M.; Cho, H.-K.; Kang, H.-M.; et al. Characterization of a novel reassortant H5N6 highly pathogenic avian influenza virus clade 2.3.4.4 in Korea, 2017. Emerg. Microbes Infect. 2018, 7, 103. [Google Scholar] [CrossRef] [Green Version]
- Herfst, S.; Imai, M.; Kawaoka, Y.; Fouchier, R.A.M. Avian influenza virus transmission to mammals. In Influenza Pathogenesis and Control; Current Topics in Microbiology and Immunology 385; Springer: Berlin, Germany, 2014; Volume I, pp. 137–155. [Google Scholar] [CrossRef]
- Reperant, L.; Rimmelzwaan, G.; Kuiken, T. Avian influenza viruses in mammals. Rev. Sci. Tech. L’oie 2009, 28, 137–159. [Google Scholar] [CrossRef]
- Kurtz, J.; Manvell, R.J.; Banks, J. Avian influenza virus isolated from a woman with conjunctivitis. Lancet 1996, 348, 901–902. [Google Scholar] [CrossRef]
- Fouchier, R.A.M.; Schneeberger, P.M.; Rozendaal, F.W.; Broekman, J.M.; Kemink, S.A.G.; Munster, V.; Kuiken, T.; Rimmelzwaan, G.F.; Schutten, M.; van Doornum, G.J.J.; et al. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc. Natl. Acad. Sci. USA 2004, 101, 1356–1361. [Google Scholar] [CrossRef] [Green Version]
- Puzelli, S.; Rossini, G.; Facchini, M.; Vaccari, G.; Di Trani, L.; Di Martino, A.; Gaibani, P.; Vocale, C.; Cattoli, G.; Bennett, M.; et al. Human infection with highly pathogenic A (H7N7) avian influenza virus, Italy, 2013. Emerg. Infect. Dis. 2014, 20, 1741–1745. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Xiao, L.; Li, L. Research progress on human infection with avian influenza H7N9. Front. Med. 2020, 14, 8–20. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Deng, G.; Ma, S.; Zeng, X.; Yin, X.; Li, M.; Zhang, B.; Cui, P.; Chen, Y.; Yang, H.; et al. Rapid evolution of H7N9 highly pathogenic viruses that emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippon, D.A.M.; Wu, P.; Cowling, B.J.; Lau, E.H.Y. Avian influenza human infections at the human-animal interface. J. Infect. Dis. 2020, 222, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Deng, G.; Zeng, X.; Cui, P.; Hou, Y.; Liu, Y.; Fang, J.; Pan, S.; Wang, D.; Chen, X.; et al. Genetic and biological properties of H7N9 avian influenza viruses detected after application of the H7N9 poultry vaccine in China. PLoS Pathog. 2021, 17, e1009561. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tian, G.; Shi, J.; Deng, G.; Li, C.; Chen, H. Vaccination of poultry successfully eliminated human infection with H7N9 virus in China. Sci. China Life Sci. 2018, 61, 1465–1473. [Google Scholar] [CrossRef]
- Belser, J.A.; Bridges, C.B.; Katz, J.M.; Tumpey, T.M. Past, present, and possible future human infection with influenza virus a subtype H7. Emerg. Infect. Dis. 2009, 15, 859–865. [Google Scholar] [CrossRef]
- Huo, X.; Cui, L.-B.; Chen, C.; Wang, D.; Qi, X.; Zhou, M.-H.; Guo, X.; Wang, F.; Liu, W.J.; Kong, W.; et al. Severe human infection with a novel avian-origin influenza A(H7N4) virus. Sci. Bull. 2018, 63, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Stear, M.J. OIE manual of diagnostic tests and vaccines for terrestrial animals (mammals, birds and bees) 5th Edn. Volumes 1 & 2. World organization for animal health 2004. ISBN 92 9044 622 6. €140. Parasitology 2005, 130, 727. [Google Scholar] [CrossRef]
- Hoffmann, B.; Hoffmann, D.; Henritzi, D.; Beer, M.; Harder, T.C. Riems influenza a typing array (RITA): An RT-qPCR-based low density array for subtyping avian and mammalian influenza a viruses. Sci. Rep. 2016, 6, 27211. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Lee, H.-J.; Kang, H.-M.; Jeong, O.-M.; Kim, M.-C.; Kwon, J.-S.; Kwon, J.-H.; Kim, C.-B.; Lee, J.-B.; Park, S.-Y.; et al. DNA barcoding techniques for avian influenza virus surveillance in migratory bird habitats. J. Wildl. Dis. 2010, 46, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.; Muench, H. A simple method of estimating fifty percent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and viral determinants of influenza a virus species specificity. Nat. Rev. Microbiol. 2019, 17, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Blixt, O.; Stevens, J.; Lipatov, A.S.; Davis, C.T.; Collins, B.E.; Cox, N.J.; Paulson, J.C.; Donis, R.O. In vitro evolution of H5N1 avian influenza virus toward human-type receptor specificity. Virology 2012, 422, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Zhang, W.; Wang, F.; Qi, J.; Wu, Y.; Song, H.; Gao, F.; Bi, Y.; Zhang, Y.; Fan, Z.; et al. Structures and receptor binding of hemagglutinins from human-infecting H7N9 influenza viruses. Science 2013, 342, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ishaq, M.; Prudence, M.; Xi, X.; Hu, T.; Liu, Q.; Guo, D. Single mutation at the amino acid position 627 of PB2 that leads to increased virulence of an H5N1 avian influenza virus during adaptation in mice can be compensated by multiple mutations at other sites of PB2. Virus Res. 2009, 144, 123–129. [Google Scholar] [CrossRef]
- Jonges, M.; Welkers, M.; Jeeninga, R.; Meijer, A.; Schneeberger, P.; Fouchier, R.; De Jong, M.; Koopmans, M. Emergence of the virulence-associated PB2 E627K substitution in a fatal human case of highly pathogenic avian influenza virus A(H7N7) infection as determined by illumina ultra-deep sequencing. J. Virol. 2013, 88, 1694–1702. [Google Scholar] [CrossRef] [Green Version]
- Conenello, G.M.; Zamarin, D.; Perrone, L.A.; Tumpey, T.; Palese, P. A single mutation in the PB1-F2 of H5N1 (HK/97) and 1918 influenza a viruses contributes to increased virulence. PLoS Pathog. 2007, 3, e141. [Google Scholar] [CrossRef]
- Senne, D.A.; Panigrahy, B.; Kawaoka, Y.; Pearson, J.E.; Süss, J.; Lipkind, M.; Kida, H.; Webster, R.G. Survey of the hemagglutinin (HA) cleavage site sequence of H5 and H7 avian influenza viruses: Amino acid sequence at the HA cleavage site as a marker of pathogenicity potential. Avian Dis. 1996, 40, 425–437. [Google Scholar] [CrossRef]
- Fan, S.; Hatta, M.; Kim, J.H.; Halfmann, P.; Imai, M.; Macken, C.A.; Le, M.Q.; Nguyen, T.; Neumann, G.; Kawaoka, Y. Novel residues in avian influenza virus PB2 protein affect virulence in mammalian hosts. Nat. Commun. 2014, 5, 5021. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.; Ma, S.; Wang, J.; Gu, C.; Wang, Z.; Shi, J.; Deng, G.; Guan, Y.; Chen, H. Identification of key amino acids in the PB2 and M1 proteins of H7N9 influenza virus that affect its transmission in guinea pigs. J. Virol. 2019, 94, e01180-19. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Yuan, S.; Zhang, K.; Singh, K.; Ma, Q.; Zhou, J.; Chu, H.; Zheng, B.-J. PB2 substitutions V598T/I increase the virulence of H7N9 influenza A virus in mammals. Virology 2017, 501, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, E.K.; London, W.; Murphy, B.R. A single amino acid in the PB2 gene of influenza A virus is a determinant of host range. J. Virol. 1993, 67, 1761–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zhang, Y.; Shinya, K.; Deng, G.; Jiang, Y.; Li, Z.; Guan, Y.; Tian, G.; Li, Y.; Shi, J.; et al. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog. 2009, 5, e1000709. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Jiang, L.; Li, J.; Zhao, Q.; Wang, J.; He, X.; Huang, S.; Wang, Q.; Zhao, Y.; Wang, G.; et al. Low polymerase activity attributed to pa drives the acquisition of the PB2 E627K mutation of H7N9 avian influenza virus in mammals. Mbio 2019, 10, e01162-19. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Uchida, Y.; Tanikawa, T.; Maeda, N.; Takemae, N.; Saito, T. Amino acid substitutions in PB1 of avian influenza viruses influence pathogenicity and transmissibility in chickens. J. Virol. 2014, 88, 11130–11139. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Wang, Z.; Shi, J.; Deng, G.; Kong, H.; Tao, S.; Li, C.; Liu, L.; Guan, Y.; Chen, H. Glycine at position 622 in PB1 contributes to the virulence of H5N1 avian influenza virus in mice. J. Virol. 2016, 90, 1872–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamayoshi, S.; Yamada, S.; Fukuyama, S.; Murakami, S.; Zhao, D.; Uraki, R.; Watanabe, T.; Tomita, Y.; Macken, C.; Neumann, G.; et al. Virulence-affecting amino acid changes in the PA protein of H7N9 influenza a viruses. J. Virol. 2014, 88, 3127–3134. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, H.; Xu, J.; Zhao, D.; Shi, J.; Li, Y.; Deng, G.; Jiang, Y.; Li, X.; Zhu, P.; et al. The PA protein directly contributes to the virulence of H5N1 avian influenza viruses in domestic ducks. J. Virol. 2011, 85, 2180–2188. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Zou, X.; Zhou, J.; Tang, J.; Shu, Y. Residues 41V and/or 210D in the NP protein enhance polymerase activities and potential replication of novel influenza (H7N9) viruses at low temperature. Virol. J. 2015, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Suzuki, K.; Sakurai, Y.; Kubo, M.; Okada, H.; Itoh, T.; Tsukamoto, K. NP body domain and PB2 contribute to increased virulence of H5N1 highly pathogenic avian influenza viruses in chickens. J. Virol. 2010, 85, 1834–1846. [Google Scholar] [CrossRef] [Green Version]
- Wasilenko, J.L.; Sarmento, L.; Pantin-Jackwood, M.J. A single substitution in amino acid 184 of the NP protein alters the replication and pathogenicity of H5N1 avian influenza viruses in chickens. Arch. Virol. 2009, 154, 969–979. [Google Scholar] [CrossRef]
- Li, J.; Zheng, W.; Hou, L.; Chen, C.; Fan, W.; Qu, H.; Jiang, J.; Liu, J.; Gao, G.F.; Zhou, J.; et al. Differential nucleocytoplasmic shuttling of the nucleoprotein of influenza a viruses and association with host tropism. Cell. Microbiol. 2016, 19, e12692. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Zhang, B.; Shi, J.; Yin, X.; Wang, G.; Cui, P.; Liu, L.; Deng, G.; Jiang, Y.; Li, C.; et al. Amino acid mutations A286V and T437M in the nucleoprotein attenuate H7N9 viruses in mice. J. Virol. 2020, 94, e01530-19. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Xiao, H.; Chen, Q.; Wu, Y.; Fu, L.; Quan, C.; Wong, G.; Liu, J.; Haywood, J.; Liu, Y.; et al. Changes in the length of the neuraminidase stalk region impact H7N9 virulence in mice. J. Virol. 2016, 90, 2142–2149. [Google Scholar] [CrossRef] [Green Version]
- Kode, S.S.; Pawar, S.D.; Tare, D.S.; Keng, S.S.; Hurt, A.C.; Mullick, J. A novel I117T substitution in neuraminidase of highly pathogenic avian influenza H5N1 virus conferring reduced susceptibility to oseltamivir and zanamivir. Vet. Microbiol. 2019, 235, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Nao, N.; Kajihara, M.; Manzoor, R.; Maruyama, J.; Yoshida, R.; Muramatsu, M.; Miyamoto, H.; Igarashi, M.; Eguchi, N.; Sato, M.; et al. A single amino acid in the M1 protein responsible for the different pathogenic potentials of H5N1 highly pathogenic avian influenza virus strains. PLoS ONE 2015, 10, e0137989. [Google Scholar] [CrossRef] [Green Version]
- Cheung, C.; Rayner, J.M.; Smith, G.J.; Wang, P.; Naipospos, T.S.P.; Zhang, J.; Yuen, K.-Y.; Webster, R.G.; Peiris, J.S.M.; Guan, Y.; et al. Distribution of amantadine-resistant H5N1 avian influenza variants in Asia. J. Infect. Dis. 2006, 193, 1626–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wang, G.; Zhang, H.; Xin, G.; Zhang, D.; Zeng, J.; Chen, X.; Xu, Y.; Cui, Y.; Li, K. Effects of NS1 variants of H5N1 influenza virus on interferon induction, TNFα response and p53 activity. Cell. Mol. Immunol. 2010, 7, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Dankar, S.K.; Wang, S.; Ping, J.; Forbes, N.E.; Keleta, L.; Li, Y.; Brown, E.G. Influenza a virus NS1 gene mutations F103L and M106I increase replication and virulence. Virol. J. 2011, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, K.; Chen, Q.; Zhang, X.; Sun, Y.; Bi, Y.; Zhang, S.; Gu, J.; Li, J.; Liu, D.; et al. Three amino acid substitutions in the NS1 protein change the virus replication of H5N1 influenza virus in human cells. Virology 2018, 519, 64–73. [Google Scholar] [CrossRef]
- Rutten, N.; Gonzales, J.L.; Elbers, A.R.W.; Velthuis, A. Cost analysis of various low pathogenic avian influenza surveillance systems in the Dutch egg layer sector. PLoS ONE 2012, 7, e33930. [Google Scholar] [CrossRef]
- Thompson, J.; Seitzinger, A.H. Economic evaluation of low pathogenic avian influenza in northeastern US live bird markets. J. Appl. Poult. Res. 2019, 28, 78–84. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, H.; Wu, Y.; Liu, D.; Qi, X.; Shi, Y.; Gao, G.F. H7N9: A low pathogenic avian influenza A virus infecting humans. Curr. Opin. Virol. 2014, 5, 91–97. [Google Scholar] [CrossRef]
- Shi, Y.; Wu, Y.; Zhang, W.; Qi, J.; Gao, G.F. Enabling the ‘host jump’: Structural determinants of receptor-binding specificity in influenza A viruses. Nat. Rev. Genet. 2014, 12, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Koçer, Z.A.; Poulson, R.L.; Gabbard, J.D.; Howerth, E.W.; Jones, C.A.; Friedman, K.; Seiler, J.; Danner, A.; Kercher, L.; et al. Potential for low-pathogenic avian H7 influenza a viruses to replicate and cause disease in a mammalian model. J. Virol. 2017, 91, e01934-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Li, X.; Tang, L.; Wang, T.; He, G. Characterization of the low-pathogenic H7N7 avian influenza virus in Shanghai, China. Poult. Sci. 2020, 100, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Sun, X.; Brock, N.; Pulit-Penaloza, J.A.; Jones, J.; Zanders, N.; Davis, C.T.; Tumpey, T.M.; Maines, T.R. Mammalian pathogenicity and transmissibility of low pathogenic avian influenza H7N1 and H7N3 viruses isolated from North America in 2018. Emerg. Microbes Infect. 2020, 9, 1037–1045. [Google Scholar] [CrossRef]
- Zhu, W.; Dong, J.; Zhang, Y.; Yang, L.; Li, X.; Chen, T.; Zhao, X.; Wei, H.; Bo, H.; Zeng, X.; et al. A gene constellation in avian influenza a (H7N9) viruses may have facilitated the fifth wave outbreak in China. Cell Rep. 2018, 23, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Tscherne, D.M.; García-Sastre, A. Virulence determinants of pandemic influenza viruses. J. Clin. Investig. 2011, 121, 6–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viral Protein | Amino Acid Residue | Virus Strains | Comments | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|
| 20X-20 (2021) | 34X-2 (2021) | Chicken/1 (2018) | Jiangsu/1 (2018) | Anhui/1 (2013) | Italy/3 (2013) | ||||
| HA | Cleavage site | ELPKGR/GLF | ELPKGR/GLF | ELPKGR/GLF | ELPKGR/GLF | ELPKGR/GLF | ETPKRRERR/GLF | LPAIV -monobasic | [49] |
| G186V | G | G | G | G | V | G | Increased α2-6 binding | [45] | |
| E190D | E | E | E | E | E | E | [43] | ||
| Q/G225D | G | G | G | G | G | G | [43] | ||
| Q226L | Q | Q | Q | Q | L | Q | [43] | ||
| S227N | S | S | S | S | S | S | [44] | ||
| G228S | G | G | G | G | G | G | [43] | ||
| PB2 | L89V | V | V | V | V | V | V | Increased polymerase activity in mammalian cell lines and mice | [46] |
| I147T | I | I | I | I | I | T | [50] | ||
| I292V | I | I | I | I | V | I | [51] | ||
| G309D | D | D | D | D | D | D | [46] | ||
| T339K | K | K | K | K | K | K | [46] | ||
| K389R | R | R | R | R | K | R | [52] | ||
| E627K | E | E | E | K | K | E | Increased virulence in mice | [53] | |
| D701N | D | D | D | D | D | D | [54,55] | ||
| V598T/I | T | T | T | T | V | T | Increased polymerase activity in mammalian cell line | [52] | |
| PB1 | C38Y | Y | Y | Y | Y | Y | Y | Increased polymerase activity in mammalian cell line and pathogenicity in chicken | [56] |
| D622G | G | G | G | G | G | G | Increased polymerase activity and virulence in mice | [57] | |
| PB1-F2 | N66S | S | S | N | N | N | N | Increased virulence in mice | [48] |
| PA | S37A | A | A | A | A | S | A | Increased polymerase activity in mammalian cell line | [58] |
| K142R | R | K | K | K | K | K | [55] | ||
| N383D | D | D | D | D | D | D | Increased pathogenicity in ducks | [59] | |
| N409S | S | S | S | S | N | S | Increased polymerase activity in mammalian cell line | [58] | |
| NP | I41V | I | I | I | I | I | I | Increased polymerase activity in mammalian cell line | [60] |
| M105V | M | V | M | M | V | V | Increased virulence in chicken | [61] | |
| A184K | K | K | K | K | K | K | [62] | ||
| F253I | I | I | I | I | I | I | Increased virulence in mice | [63] | |
| V286A | A | A | A | A | A | A | [64] | ||
| M437T | T | T | T | T | T | T | |||
| NA | 69–73 deletion (QISNT) | No | No | No | No | Yes | No | Increased virulence in mice | [65] |
| I117T | T | T | I | I | T | T | Increased resistance to antiviral drugs (oseltamivir and zanamivir) | [66] | |
| M1 | N30D | D | D | D | D | D | D | Increased virulence in mice | [50] |
| I43M | M | M | M | M | M | M | [67] | ||
| T215A | A | A | A | A | A | A | [50] | ||
| M2 | L26F | L | L | L | L | L | L | Increased resistance to antiviral drugs (amantadine and rimantadine) | [68] |
| S31N | S | S | S | S | N | S | [68] | ||
| NS1 | P42S | S | S | S | S | S | S | Increased virulence in mice | [69] |
| D92E | D | D | D | D | D | D | Increased virulence in mice | [70] | |
| L103F | F | F | F | F | L | F | Increased replication and virulence in mice | [71] | |
| C138F | F | F | F | F | F | F | Increased replication in mammalian cell line | [72] | |
| Gene | 20X-20 | Genetic Identity | 34X-2 | Genetic Identity |
|---|---|---|---|---|
| PB2 | A/wild_duck/South_Korea/KNU18-106/2018(H7N7) | 98.3% | A/red-crowned crane/South Korea/H1026/2017(H7N7) | 96.6% |
| PB1 | A/red-crowned crane/South Korea/H1026/2017(H7N7) | 95% | A/wild_duck/South_Korea/KNU18-104/2018(H7N7) | 93.6% |
| PA | A/wild_duck/South_Korea/KNU18-106/2018(H7N7) | 99.1% | A/wild_bird/Eastern_China/1758/2017(H5N3) | 98.6% |
| HA | A/wild_duck/South_Korea/KNU18-106/2018(H7N7) | 97.8% | A/wild_duck/South_Korea/KNU18-106/2018(H7N7) | 97.3% |
| NP | A/common teal/Shanghai/NH110923/2019(H1N1) | 98.8% | A/duck/Mongolia/926/2019(H5N3) | 99.3% |
| NA | A/Anas platyrhynchos/South Korea/JB31-96/2019(H11N9) | 98.2% | A/mallard/Korea/A15/2016(H7N7) | 97.4% |
| MP | A/duck/Mongolia/916/2018(H3N8) | 99.7% | A/northern pintail/Alaska/362/2013(H3N8) | 99.4% |
| NS | A/duck/Bangladesh/37509/2019(H8N4) | 99.6% | A/duck/Bangladesh/37509/2019(H8N4) | 99.6% |
| Isolate | Days Post Infection (Mean ± SD, log10EID50/mL) | ||||
|---|---|---|---|---|---|
| 1 | 3 | 5 | 7 | 14 | |
| 20X-20 (H7N9) | 3/3 (2.36 ± 1.01) | 3/3 (1.31 ± 2.28) | 3/3 (2.80 ± 2.43) | 3/3 (0.48 ± 0.83) | 0/3 |
| 34X-2 (H7N7) | 3/3 (2.71 ± 2.36) | 3/3 (4.28 ± 0.14) | 3/3 (4.14 ± 0.10) | 3/3 (3.86 ± 0.80) | 0/3 |
| Seroconversion: Positive/Total (HI Titers) | |||
|---|---|---|---|
| SerumVirus | 20X-20 | 34X-2 | |
| Antigen | |||
| 20X-20 | 3/3 (8, 32, 32) | 2/3 (8, 32) | |
| 34X-2 | 3/3 (8, 32, 32) | 2/3 (8, 32) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na, E.-J.; Kim, Y.-S.; Kim, Y.-J.; Park, J.-S.; Oem, J.-K. Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea. Viruses 2021, 13, 2057. https://doi.org/10.3390/v13102057
Na E-J, Kim Y-S, Kim Y-J, Park J-S, Oem J-K. Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea. Viruses. 2021; 13(10):2057. https://doi.org/10.3390/v13102057
Chicago/Turabian StyleNa, Eun-Jee, Young-Sik Kim, Yoon-Ji Kim, Jun-Soo Park, and Jae-Ku Oem. 2021. "Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea" Viruses 13, no. 10: 2057. https://doi.org/10.3390/v13102057
APA StyleNa, E. -J., Kim, Y. -S., Kim, Y. -J., Park, J. -S., & Oem, J. -K. (2021). Genetic Characterization and Pathogenicity of H7N7 and H7N9 Avian Influenza Viruses Isolated from South Korea. Viruses, 13(10), 2057. https://doi.org/10.3390/v13102057