
Flavivirus Persistence in Wildlife Populations


Abstract
:1. Introduction
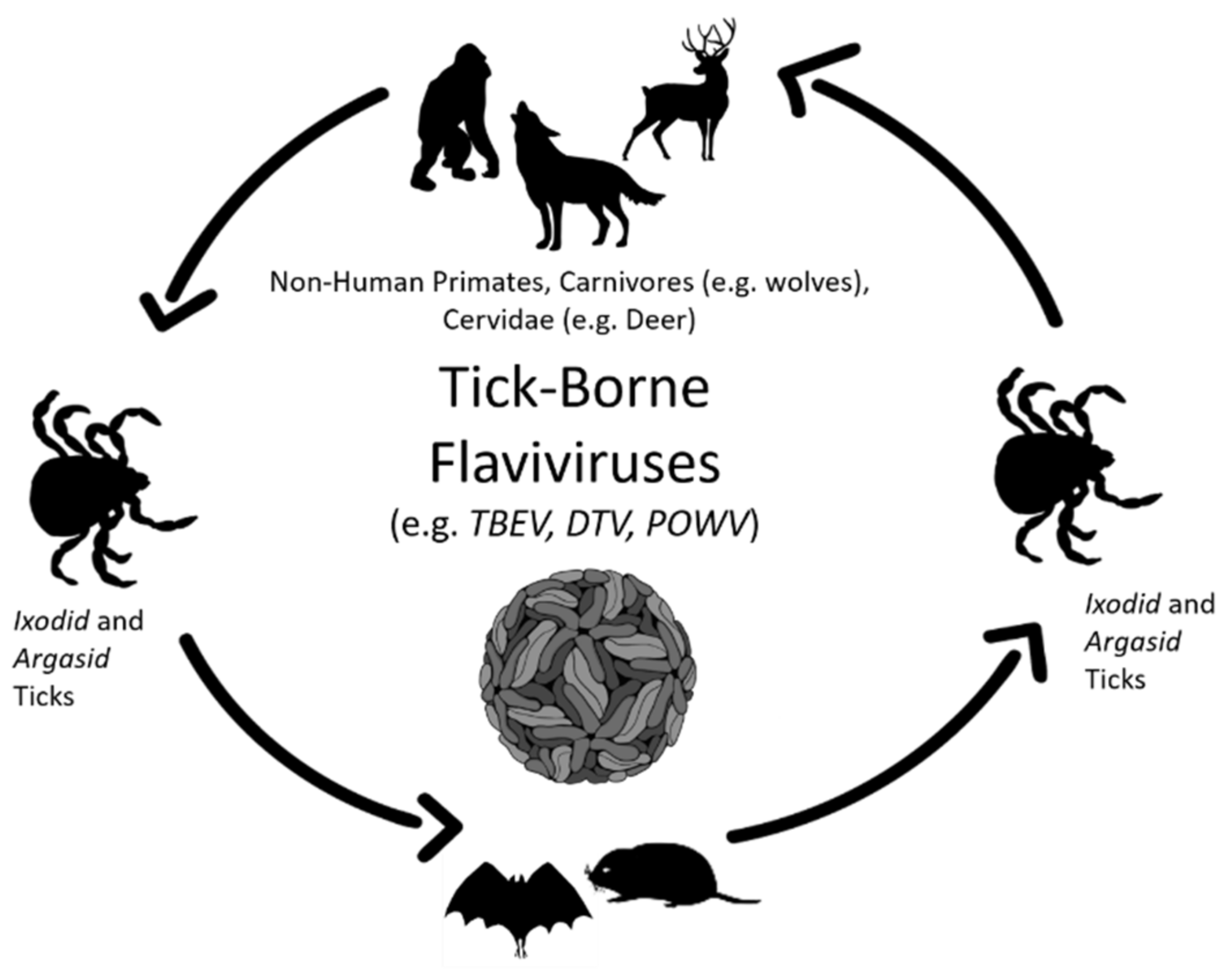
2. Infection and Persistence of Flaviviruses in Wildlife
3. Mechanisms of Flavivirus Persistence
4. Detection
5. Potential Areas of Future Research Endeavors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, W.S.; Gurgel-Goncalves, R.; Garcez, L.M.; Abad-Franch, F. Deforestation effects on Attalea palms and their resident Rhodnius, vectors of Chagas disease, in eastern Amazonia. PLoS ONE 2021, 16, e0252071. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.L.; Guegan, J.F.; Andreou, D.; Marsollier, L.; Carolan, K.; Le Croller, M.; Sanhueza, D.; Gozlan, R.E. Deforestation-driven food-web collapse linked to emerging tropical infectious disease, Mycobacterium ulcerans. Sci. Adv. 2016, 2, e1600387. [Google Scholar] [CrossRef] [Green Version]
- Chaves, L.F.; Cohen, J.M.; Pascual, M.; Wilson, M.L. Social exclusion modifies climate and deforestation impacts on a vector-borne disease. PLoS Negl. Trop. Dis. 2008, 2, e176. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Dorey-Robinson, D.; Hernandez-Triana, L.M.; Phipps, L.P.; Johnson, N. Emerging threats to animals in the United Kingdom by Arthropod-Borne diseases. Front. Vet. Sci. 2020, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Migne, C.V.; Moutailler, S.; Attoui, H. Strategies for assessing arbovirus genetic variability in vectors and/or mammals. Pathogens 2020, 9, 915. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV virus taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Banyai, K. Tick-borne flaviviruses, with a focus on powassan virus. Clin. Microbiol. Rev. 2019, 32, e00106-17. [Google Scholar] [CrossRef] [Green Version]
- Beck, C.; Jimenez-Clavero, M.A.; Leblond, A.; Durand, B.; Nowotny, N.; Leparc-Goffart, I.; Zientara, S.; Jourdain, E.; Lecollinet, S. Flaviviruses in Europe: Complex circulation patterns and their consequences for the diagnosis and control of West Nile disease. Int. J. Environ. Res. Public Health 2013, 10, 6049–6083. [Google Scholar] [CrossRef] [Green Version]
- Fatmi, S.S.; Zehra, R.; Carpenter, D.O. Powassan virus—A new reemerging tick-borne disease. Front. Public Health 2017, 5, 342. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef]
- Trovato, M.; Sartorius, R.; D’Apice, L.; Manco, R.; De Berardinis, P. Viral emerging diseases: Challenges in developing vaccination strategies. Front. Immunol. 2020, 11, 2130. [Google Scholar] [CrossRef] [PubMed]
- Renner, M.; Dejnirattisai, W.; Carrique, L.; Martin, I.S.; Karia, D.; Ilca, S.L.; Ho, S.F.; Kotecha, A.; Keown, J.R.; Mongkolsapaya, J.; et al. Flavivirus maturation leads to the formation of an occupied lipid pocket in the surface glycoproteins. Nat. Commun. 2021, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergstrom, S.; Vial, L.; Boulanger, N. Relapsing fevers: Neglected tick-borne diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brites-Neto, J.; Duarte, K.M.; Martins, T.F. Tick-borne infections in human and animal population worldwide. Vet. World 2015, 8, 301–315. [Google Scholar] [CrossRef]
- Kenney, J.L.; Anishchenko, M.; Hermance, M.; Romo, H.; Chen, C.I.; Thangamani, S.; Brault, A.C. Generation of a lineage II powassan virus (deer tick virus) cDNA clone: Assessment of flaviviral genetic determinants of tick and mosquito vector competence. Vector Borne Zoonotic Dis. 2018, 18, 371–381. [Google Scholar] [CrossRef]
- Mendoza-Roldan, J.A.; Mendoza-Roldan, M.A.; Otranto, D. Reptile vector-borne diseases of zoonotic concern. Int. J. Parasitol. Parasites Wildl. 2021, 15, 132–142. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile virus and Usutu virus infections in wild and resident birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, K.L.; Hernandez-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet. Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Cook, C.L.; Huang, Y.S.; Lyons, A.C.; Alto, B.W.; Unlu, I.; Higgs, S.; Vanlandingham, D.L. North American Culex pipiens and Culex quinquefasciatus are competent vectors for Usutu virus. PLoS Negl. Trop. Dis. 2018, 12, e0006732. [Google Scholar] [CrossRef] [Green Version]
- Ushijima, Y.; Abe, H.; Nguema Ondo, G.; Bikangui, R.; Massinga Loembe, M.; Zadeh, V.R.; Essimengane, J.G.E.; Mbouna, A.V.N.; Bache, E.B.; Agnandji, S.T.; et al. Surveillance of the major pathogenic arboviruses of public health concern in Gabon, Central Africa: Increased risk of West Nile virus and dengue virus infections. BMC Infect. Dis. 2021, 21, 265. [Google Scholar] [CrossRef]
- Offerdahl, D.K.; Dorward, D.W.; Hansen, B.T.; Bloom, M.E. A three-dimensional comparison of tick-borne flavivirus infection in mammalian and tick cell lines. PLoS ONE 2012, 7, e47912. [Google Scholar] [CrossRef]
- Qiu, Y.; Xu, Y.P.; Wang, M.; Miao, M.; Zhou, H.; Xu, J.; Kong, J.; Zheng, D.; Li, R.T.; Zhang, R.R.; et al. Flavivirus induces and antagonizes antiviral RNA interference in both mammals and mosquitoes. Sci. Adv. 2020, 6, eaax7989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sempere, R.N.; Arias, A. Establishment of a cell culture model of persistent flaviviral infection: Usutu virus shows sustained replication during passages and resistance to extinction by antiviral nucleosides. Viruses 2019, 11, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lannes, N.; Garcia-Nicolas, O.; Demoulins, T.; Summerfield, A.; Filgueira, L. CX3CR1-CX3CL1-dependent cell-to-cell Japanese encephalitis virus transmission by human microglial cells. Sci. Rep. 2019, 9, 4833. [Google Scholar] [CrossRef]
- Wahaab, A.; Liu, K.; Hameed, M.; Anwar, M.N.; Kang, L.; Li, C.; Ma, X.; Wajid, A.; Yang, Y.; Khan, U.H.; et al. Identification of cleavage sites proteolytically processed by NS2B-NS3 protease in polyprotein of Japanese encephalitis virus. Pathogens 2021, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Mottin, M.; Borba, J.; Braga, R.C.; Torres, P.H.M.; Martini, M.C.; Proenca-Modena, J.L.; Judice, C.C.; Costa, F.T.M.; Ekins, S.; Perryman, A.L.; et al. The A-Z of Zika drug discovery. Drug Discov. Today 2018, 23, 1833–1847. [Google Scholar] [CrossRef] [PubMed]
- Shiryaev, S.A.; Strongin, A.Y. Structural and functional parameters of the flaviviral protease: A promising antiviral drug target. Future Virol. 2010, 5, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, Y.Y.; Balasuriya, U.B.; Lee, C.K. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theze, J.; Lowes, S.; Parker, J.; Pybus, O.G. Evolutionary and phylogenetic analysis of the hepaciviruses and pegiviruses. Genome Biol. Evol. 2015, 7, 2996–3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, D.; Mostashari, F.; Fine, A.; Miller, J.; O’Leary, D.; Murray, K.; Huang, A.; Rosenberg, A.; Greenberg, A.; Sherman, M.; et al. The outbreak of West Nile virus infection in the New York City area in 1999. N. Engl. J. Med. 2001, 344, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Swetnam, D.; Widen, S.G.; Wood, T.G.; Reyna, M.; Wilkerson, L.; Debboun, M.; Symonds, D.A.; Mead, D.G.; Beaty, B.J.; Guzman, H.; et al. Terrestrial bird migration and West Nile virus circulation, United States. Emerg. Infect. Dis. 2018, 24, 2184–2194. [Google Scholar] [CrossRef] [Green Version]
- Gwee, S.X.W.; St John, A.L.; Gray, G.C.; Pang, J. Animals as potential reservoirs for dengue transmission: A systematic review. One Health 2021, 12, 100216. [Google Scholar] [CrossRef]
- Robles-Fernandez, A.L.; Santiago-Alarcon, D.; Lira-Noriega, A. American mammals susceptibility to dengue according to geographical, environmental, and phylogenetic distances. Front. Vet. Sci. 2021, 8, 604560. [Google Scholar] [CrossRef] [PubMed]
- Guth, S.; Hanley, K.A.; Althouse, B.M.; Boots, M. Ecological processes underlying the emergence of novel enzootic cycles: Arboviruses in the neotropics as a case study. PLoS Negl. Trop. Dis. 2020, 14, e0008338. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Dos-Santos, T.; Roiz, D.; Lourenco-de-Oliveira, R.; Paupy, C. A systematic review: Is Aedes albopictus an efficient bridge vector for zoonotic arboviruses? Pathogens 2020, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Benzarti, E.; Linden, A.; Desmecht, D.; Garigliany, M. Mosquito-borne epornitic flaviviruses: An update and review. J. Gen. Virol. 2019, 100, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu virus: An arbovirus on the rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guggemos, H.D.; Fendt, M.; Hieke, C.; Heyde, V.; Mfune, J.K.E.; Borgemeister, C.; Junglen, S. Simultaneous circulation of two West Nile virus lineage 2 clades and Bagaza virus in the Zambezi region, Namibia. PLoS Negl. Trop. Dis. 2021, 15, e0009311. [Google Scholar] [CrossRef] [PubMed]
- Mlera, L.; Bloom, M.E. The role of mammalian reservoir hosts in tick-borne flavivirus biology. Front. Cell. Infect. Microbiol. 2018, 8, 298. [Google Scholar] [CrossRef]
- Michelitsch, A.; Wernike, K.; Klaus, C.; Dobler, G.; Beer, M. Exploring the reservoir hosts of tick-borne Encephalitis virus. Viruses 2019, 11, 669. [Google Scholar] [CrossRef] [Green Version]
- Ott, D.; Ulrich, K.; Ginsbach, P.; Ohme, R.; Bock-Hensley, O.; Falk, U.; Teinert, M.; Lenhard, T. Tick-borne encephalitis virus (TBEV) prevalence in field-collected ticks (Ixodes ricinus) and phylogenetic, structural and virulence analysis in a TBE high-risk endemic area in southwestern Germany. Parasites Vectors 2020, 13, 303. [Google Scholar] [CrossRef]
- Bakhvalova, V.N.; Potapova, O.F.; Panov, V.V.; Morozova, O.V. Vertical transmission of tick-borne encephalitis virus between generations of adapted reservoir small rodents. Virus Res. 2009, 140, 172–178. [Google Scholar] [CrossRef]
- Kjaer, L.J.; Klitgaard, K.; Soleng, A.; Edgar, K.S.; Lindstedt, H.E.H.; Paulsen, K.M.; Andreassen, A.K.; Korslund, L.; Kjelland, V.; Slettan, A.; et al. Spatial patterns of pathogen prevalence in questing Ixodes ricinus nymphs in southern Scandinavia, 2016. Sci. Rep. 2020, 10, 19376. [Google Scholar] [CrossRef]
- Grzybek, M.; Alsarraf, M.; Tolkacz, K.; Behnke-Borowczyk, J.; Biernat, B.; Stanczak, J.; Strachecka, A.; Guz, L.; Szczepaniak, K.; Paleolog, J.; et al. Seroprevalence of TBEV in bank voles from Poland-a long-term approach. Emerg. Microbes Infect. 2018, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Corrin, T.; Greig, J.; Harding, S.; Young, I.; Mascarenhas, M.; Waddell, L.A. Powassan virus, a scoping review of the global evidence. Zoonoses Public Health 2018, 65, 595–624. [Google Scholar] [CrossRef]
- Smith, K.; Oesterle, P.T.; Jardine, C.M.; Dibernardo, A.; Huynh, C.; Lindsay, R.; Pearl, D.L.; Bosco-Lauth, A.M.; Nemeth, N.M. Powassan virus and other arthropod-borne viruses in wildlife and ticks in Ontario, Canada. Am. J. Trop. Med. Hyg. 2018, 99, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.M.; Root, J.; Hartwig, A.E.; Bowen, R.A.; Bosco-Lauth, A.M. Powassan virus experimental infections in three wild mammal species. Am. J. Trop. Med. Hyg. 2021, 104, 1048–1054. [Google Scholar] [CrossRef]
- Deardorff, E.R.; Nofchissey, R.A.; Cook, J.A.; Hope, A.G.; Tsvetkova, A.; Talbot, S.L.; Ebel, G.D. Powassan virus in mammals, Alaska and New Mexico, U.S.A. and Russia, 2004–2007. Emerg. Infect. Dis. 2013, 19, 2012–2016. [Google Scholar] [CrossRef]
- Dupuis, A.P., 2nd; Peters, R.J.; Prusinski, M.A.; Falco, R.C.; Ostfeld, R.S.; Kramer, L.D. Isolation of deer tick virus (Powassan virus, lineage II) from Ixodes scapularis and detection of antibody in vertebrate hosts sampled in the Hudson Valley, New York State. Parasites Vectors 2013, 6, 185. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, J.M.; Mlera, L.; Offerdahl, D.K.; VanSickle, M.; Bloom, M.E. Tick-borne flaviviruses depress AKT activity during acute infection by modulating AKT1/2. Viruses 2020, 12, 1059. [Google Scholar] [CrossRef]
- Hermance, M.E.; Hart, C.E.; Esterly, A.T.; Reynolds, E.S.; Bhaskar, J.R.; Thangamani, S. Development of a small animal model for deer tick virus pathogenesis mimicking human clinical outcome. PLoS Negl. Trop. Dis. 2020, 14, e0008359. [Google Scholar] [CrossRef]
- Franzo, G.; Grassi, L.; Tucciarone, C.M.; Drigo, M.; Martini, M.; Pasotto, D.; Mondin, A.; Menandro, M.L. A wild circulation: High presence of Porcine circovirus 3 in different mammalian wild hosts and ticks. Transbound. Emerg. Dis. 2019, 66, 1548–1557. [Google Scholar] [CrossRef]
- Lommano, E.; Dvorak, C.; Vallotton, L.; Jenni, L.; Gern, L. Tick-borne pathogens in ticks collected from breeding and migratory birds in Switzerland. Ticks Tick Borne Dis. 2014, 5, 871–882. [Google Scholar] [CrossRef]
- Weaver, S.C.; Barrett, A.D. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic cycles of arboviruses in non-human primates. Parasites Vectors 2019, 12, 463. [Google Scholar] [CrossRef]
- Nemeth, N.; Bosco-Lauth, A.; Oesterle, P.; Kohler, D.; Bowen, R. North American birds as potential amplifying hosts of Japanese encephalitis virus. Am. J. Trop. Med. Hyg. 2012, 87, 760–767. [Google Scholar] [CrossRef]
- Borah, J.; Dutta, P.; Khan, S.A.; Mahanta, J. Epidemiological concordance of Japanese encephalitis virus infection among mosquito vectors, amplifying hosts and humans in India. Epidemiol. Infect. 2013, 141, 74–80. [Google Scholar] [CrossRef]
- Oliveira, A.R.S.; Cohnstaedt, L.W.; Noronha, L.E.; Mitzel, D.; McVey, D.S.; Cernicchiaro, N. Perspectives regarding the risk of introduction of the Japanese encephalitis virus (JEV) in the United States. Front. Vet. Sci. 2020, 7, 48. [Google Scholar] [CrossRef]
- Yap, G.; Mailepessov, D.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Chong, C.S.; Lam-Phua, S.G.; Lee, R.; et al. Detection of Japanese encephalitis virus in culex mosquitoes in Singapore. Am. J. Trop. Med. Hyg. 2020, 103, 1234–1240. [Google Scholar] [CrossRef]
- Cleton, N.B.; Bosco-Lauth, A.; Page, M.J.; Bowen, R.A. Age-related susceptibility to Japanese encephalitis virus in domestic ducklings and chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef]
- Yap, G.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Mailepessov, D.; Kong, M.; Lai, Y.L.; Okumura, C.; et al. Serological evidence of continued Japanese encephalitis virus transmission in Singapore nearly three decades after end of pig farming. Parasites Vectors 2019, 12, 244. [Google Scholar] [CrossRef] [Green Version]
- Hameed, M.; Wahaab, A.; Nawaz, M.; Khan, S.; Nazir, J.; Liu, K.; Wei, J.; Ma, Z. Potential role of birds in Japanese encephalitis virus zoonotic transmission and genotype shift. Viruses 2021, 13, 357. [Google Scholar] [CrossRef]
- Hegde, N.R.; Gore, M.M. Japanese encephalitis vaccines: Immunogenicity, protective efficacy, effectiveness, and impact on the burden of disease. Hum. Vaccines Immunother. 2017, 13, 1320–1337. [Google Scholar] [CrossRef] [Green Version]
- Schutten, K.; Chabot, A.; Wheeler, H. West Nile virus seroconversion in eastern Loggerhead Shrike (Lanius Ludovicianus Migrans) after vaccination with a killed vaccine. J. Zoo Wildl. Med. 2021, 52, 185–191. [Google Scholar] [CrossRef]
- Malekian, M.; Shagholian, J.; Hosseinpour, Z. Pathogen presence in wild birds inhabiting landfills in central Iran. Ecohealth 2021, 18, 76–83. [Google Scholar] [CrossRef]
- Islam, A.; Islam, S.; Hossain, M.E.; Ferdous, J.; Abedin, J.; Ziaur Rahman, M.; Rahman, M.K.; Hoque, M.A.; Hassan, M.M. Serological evidence of West Nile virus in wild birds in Bangladesh. Vet. Sci. 2020, 7, 164. [Google Scholar] [CrossRef]
- Foppa, I.M.; Beard, R.H.; Mendenhall, I.H. The impact of West Nile virus on the abundance of selected North American birds. BMC Vet. Res. 2011, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, A.M.; Wheeler, S.S. Impact of West Nile Virus on bird populations: Limited lasting effects, evidence for recovery, and gaps in our understanding of impacts on ecosystems. J. Med. Entomol. 2019, 56, 1491–1497. [Google Scholar] [CrossRef]
- McKenzie, V.J.; Goulet, N.E. Bird community composition linked to human West Nile virus cases along the Colorado front range. Ecohealth 2010, 7, 439–447. [Google Scholar] [CrossRef]
- Wheeler, S.S.; Langevin, S.A.; Brault, A.C.; Woods, L.; Carroll, B.D.; Reisen, W.K. Detection of persistent west nile virus RNA in experimentally and naturally infected avian hosts. Am. J. Trop. Med. Hyg. 2012, 87, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.M.; Oesterle, P.T.; Bowen, R.A. Humoral immunity to West Nile virus is long-lasting and protective in the house sparrow (Passer domesticus). Am. J. Trop. Med. Hyg. 2009, 80, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Montecino-Latorre, D.; Barker, C.M. Overwintering of West Nile virus in a bird community with a communal crow roost. Sci. Rep. 2018, 8, 6088. [Google Scholar] [CrossRef] [Green Version]
- Semenov, B.F.; Chunikhin, S.P.; Karmysheva, V.; Iakovleva, N.I. Study of chronic forms of arbovirus infections in birds. 1. Experiments with West Nile, Sindbis, Bhandja and Sicilian mosquito fever viruses. Vestn. Akad. Med. Nauk. SSSR 1973, 28, 79–83. [Google Scholar]
- Mlera, L.; Melik, W.; Bloom, M.E. The role of viral persistence in flavivirus biology. Pathog. Dis. 2014, 71, 137–163. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasites Vectors 2010, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- VanDalen, K.K.; Hall, J.S.; Clark, L.; McLean, R.G.; Smeraski, C. West Nile virus infection in American Robins: New insights on dose response. PLoS ONE 2013, 8, e68537. [Google Scholar] [CrossRef]
- Hannon, E.R.; Jackson, K.C.; Biggerstaff, B.J.; Raman, V.; Komar, N. Bloodmeal host selection of culex quinquefasciatus (Diptera: Culicidae) in Las Vegas, Nevada, United States. J. Med. Entomol. 2019, 56, 603–608. [Google Scholar] [CrossRef]
- Duggal, N.K.; Langwig, K.E.; Ebel, G.D.; Brault, A.C. On the fly: Interactions between birds, mosquitoes, and environment that have molded West Nile virus genomic structure over two decades. J. Med. Entomol. 2019, 56, 1467–1474. [Google Scholar] [CrossRef]
- Steinman, A.; Banet-Noach, C.; Tal, S.; Levi, O.; Simanov, L.; Perk, S.; Malkinson, M.; Shpigel, N. West Nile virus infection in crocodiles. Emerg. Infect. Dis. 2003, 9, 887–889. [Google Scholar] [CrossRef] [Green Version]
- Habarugira, G.; Suen, W.W.; Hobson-Peters, J.; Hall, R.A.; Bielefeldt-Ohmann, H. West Nile virus: An update on pathobiology, epidemiology, diagnostics, control and “One Health” implications. Pathogens 2020, 9, 589. [Google Scholar] [CrossRef]
- Kopitar-Jerala, N. The Role of interferons in inflammation and inflammasome activation. Front. Immunol. 2017, 8, 873. [Google Scholar] [CrossRef] [Green Version]
- Mesev, E.V.; LeDesma, R.A.; Ploss, A. Decoding type I and III interferon signalling during viral infection. Nat. Microbiol. 2019, 4, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.H.; Wang, Y.N.; Chang, Q.Y.; Ma, P.; Hu, Y.; Cao, X. Type III interferons in viral infection and antiviral immunity. Cell. Physiol. Biochem. 2018, 51, 173–185. [Google Scholar] [CrossRef]
- Refolo, G.; Vescovo, T.; Piacentini, M.; Fimia, G.M.; Ciccosanti, F. Mitochondrial interactome: A focus on antiviral signaling pathways. Front. Cell. Dev. Biol. 2020, 8, 8. [Google Scholar] [CrossRef]
- Onomoto, K.; Onoguchi, K.; Yoneyama, M. Regulation of RIG-I-like receptor-mediated signaling: Interaction between host and viral factors. Cell. Mol. Immunol. 2021, 18, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.T.; Lubick, K.J.; Robertson, S.J.; Broughton, J.P.; Bloom, M.E.; Bresnahan, W.A.; Best, S.M. TRIM79alpha, an interferon-stimulated gene product, restricts tick-borne encephalitis virus replication by degrading the viral RNA polymerase. Cell Host Microbe 2011, 10, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.F.; Nisole, S. West Nile virus restriction in mosquito and human cells: A virus under confinement. Vaccines 2020, 8, 256. [Google Scholar] [CrossRef]
- Avirutnan, P.; Fuchs, A.; Hauhart, R.E.; Somnuke, P.; Youn, S.; Diamond, M.S.; Atkinson, J.P. Antagonism of the complement component C4 by flavivirus nonstructural protein NS1. J. Exp. Med. 2010, 207, 793–806. [Google Scholar] [CrossRef] [Green Version]
- Coelho, D.R.; Carneiro, P.H.; Mendes-Monteiro, L.; Conde, J.N.; Andrade, I.; Cao, T.; Allonso, D.; White-Dibiasio, M.; Kuhn, R.J.; Mohana-Borges, R. ApoA1 neutralizes proinflammatory effects of dengue virus NS1 protein and modulates viral immune evasion. J. Virol. 2021, 95, e0197420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Ye, H.Q.; Liu, S.Q.; Deng, C.L.; Li, X.D.; Shi, P.Y.; Zhang, B. West Nile virus NS1 antagonizes interferon beta production by targeting RIG-I and MDA5. J. Virol. 2017, 91, e02396-16. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.A.; Young, P.R. The flavivirus NS1 protein: Molecular and structural biology, immunology, role in pathogenesis and application as a diagnostic biomarker. Antivir. Res. 2013, 98, 192–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, W.L.; Nguyen-Dinh, V.; Larsson, E.; Lindqvist, R.; Overby, A.K.; Lundmark, R. Model system for the formation of tick-borne encephalitis virus replication compartments without viral RNA replication. J. Virol. 2019, 93, e00292-19. [Google Scholar] [CrossRef] [Green Version]
- Domnich, A.; Panatto, D.; Arbuzova, E.K.; Signori, A.; Avio, U.; Gasparini, R.; Amicizia, D. Immunogenicity against Far Eastern and Siberian subtypes of tick-borne encephalitis (TBE) virus elicited by the currently available vaccines based on the European subtype: Systematic review and meta-analysis. Hum. Vaccines Immunother. 2014, 10, 2819–2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.J.; Wang, X.J.; Clark, D.C.; Lobigs, M.; Hall, R.A.; Khromykh, A.A. A single amino acid substitution in the West Nile virus nonstructural protein NS2A disables its ability to inhibit alpha/beta interferon induction and attenuates virus virulence in mice. J. Virol. 2006, 80, 2396–2404. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.; Liew, O.W.; Chen, G.; Chong, P.C.; Lee, S.H.; Chen, K.; Jiang, H.; Puah, C.M.; Zhu, W. Mechanism of NS2B-mediated activation of NS3pro in dengue virus: Molecular dynamics simulations and bioassays. J. Virol. 2009, 83, 1060–1070. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, J.M.; Khromykh, A.A.; Jones, M.K.; Westaway, E.G. Subcellular localization and some biochemical properties of the flavivirus Kunjin nonstructural proteins NS2A and NS4A. Virology 1998, 245, 203–215. [Google Scholar] [CrossRef]
- Venkataraman, S.; Prasad, B.; Selvarajan, R. RNA dependent RNA polymerases: Insights from structure, function and evolution. Viruses 2018, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Siddharthan, V.; Hall, J.O.; Morrey, J.D. Autonomic nervous dysfunction in hamsters infected with West Nile virus. PLoS ONE 2011, 6, e19575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh Roy, S.; Sadigh, B.; Datan, E.; Lockshin, R.A.; Zakeri, Z. Regulation of cell survival and death during Flavivirus infections. World J. Biol. Chem. 2014, 5, 93–105. [Google Scholar] [PubMed]
- Bhatnagar, P.; Sreekanth, G.P.; Murali-Krishna, K.; Chandele, A.; Sitaraman, R. Dengue virus non-structural protein 5 as a versatile, multi-functional effector in host-pathogen interactions. Front. Cell. Infect. Microbiol. 2021, 11, 574067. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.D.; Yeh, W.T.; Chen, R.F.; Chuon, H.L.; Tsai, H.P.; Yao, C.W.; Shaio, M.F. A model to study neurotropism and persistency of Japanese encephalitis virus infection in human neuroblastoma cells and leukocytes. J. Gen. Virol. 2004, 85, 635–642. [Google Scholar] [CrossRef]
- Lin, C.W.; Lin, K.H.; Lyu, P.C.; Chen, W.J. Japanese encephalitis virus NS2B-NS3 protease binding to phage-displayed human brain proteins with the domain of trypsin inhibitor and basic region leucine zipper. Virus Res. 2006, 116, 106–113. [Google Scholar] [CrossRef]
- Pan, Y.; Cheng, A.; Wang, M.; Yin, Z.; Jia, R. The dual regulation of apoptosis by flavivirus. Front. Microbiol. 2021, 12, 654494. [Google Scholar] [CrossRef] [PubMed]
- Prikhod’ko, G.G.; Prikhod’ko, E.A.; Pletnev, A.G.; Cohen, J.I. Langat flavivirus protease NS3 binds caspase-8 and induces apoptosis. J. Virol. 2002, 76, 5701–5710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roosendaal, J.; Westaway, E.G.; Khromykh, A.; Mackenzie, J.M. Regulated cleavages at the West Nile virus NS4A-2K-NS4B junctions play a major role in rearranging cytoplasmic membranes and Golgi trafficking of the NS4A protein. J. Virol. 2006, 80, 4623–4632. [Google Scholar] [CrossRef] [Green Version]
- Brisse, M.; Ly, H. Comparative structure and function analysis of the RIG-I-like receptors: RIG-I and MDA5. Front. Immunol. 2019, 10, 1586. [Google Scholar] [CrossRef] [PubMed]
- McLean, J.E.; Wudzinska, A.; Datan, E.; Quaglino, D.; Zakeri, Z. Flavivirus NS4A-induced autophagy protects cells against death and enhances virus replication. J. Biol. Chem. 2011, 286, 22147–22159. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Wang, X.J.; Mokhonov, V.V.; Shi, P.Y.; Randall, R.; Khromykh, A.A. Inhibition of interferon signaling by the New York 99 strain and Kunjin subtype of West Nile virus involves blockage of STAT1 and STAT2 activation by nonstructural proteins. J. Virol. 2005, 79, 1934–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz-Jordan, J.L.; Sanchez-Burgos, G.G.; Laurent-Rolle, M.; Garcia-Sastre, A. Inhibition of interferon signaling by dengue virus. Proc. Natl. Acad. Sci. USA 2003, 100, 14333–14338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.J.; Chang, B.L.; Yu, H.P.; Liao, C.L.; Lin, Y.L. Blocking of interferon-induced Jak-Stat signaling by Japanese encephalitis virus NS5 through a protein tyrosine phosphatase-mediated mechanism. J. Virol. 2006, 80, 5908–5918. [Google Scholar] [CrossRef] [Green Version]
- Mazzon, M.; Jones, M.; Davidson, A.; Chain, B.; Jacobs, M. Dengue virus NS5 inhibits interferon-alpha signaling by blocking signal transducer and activator of transcription 2 phosphorylation. J. Infect. Dis. 2009, 200, 1261–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, S.M.; Morris, K.L.; Shannon, J.G.; Robertson, S.J.; Mitzel, D.N.; Park, G.S.; Boer, E.; Wolfinbarger, J.B.; Bloom, M.E. Inhibition of interferon-stimulated JAK-STAT signaling by a tick-borne flavivirus and identification of NS5 as an interferon antagonist. J. Virol. 2005, 79, 12828–12839. [Google Scholar] [CrossRef] [Green Version]
- Park, G.S.; Morris, K.L.; Hallett, R.G.; Bloom, M.E.; Best, S.M. Identification of residues critical for the interferon antagonist function of Langat virus NS5 reveals a role for the RNA-dependent RNA polymerase domain. J. Virol. 2007, 81, 6936–6946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werme, K.; Wigerius, M.; Johansson, M. Tick-borne encephalitis virus NS5 associates with membrane protein scribble and impairs interferon-stimulated JAK-STAT signalling. Cell. Microbiol. 2008, 10, 696–712. [Google Scholar] [CrossRef]
- Rastogi, M.; Sharma, N.; Singh, S.K. Flavivirus NS1: A multifaceted enigmatic viral protein. Virol. J. 2016, 13, 131. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenko, Y.V.; Starodubova, E.S.; Shevtsova, A.S.; Chernokhaeva, L.L.; Latanova, A.A.; Preobrazhenskaia, O.V.; Timofeev, A.V.; Karganova, G.G.; Karpov, V.L. Intracellular degradation and localization of NS1 of tick-borne encephalitis virus affect its protective properties. J. Gen. Virol. 2017, 98, 50–55. [Google Scholar] [CrossRef]
- Rosales Ramirez, R.; Ludert, J.E. The dengue virus nonstructural protein 1 (NS1) is secreted from mosquito cells in association with the intracellular cholesterol transporter chaperone caveolin complex. J. Virol. 2019, 93, e01985-18. [Google Scholar] [CrossRef] [Green Version]
- Edeling, M.A.; Diamond, M.S.; Fremont, D.H. Structural basis of Flavivirus NS1 assembly and antibody recognition. Proc. Natl. Acad. Sci. USA 2014, 111, 4285–4290. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.R.; Lai, Y.C.; Yeh, T.M. Dengue virus non-structural protein 1: A pathogenic factor, therapeutic target, and vaccine candidate. J. Biomed. Sci. 2018, 25, 58. [Google Scholar] [CrossRef] [PubMed]
- Mehrbod, P.; Ande, S.R.; Alizadeh, J.; Rahimizadeh, S.; Shariati, A.; Malek, H.; Hashemi, M.; Glover, K.K.M.; Sher, A.A.; Coombs, K.M.; et al. The roles of apoptosis, autophagy and unfolded protein response in arbovirus, influenza virus, and HIV infections. Virulence 2019, 10, 376–413. [Google Scholar] [CrossRef] [Green Version]
- Rego, R.O.M.; Trentelman, J.J.A.; Anguita, J.; Nijhof, A.M.; Sprong, H.; Klempa, B.; Hajdusek, O.; Tomas-Cortazar, J.; Azagi, T.; Strnad, M.; et al. Counterattacking the tick bite: Towards a rational design of anti-tick vaccines targeting pathogen transmission. Parasites Vectors 2019, 12, 229. [Google Scholar] [CrossRef]
- Wang, X.; Chang, C.H.; Jiang, J.; Liu, X.; Li, J.; Liu, Q.; Liao, Y.P.; Li, L.; Nel, A.E.; Xia, T. Mechanistic differences in cell death responses to metal-based engineered nanomaterials in kupffer cells and hepatocytes. Small 2020, 16, e2000528. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, M.P.; Chambers, J.A.; Pankhong, P.; Chattergoon, M.; Attatippaholkun, W.; Dang, K.; Shah, N.; Weiner, D.B. Host cell killing by the West Nile Virus NS2B-NS3 proteolytic complex: NS3 alone is sufficient to recruit caspase-8-based apoptotic pathway. Virology 2006, 345, 56–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miorin, L.; Maestre, A.M.; Fernandez-Sesma, A.; Garcia-Sastre, A. Antagonism of type I interferon by flaviviruses. Biochem. Biophys. Res. Commun. 2017, 492, 587–596. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Aktepe, T.E.; Mackenzie, J.M. Shaping the flavivirus replication complex: It is curvaceous! Cell. Microbiol. 2018, 20, e12884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.; Li, M.M.H. All about the RNA: Interferon-stimulated genes that interfere with viral RNA processes. Front. Immunol. 2020, 11, 605024. [Google Scholar] [CrossRef]
- Thurmond, S.; Wang, B.; Song, J.; Hai, R. Suppression of type I interferon signaling by flavivirus NS5. Viruses 2018, 10, 712. [Google Scholar] [CrossRef] [Green Version]
- Slon Campos, J.L.; Mongkolsapaya, J.; Screaton, G.R. The immune response against flaviviruses. Nat. Immunol. 2018, 19, 1189–1198. [Google Scholar] [CrossRef]
- Rathore, A.P.S.; St John, A.L. Cross-reactive immunity among flaviviruses. Front. Immunol. 2020, 11, 334. [Google Scholar] [CrossRef] [Green Version]
- Stonedahl, S.; Clarke, P.; Tyler, K.L. The role of microglia during West Nile virus infection of the central nervous system. Vaccines 2020, 8, 485. [Google Scholar] [CrossRef]
- Winkelmann, E.R.; Luo, H.; Wang, T. West Nile virus infection in the central nervous system. F1000Research 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Engle, M.J.; Diamond, M.S. Antibody prophylaxis and therapy against West Nile virus infection in wild-type and immunodeficient mice. J. Virol. 2003, 77, 12941–12949. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Xu, Q.; Nelson, S.; Oliphant, T.; Nybakken, G.E.; Fremont, D.H.; Diamond, M.S. The stoichiometry of antibody-mediated neutralization and enhancement of West Nile virus infection. Cell Host Microbe 2007, 1, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Ye, P.; Wang, X.; Xu, X.; Reisen, W. Real-time monitoring of flavivirus induced cytopathogenesis using cell electric impedance technology. J. Virol. Methods 2011, 173, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.H.; Huang, L.M.; Wu, S.C. A quantitative luciferase-based cell-cell fusion assay to measure four-serotype dengue virus E protein-triggered membrane fusion. Hum. Vaccin. Immunother. 2020, 16, 2176–2182. [Google Scholar] [CrossRef]
- Eyre, N.S.; Johnson, S.M.; Eltahla, A.A.; Aloi, M.; Aloia, A.L.; McDevitt, C.A.; Bull, R.A.; Beard, M.R. Genome-wide mutagenesis of dengue virus reveals plasticity of the NS1 protein and enables generation of infectious tagged reporter viruses. J. Virol. 2017, 91, e01455-17. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.H.; Tsai, M.H.; Tsai, K.N.; Tian, J.N.; Wu, J.S.; Wu, S.Y.; Chern, J.H.; Chen, C.H.; Yueh, A. Mutagenesis of dengue virus protein NS2A revealed a novel domain responsible for virus-induced cytopathic effect and interactions between NS2A and NS2B transmembrane segments. J. Virol. 2017, 91, e01836-16. [Google Scholar] [CrossRef] [Green Version]
- Shafee, N.; AbuBakar, S. Dengue virus type 2 NS3 protease and NS2B-NS3 protease precursor induce apoptosis. J. Gen. Virol. 2003, 84, 2191–2195. [Google Scholar] [CrossRef] [PubMed]
- Vicenzi, E.; Pagani, I.; Ghezzi, S.; Taylor, S.L.; Rudd, T.R.; Lima, M.A.; Skidmore, M.A.; Yates, E.A. Subverting the mechanisms of cell death: Flavivirus manipulation of host cell responses to infection. Biochem. Soc. Trans. 2018, 46, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Puerta-Guardo, H.; Biering, S.B.; Glasner, D.R.; Tran, E.B.; Patana, M.; Gomberg, T.A.; Malvar, C.; Lo, N.T.N.; Espinosa, D.A.; et al. Endocytosis of flavivirus NS1 is required for NS1-mediated endothelial hyperpermeability and is abolished by a single N-glycosylation site mutation. PLoS Pathog. 2019, 15, e1007938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, C.M.R.; Sevvana, M.; Kuhn, R.J. Structure-guided paradigm shifts in flavivirus assembly and maturation mechanisms. Adv. Virus Res. 2020, 108, 33–83. [Google Scholar]
- Lee, C.J.; Liao, C.L.; Lin, Y.L. Flavivirus activates phosphatidylinositol 3-kinase signaling to block caspase-dependent apoptotic cell death at the early stage of virus infection. J. Virol. 2005, 79, 8388–8399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; He, Y.; Xu, Y.; Mo, F.; Mi, T.; Shen, Q.S.; Li, C.; Li, Y.; Liu, J.; Wu, Y.; et al. Differential antiviral immunity to Japanese encephalitis virus in developing cortical organoids. Cell Death Dis. 2018, 9, 719. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, L.; Fikrig, E.; Wang, P. An essential role of PI3K in the control of West Nile virus infection. Sci. Rep. 2017, 7, 3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoni, T.B.; Lopez, C.B. Defective (interfering) viral genomes re-explored: Impact on antiviral immunity and virus persistence. Future Virol. 2018, 13, 493–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, N.C.; Tiernery, R.; Sil, B.K.; Wills, M.R.; Barrett, A.D. In vitro homotypic and heterotypic interference by defective interfering particles of West Nile virus. J. Gen. Virol. 1991, 72, 2705–2711. [Google Scholar] [CrossRef]
- Vignuzzi, M.; Lopez, C.B. Defective viral genomes are key drivers of the virus-host interaction. Nat. Microbiol. 2019, 4, 1075–1087. [Google Scholar] [CrossRef]
- Li, D.; Lott, W.B.; Lowry, K.; Jones, A.; Thu, H.M.; Aaskov, J. Defective interfering viral particles in acute dengue infections. PLoS ONE 2011, 6, e19447. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Lyu, T.; Zhou, R.; He, X.; Ye, K.; Xie, Q.; Zhu, L.; Chen, T.; Shen, C.; Wu, Q.; et al. The antiviral and antitumor effects of defective interfering particles/genomes and their mechanisms. Front. Microbiol. 2019, 10, 1852. [Google Scholar] [CrossRef] [Green Version]
- Bugrysheva, J.V.; Matveeva, V.A.; Dobrikova, E.Y.; Bykovskaya, N.V.; Korobova, S.A.; Bakhvalova, V.N.; Morozova, O.V. Tick-borne encephalitis virus NS1 glycoprotein during acute and persistent infection of cells. Virus Res. 2001, 76, 161–169. [Google Scholar] [CrossRef]
- Mlera, L.; Offerdahl, D.K.; Martens, C.; Porcella, S.F.; Melik, W.; Bloom, M.E. Development of a model system for tick-borne flavivirus persistence in HEK 293T cells. mBio 2015, 6, e00614. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.U.; Hodgetts, S.I.; Mackenzie, J.S.; Urosevic, N. Characterization of defective viral RNA produced during persistent infection of Vero cells with Murray Valley encephalitis virus. J. Virol. 1998, 72, 2474–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poidinger, M.; Coelen, R.J.; Mackenzie, J.S. Persistent infection of Vero cells by the flavivirus Murray Valley encephalitis virus. J. Gen. Virol. 1991, 72, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Pesko, K.N.; Fitzpatrick, K.A.; Ryan, E.M.; Shi, P.Y.; Zhang, B.; Lennon, N.J.; Newman, R.M.; Henn, M.R.; Ebel, G.D. Internally deleted WNV genomes isolated from exotic birds in New Mexico: Function in cells, mosquitoes, and mice. Virology 2012, 427, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Conway, M.J.; Montgomery, R.R.; Fikrig, E. West Nile Virus: Biology, transmission, and human infection. Clin. Microbiol. Rev. 2012, 25, 635–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Di, D.; Huang, H.; Wang, X.; Xia, Q.; Ma, X.; Liu, K.; Li, B.; Shao, D.; Qiu, Y.; et al. NS5-V372A and NS5-H386Y variations are responsible for differences in interferon alpha/beta induction and co-contribute to the replication advantage of Japanese encephalitis virus genotype I over genotype III in ducklings. PLoS Pathog. 2020, 16, e1008773. [Google Scholar] [CrossRef]
- Sanchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito’s RNA interference pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessions, O.M.; Wilm, A.; Kamaraj, U.S.; Choy, M.M.; Chow, A.; Chong, Y.; Ong, X.M.; Nagarajan, N.; Cook, A.R.; Ooi, E.E. Analysis of dengue virus genetic diversity during human and mosquito infection reveals genetic constraints. PLoS Negl. Trop. Dis. 2015, 9, e0004044. [Google Scholar] [CrossRef]
- Espada-Murao, L.A.; Morita, K. Delayed cytosolic exposure of Japanese encephalitis virus double-stranded RNA impedes interferon activation and enhances viral dissemination in porcine cells. J. Virol. 2011, 85, 6736–6749. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.Y.; Hsu, T.W.; Chen, Y.L.; Liu, S.F.; Tsai, Y.J.; Lin, Y.T.; Chen, Y.S.; Fan, Y.H. Japanese encephalitis virus non-coding RNA inhibits activation of interferon by blocking nuclear translocation of interferon regulatory factor 3. Vet. Microbiol. 2013, 166, 11–21. [Google Scholar] [CrossRef]
- Singh, H.; Koury, J.; Kaul, M. Innate immune sensing of viruses and its consequences for the central nervous system. Viruses 2021, 13, 170. [Google Scholar] [CrossRef]
- Overby, A.K.; Popov, V.L.; Niedrig, M.; Weber, F. Tick-borne encephalitis virus delays interferon induction and hides its double-stranded RNA in intracellular membrane vesicles. J. Virol. 2010, 84, 8470–8483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.L.; Lin, Y.L.; Shen, S.C.; Shen, J.Y.; Su, H.L.; Huang, Y.L.; Ma, S.H.; Sun, Y.C.; Chen, K.P.; Chen, L.K. Antiapoptotic but not antiviral function of human bcl-2 assists establishment of Japanese encephalitis virus persistence in cultured cells. J. Virol. 1998, 72, 9844–9854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.J.; Yu, H.P.; Chang, B.L.; Tang, W.C.; Liao, C.L.; Lin, Y.L. Distinct antiviral roles for human 2′,5′-oligoadenylate synthetase family members against dengue virus infection. J. Immunol. 2009, 183, 8035–8043. [Google Scholar] [CrossRef] [Green Version]
- Choi, U.Y.; Kang, J.S.; Hwang, Y.S.; Kim, Y.J. Oligoadenylate synthase-like (OASL) proteins: Dual functions and associations with diseases. Exp. Mol. Med. 2015, 47, e144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinton, M.A. Genetic resistance to flaviviruses. Monoclon. Antibodies Immunodiagn. Immunother. 2014, 33, 183–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangster, M.Y.; Heliams, D.B.; MacKenzie, J.S.; Shellam, G.R. Genetic studies of flavivirus resistance in inbred strains derived from wild mice: Evidence for a new resistance allele at the flavivirus resistance locus (Flv). J. Virol. 1993, 67, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Elbahesh, H.; Jha, B.K.; Silverman, R.H.; Scherbik, S.V.; Brinton, M.A. The Flvr-encoded murine oligoadenylate synthetase 1b (Oas1b) suppresses 2-5A synthesis in intact cells. Virology 2011, 409, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura-Neto, J.A.; Braga Silva, C.A.; Moura, A.F.; Rocco Suassuna, J.H. Emergent arboviruses and renal transplantation: A global challenge. Kidney Int. Rep. 2019, 4, 647–655. [Google Scholar] [CrossRef] [Green Version]
- Appler, K.K.; Brown, A.N.; Stewart, B.S.; Behr, M.J.; Demarest, V.L.; Wong, S.J.; Bernard, K.A. Persistence of West Nile virus in the central nervous system and periphery of mice. PLoS ONE 2010, 5, e10649. [Google Scholar] [CrossRef] [PubMed]
- Garber, C.; Soung, A.; Vollmer, L.L.; Kanmogne, M.; Last, A.; Brown, J.; Klein, R.S. T cells promote microglia-mediated synaptic elimination and cognitive dysfunction during recovery from neuropathogenic flaviviruses. Nat. Neurosci. 2019, 22, 1276–1288. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Samuel, M.A.; Diamond, M.S. CD8+ T cells require perforin to clear West Nile virus from infected neurons. J. Virol. 2006, 80, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.J.; Mitzel, D.N.; Taylor, R.T.; Best, S.M.; Bloom, M.E. Tick-borne flaviviruses: Dissecting host immune responses and virus countermeasures. Immunol. Res. 2009, 43, 172–186. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, S.G.; Murphy, F.A.; Sudia, W.D. St. Louis encephalitis virus: An ultrastructural study of infection in a mosquito vector. Virology 1973, 56, 70–87. [Google Scholar] [CrossRef]
- Brackney, D.E.; Foy, B.D.; Olson, K.E. The effects of midgut serine proteases on dengue virus type 2 infectivity of Aedes aegypti. Am. J. Trop. Med. Hyg. 2008, 79, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; De Chesse, R.; Belhouchet, M.; Lemasson, J.J.; de Micco, P.; de Lamballerie, X. Sequences of flavivirus-related RNA viruses persist in DNA form integrated in the genome of Aedes spp. mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Tsai, K.N.; Chen, Y.S.; Chang, R.Y. Differential miRNA expression profiling reveals correlation of miR125b-5p with persistent infection of Japanese encephalitis virus. Int. J. Mol. Sci. 2021, 22, 4218. [Google Scholar] [CrossRef]
- Vaumourin, E.; Vourc’h, G.; Gasqui, P.; Vayssier-Taussat, M. The importance of multiparasitism: Examining the consequences of co-infections for human and animal health. Parasites Vectors 2015, 8, 545. [Google Scholar] [CrossRef]
- Petney, T.N.; Andrews, R.H. Multiparasite communities in animals and humans: Frequency, structure and pathogenic significance. Int. J. Parasitol. 1998, 28, 377–393. [Google Scholar] [CrossRef]
- Bordes, F.; Morand, S. The impact of multiple infections on wild animal hosts: A review. Infect. Ecol. Epidemiol. 2011, 1, 7346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzauti, M.; Galan, M.; Bernard, M.; Maman, S.; Klopp, C.; Charbonnel, N.; Vayssier-Taussat, M.; Eloit, M.; Cosson, J.F. A comparison between transcriptome sequencing and 16S metagenomics for detection of bacterial pathogens in wildlife. PLoS Negl. Trop. Dis. 2015, 9, e0003929. [Google Scholar] [CrossRef] [Green Version]
- Belak, S.; Karlsson, O.E.; Leijon, M.; Granberg, F. High-throughput sequencing in veterinary infection biology and diagnostics. Rev. Sci. Tech. 2013, 32, 893–915. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Bocanegra, I.; Paniagua, J.; Gutierrez-Guzman, A.V.; Lecollinet, S.; Boadella, M.; Arenas-Montes, A.; Cano-Terriza, D.; Lowenski, S.; Gortazar, C.; Hofle, U. Spatio-temporal trends and risk factors affecting West Nile virus and related flavivirus exposure in Spanish wild ruminants. BMC Vet. Res. 2016, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Chaisiri, K.; Chou, M.; Siew, C.C.; Morand, S.; Ribas, A. Gastrointestinal helminth fauna of rodents from Cambodia: Emphasizing the community ecology of host-parasite associations. J. Helminthol. 2017, 91, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Ezenwa, V.O. Helminth-microparasite co-infection in wildlife: Lessons from ruminants, rodents and rabbits. Parasite Immunol. 2016, 38, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef]
- Kading, R.C.; Borland, E.M.; Cranfield, M.; Powers, A.M. Prevalence of antibodies to alphaviruses and flaviviruses in free-ranging game animals and nonhuman primates in the greater Congo basin. J. Wildl. Dis. 2013, 49, 587–599. [Google Scholar] [CrossRef] [Green Version]
- Klaus, C.; Ziegler, U.; Hoffmann, D.; Press, F.; Fast, C.; Beer, M. Tick-borne encephalitis virus (TBEV) antibodies in animal sera—Occurrence in goat flocks in Germany, longevity and ability to recall immunological information after more than six years. BMC Vet. Res. 2019, 15, 399. [Google Scholar] [CrossRef]
- Bewley, K.R.; Coombes, N.S.; Gagnon, L.; McInroy, L.; Baker, N.; Shaik, I.; St-Jean, J.R.; St-Amant, N.; Buttigieg, K.R.; Humphries, H.E.; et al. Quantification of SARS-CoV-2 neutralizing antibody by wild-type plaque reduction neutralization, microneutralization and pseudotyped virus neutralization assays. Nat. Protoc. 2021, 16, 3114–3140. [Google Scholar] [CrossRef]
- Badolato-Correa, J.; Sanchez-Arcila, J.C.; Alves de Souza, T.M.; Santos Barbosa, L.; Conrado Guerra Nunes, P.; da Rocha Queiroz Lima, M.; Gandini, M.; Bispo de Filippis, A.M.; Venancio da Cunha, R.; Leal de Azeredo, E.; et al. Human T cell responses to Dengue and Zika virus infection compared to Dengue/Zika coinfection. Immun. Inflamm. Dis. 2018, 6, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Zanini, F.; Pu, S.Y.; Bekerman, E.; Einav, S.; Quake, S.R. Single-cell transcriptional dynamics of flavivirus infection. eLife 2018, 7, e32942. [Google Scholar] [CrossRef] [Green Version]
- Hollidge, B.S.; Weiss, S.R.; Soldan, S.S. The role of interferon antagonist, non-structural proteins in the pathogenesis and emergence of arboviruses. Viruses 2011, 3, 629–658. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.D.; Michel, F.; Wylezich, C.; Hoper, D.; Keller, M.; Holicki, C.M.; Szentiks, C.A.; Eiden, M.; Muluneh, A.; Neubauer-Juric, A.; et al. Co-infections: Simultaneous detections of West Nile virus and Usutu virus in birds from Germany. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cacciotti, G.; Caputo, B.; Selvaggi, C.; la Sala, A.; Vitiello, L.; Diallo, D.; Ceianu, C.; Antonelli, G.; Nowotny, N.; Scagnolari, C. Variation in interferon sensitivity and induction between Usutu and West Nile (lineages 1 and 2) viruses. Virology 2015, 485, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Kanthong, N.; Khemnu, N.; Pattanakitsakul, S.N.; Malasit, P.; Flegel, T.W. Persistent, triple-virus co-infections in mosquito cells. BMC Microbiol. 2010, 10, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogels, C.B.F.; Ruckert, C.; Cavany, S.M.; Perkins, T.A.; Ebel, G.D.; Grubaugh, N.D. Arbovirus coinfection and co-transmission: A neglected public health concern? PLoS Biol. 2019, 17, e3000130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Zhang, Z.R.; Zhang, Y.N.; Liu, J.; Deng, C.L.; Shi, P.Y.; Yuan, Z.M.; Ye, H.Q.; Zhang, B. A replication-defective Japanese encephalitis virus (JEV) vaccine candidate with NS1 deletion confers dual protection against JEV and West Nile virus in mice. NPJ Vaccines 2020, 5, 73. [Google Scholar] [CrossRef]
- Supramaniam, A.; Lui, H.; Bellette, B.M.; Rudd, P.A.; Herrero, L.J. How myeloid cells contribute to the pathogenesis of prominent emerging zoonotic diseases. J. Gen. Virol. 2018, 99, 953–969. [Google Scholar] [CrossRef]
- Ma, Y.; Liang, Y.; Wang, N.; Cui, L.; Chen, Z.; Wu, H.; Zhu, C.; Wang, Z.; Liu, S.; Li, H. Avian flavivirus infection of monocytes/macrophages by extensive subversion of host antiviral innate immune responses. J. Virol. 2019, 93, e00978-19. [Google Scholar] [CrossRef]
- Hermance, M.E.; Thangamani, S. Powassan virus: An emerging arbovirus of public health concern in North America. Vector Borne Zoonotic Dis. 2017, 17, 453–462. [Google Scholar] [CrossRef]
- Meurens, F.; Dunoyer, C.; Fourichon, C.; Gerdts, V.; Haddad, N.; Kortekaas, J.; Lewandowska, M.; Monchatre-Leroy, E.; Summerfield, A.; Wichgers Schreur, P.J.; et al. Animal board invited review: Risks of zoonotic disease emergence at the interface of wildlife and livestock systems. Animal 2021, 15, 100241. [Google Scholar] [CrossRef]
- Price, D.P.; Nagarajan, V.; Churbanov, A.; Houde, P.; Milligan, B.; Drake, L.L.; Gustafson, J.E.; Hansen, I.A. The fat body transcriptomes of the yellow fever mosquito Aedes aegypti, pre- and post-blood meal. PLoS ONE 2011, 6, e22573. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.F.; Serrao, J.E.; Ramalho-Ortigao, J.M.; Pimenta, P.F. A comparative study of fat body morphology in five mosquito species. Mem. Inst. Oswaldo Cruz 2011, 106, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.D.; Shapiro, L.L.M.; Thompson, G.A.; Estevez-Lao, T.Y.; Hillyer, J.F. Transstadial immune activation in a mosquito: Adults that emerge from infected larvae have stronger antibacterial activity in their hemocoel yet increased susceptibility to malaria infection. Ecol. Evol. 2019, 9, 6082–6095. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Klingler, K.A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector Borne Zoonotic Dis. 2004, 4, 109–122. [Google Scholar] [CrossRef]
- Linthicum, K.J.; Platt, K.; Myint, K.S.; Lerdthusnee, K.; Innis, B.L.; Vaughn, D.W. Dengue 3 virus distribution in the mosquito Aedes aegypti: An immunocytochemical study. Med. Vet. Entomol. 1996, 10, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sanchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zheng, X.; Wu, Y.; Gan, M.; He, A.; Li, Z.; Zhang, D.; Wu, X.; Zhan, X. Differential proteomics of Aedes albopictus salivary gland, midgut and C6/36 cell induced by dengue virus infection. Virology 2013, 444, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.; Dimopoulos, G. Dengue virus inhibits immune responses in Aedes aegypti cells. PLoS ONE 2010, 5, e10678. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, D.; Das, S.; Begum, F.; Mal, S.; Ray, U. The mosquito immune system and the life of dengue virus: What we know and do not know. Pathogens 2019, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.R.; Zimmermann, L.L.; Crawford, E.D.; Sample, H.A.; Soni, P.R.; Baker, A.N.; Khan, L.M.; DeRisi, J.L. Acute West Nile Virus meningoencephalitis diagnosed via metagenomic deep sequencing of cerebrospinal fluid in a renal transplant patient. Am. J. Transplant. 2017, 17, 803–808. [Google Scholar] [CrossRef]
- Oliveira, E.R.A.; Povoa, T.F.; Nuovo, G.J.; Allonso, D.; Salomao, N.G.; Basilio-de-Oliveira, C.A.; Geraldo, L.H.M.; Fonseca, C.G.; Lima, F.R.S.; Mohana-Borges, R.; et al. Dengue fatal cases present virus-specific HMGB1 response in peripheral organs. Sci. Rep. 2017, 7, 16011. [Google Scholar] [CrossRef] [Green Version]
- Drury, S.E.; Gough, R.E.; McArthur, S.D. Detection and isolation of a flavivirus-like agent from a leopard tortoise (Geochelone paradalis) in the United Kingdom. Vet. Rec. 2001, 148, 452. [Google Scholar] [CrossRef] [PubMed]
- Tonteri, E.; Kipar, A.; Voutilainen, L.; Vene, S.; Vaheri, A.; Vapalahti, O.; Lundkvist, A. The three subtypes of tick-borne encephalitis virus induce encephalitis in a natural host, the bank vole (Myodes glareolus). PLoS ONE 2013, 8, e81214. [Google Scholar] [CrossRef]
- Michelitsch, A.; Fast, C.; Sick, F.; Tews, B.A.; Stiasny, K.; Bestehorn-Willmann, M.; Dobler, G.; Beer, M.; Wernike, K. Long-term presence of tick-borne encephalitis virus in experimentally infected bank voles (Myodes glareolus). Ticks Tick Borne Dis. 2021, 12, 101693. [Google Scholar] [CrossRef]
- Makenov, M.; Karan, L.; Shashina, N.; Akhmetshina, M.; Zhurenkova, O.; Kholodilov, I.; Karganova, G.; Smirnova, N.; Grigoreva, Y.; Yankovskaya, Y.; et al. First detection of tick-borne encephalitis virus in Ixodes ricinus ticks and their rodent hosts in Moscow, Russia. Ticks Tick Borne Dis. 2019, 10, 101265. [Google Scholar] [CrossRef]
- Nagy, A.; Nagy, O.; Tarcsai, K.; Farkas, A.; Takacs, M. First detection of tick-borne encephalitis virus RNA in clinical specimens of acutely ill patients in Hungary. Ticks Tick Borne Dis. 2018, 9, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Caputo, B.; Russo, G.; Manica, M.; Vairo, F.; Poletti, P.; Guzzetta, G.; Merler, S.; Scagnolari, C.; Solimini, A. A comparative analysis of the 2007 and 2017 Italian chikungunya outbreaks and implication for public health response. PLoS Negl. Trop. Dis. 2020, 14, e0008159. [Google Scholar] [CrossRef] [PubMed]
- Ackermann-Gaumann, R.; Tritten, M.L.; Hassan, M.; Lienhard, R. Comparison of three commercial IgG and IgM ELISA kits for the detection of tick-borne encephalitis virus antibodies. Ticks Tick Borne Dis. 2018, 9, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Niklasson, B.; Peters, C.J.; Grandien, M.; Wood, O. Detection of human immunoglobulins G and M antibodies to Rift Valley fever virus by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1984, 19, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khristunova, E.; Barek, J.; Kratochvil, B.; Korotkova, E.; Dorozhko, E.; Vyskocil, V. Electrochemical immunoassay for the detection of antibodies to tick-borne encephalitis virus by using various types of bioconjugates based on silver nanoparticles. Bioelectrochemistry 2020, 135, 107576. [Google Scholar] [CrossRef] [PubMed]
- Korimbocus, J.; Scaramozzino, N.; Lacroix, B.; Crance, J.M.; Garin, D.; Vernet, G. DNA probe array for the simultaneous identification of herpesviruses, enteroviruses, and flaviviruses. J. Clin. Microbiol. 2005, 43, 3779–3787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reta, D.H.; Tessema, T.S.; Ashenef, A.S.; Desta, A.F.; Labisso, W.L.; Gizaw, S.T.; Abay, S.M.; Melka, D.S.; Reta, F.A. Molecular and immunological diagnostic techniques of medical viruses. Int. J. Microbiol. 2020, 2020, 8832728. [Google Scholar] [CrossRef]
- Boriskin, Y.S.; Rice, P.S.; Stabler, R.A.; Hinds, J.; Al-Ghusein, H.; Vass, K.; Butcher, P.D. DNA microarrays for virus detection in cases of central nervous system infection. J. Clin. Microbiol. 2004, 42, 5811–5818. [Google Scholar] [CrossRef] [Green Version]
- Grubaugh, N.D.; Petz, L.N.; Melanson, V.R.; McMenamy, S.S.; Turell, M.J.; Long, L.S.; Pisarcik, S.E.; Kengluecha, A.; Jaichapor, B.; O’Guinn, M.L.; et al. Evaluation of a field-portable DNA microarray platform and nucleic acid amplification strategies for the detection of arboviruses, arthropods, and bloodmeals. Am. J. Trop. Med. Hyg. 2013, 88, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Qin, C.; Li, Y.; Liu, H.; Lin, F.; Li, Y.; Li, J.; Zhu, Q.; Yang, Y. Improvement of the specificity of a pan-viral microarray by using genus-specific oligonucleotides and reduction of interference by host genomes. J. Med. Virol. 2011, 83, 1624–1630. [Google Scholar] [CrossRef]
- Yu, A.C.; Vatcher, G.; Yue, X.; Dong, Y.; Li, M.H.; Tam, P.H.; Tsang, P.Y.; Wong, A.K.; Hui, M.H.; Yang, B.; et al. Nucleic acid-based diagnostics for infectious diseases in public health affairs. Front. Med. 2012, 6, 173–186. [Google Scholar] [CrossRef]
- Bexfield, N.; Kellam, P. Metagenomics and the molecular identification of novel viruses. Vet. J. 2011, 190, 191–198. [Google Scholar] [CrossRef]
- Bumgarner, R. Overview of DNA microarrays: Types, applications, and their future. Curr. Protoc. Mol. Biol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Vina-Rodriguez, A.; Sachse, K.; Ziegler, U.; Chaintoutis, S.C.; Keller, M.; Groschup, M.H.; Eiden, M. A novel pan-flavivirus detection and identification assay based on RT-qPCR and microarray. BioMed Res. Int. 2017, 2017, 4248756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools, P.; van Lieshout, L.; Koelewijn, R.; Addiss, D.; Ajjampur, S.S.R.; Ayana, M.; Bradbury, R.S.; Cantera, J.L.; Dana, D.; Fischer, K.; et al. First international external quality assessment scheme of nucleic acid amplification tests for the detection of Schistosoma and soil-transmitted helminths, including Strongyloides: A pilot study. PLoS Negl. Trop. Dis. 2020, 14, e0008231. [Google Scholar] [CrossRef] [PubMed]
- Che Lah, E.F.; Yaakop, S.; Ahamad, M.; Md Nor, S. Molecular identification of blood meal sources of ticks (Acari, Ixodidae) using cytochrome b gene as a genetic marker. Zookeys 2015, 478, 27–43. [Google Scholar] [CrossRef] [Green Version]
- Kent, R.J.; Norris, D.E. Identification of mammalian blood meals in mosquitoes by a multiplexed polymerase chain reaction targeting cytochrome B. Am. J. Trop. Med. Hyg. 2005, 73, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Khaklang, S.; Kittayapong, P. Species composition and blood meal analysis of mosquitoes collected from a tourist island, Koh Chang, Thailand. J. Vector Ecol. 2014, 39, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Roiz, D.; Vazquez, A.; Rosa, R.; Munoz, J.; Arnoldi, D.; Rosso, F.; Figuerola, J.; Tenorio, A.; Rizzoli, A. Blood meal analysis, flavivirus screening, and influence of meteorological variables on the dynamics of potential mosquito vectors of West Nile virus in northern Italy. J. Vector Ecol. 2012, 37, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Keven, J.B.; Artzberger, G.; Gillies, M.L.; Mbewe, R.B.; Walker, E.D. Probe-based multiplex qPCR identifies blood-meal hosts in Anopheles mosquitoes from Papua New Guinea. Parasites Vectors 2020, 13, 111. [Google Scholar] [CrossRef] [Green Version]
- Norris, L.C.; Fornadel, C.M.; Hung, W.C.; Pineda, F.J.; Norris, D.E. Frequency of multiple blood meals taken in a single gonotrophic cycle by Anopheles arabiensis mosquitoes in Macha, Zambia. Am. J. Trop. Med. Hyg. 2010, 83, 33–37. [Google Scholar] [CrossRef]
- Wheeler, S.S.; Taff, C.C.; Reisen, W.K.; Townsend, A.K. Mosquito blood-feeding patterns and nesting behavior of American crows, an amplifying host of West Nile virus. Parasites Vectors 2021, 14, 331. [Google Scholar] [CrossRef]
- Vazeille, M.; Zouache, K.; Vega-Rua, A.; Thiberge, J.M.; Caro, V.; Yebakima, A.; Mousson, L.; Piorkowski, G.; Dauga, C.; Vaney, M.C.; et al. Importance of mosquito “quasispecies” in selecting an epidemic arthropod-borne virus. Sci. Rep. 2016, 6, 29564. [Google Scholar] [CrossRef] [Green Version]
- Araya-Anchetta, A.; Busch, J.D.; Scoles, G.A.; Wagner, D.M. Thirty years of tick population genetics: A comprehensive review. Infect. Genet. Evol. 2015, 29, 164–179. [Google Scholar] [CrossRef] [Green Version]
- Seki, R.; Li, C.; Fang, Q.; Hayashi, S.; Egawa, S.; Hu, J.; Xu, L.; Pan, H.; Kondo, M.; Sato, T.; et al. Functional roles of Aves class-specific cis-regulatory elements on macroevolution of bird-specific features. Nat. Commun. 2017, 8, 14229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, B.F.; Goessling, L.S.; Storch, G.A.; Thach, R.E. Blood meal analysis to identify reservoir hosts for Amblyomma americanum ticks. Emerg. Infect. Dis. 2010, 16, 433–440. [Google Scholar] [CrossRef]
- Khristunova, Y.; Korotkova, E.; Kratochvil, B.; Barek, J.; Dorozhko, E.; Vyskocil, V.; Plotnikov, E.; Voronova, O.; Sidelnikov, V. Preparation and investigation of silver nanoparticle(-)antibody bioconjugates for electrochemical immunoassay of tick-borne encephalitis. Sensors 2019, 19, 2103. [Google Scholar] [CrossRef] [Green Version]
- Carter, J.R.; Balaraman, V.; Kucharski, C.A.; Fraser, T.S.; Fraser, M.J., Jr. A novel dengue virus detection method that couples DNAzyme and gold nanoparticle approaches. Virol. J. 2013, 10, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, S.M. The many faces of the flavivirus NS5 protein in antagonism of type I interferon signaling. J. Virol. 2017, 91, e01970-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.H.; Hong, S.K.; Yoon, S.H.; Jang, S.J.; Bahk, Y.Y.; Song, M.D.; Park, P.J.; Lee, K.H.; Kim, C.G.; Kim, B.; et al. Immunogenicity of Japanese encephalitis virus envelope protein by Hyphantria cunea nuclear polyhedrosis virus vector in guinea pig. Appl. Biochem. Biotechnol. 2012, 167, 259–269. [Google Scholar] [CrossRef]
- Garcia-Sastre, A.; Egorov, A.; Matassov, D.; Brandt, S.; Levy, D.E.; Durbin, J.E.; Palese, P.; Muster, T. Influenza A virus lacking the NS1 gene replicates in interferon-deficient systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Talon, J.; Horvath, C.M.; Polley, R.; Basler, C.F.; Muster, T.; Palese, P.; Garcia-Sastre, A. Activation of interferon regulatory factor 3 is inhibited by the influenza A virus NS1 protein. J. Virol. 2000, 74, 7989–7996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munir, S.; Hillyer, P.; Le Nouen, C.; Buchholz, U.J.; Rabin, R.L.; Collins, P.L.; Bukreyev, A. Respiratory syncytial virus interferon antagonist NS1 protein suppresses and skews the human T lymphocyte response. PLoS Pathog. 2011, 7, e1001336. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.S.; Brazas, R.M.; Holtzman, M.J. Respiratory syncytial virus nonstructural proteins NS1 and NS2 mediate inhibition of Stat2 expression and alpha/beta interferon responsiveness. J. Virol. 2005, 79, 9315–9319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, H.; Luo, H.; Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Medeiros, D.B.A.; Zou, J.; Xie, X.; Giraldo, M.I.; Vasconcelos, P.F.C.; et al. An evolutionary NS1 mutation enhances Zika virus evasion of host interferon induction. Nat. Commun. 2018, 9, 414. [Google Scholar] [CrossRef] [PubMed]
- Fajardo, T.; Sanford, T.J.; Mears, H.V.; Jasper, A.; Storrie, S.; Mansur, D.S.; Sweeney, T.R. The flavivirus polymerase NS5 regulates translation of viral genomic RNA. Nucleic Acids Res. 2020, 48, 5081–5093. [Google Scholar] [CrossRef] [PubMed]
- Fleith, R.C.; Mears, H.V.; Leong, X.Y.; Sanford, T.J.; Emmott, E.; Graham, S.C.; Mansur, D.S.; Sweeney, T.R. IFIT3 and IFIT2/3 promote IFIT1-mediated translation inhibition by enhancing binding to non-self RNA. Nucleic Acids Res. 2018, 46, 5269–5285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent-Rolle, M.; Morrison, J.; Rajsbaum, R.; Macleod, J.M.L.; Pisanelli, G.; Pham, A.; Ayllon, J.; Miorin, L.; Martinez, C.; tenOever, B.R.; et al. The interferon signaling antagonist function of yellow fever virus NS5 protein is activated by type I interferon. Cell Host Microbe 2014, 16, 314–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, F.M.; Greisinger, A.J.; Wehmanen, O.A.; Mouzoon, M.E.; Hoyle, J.C.; Smith, F.A.; Glezen, W.P. Safety of influenza vaccination during pregnancy. Am. J. Obstet. Gynecol. 2005, 192, 1098–1106. [Google Scholar] [CrossRef]
- Cumberworth, S.L.; Clark, J.J.; Kohl, A.; Donald, C.L. Inhibition of type I interferon induction and signalling by mosquito-borne flaviviruses. Cell. Microbiol. 2017, 19, e12737. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Chen, H.B.; Wang, X.J.; Huang, H.; Khromykh, A.A. Analysis of adaptive mutations in Kunjin virus replicon RNA reveals a novel role for the flavivirus nonstructural protein NS2A in inhibition of beta interferon promoter-driven transcription. J. Virol. 2004, 78, 12225–12235. [Google Scholar] [CrossRef] [Green Version]
- Anglero-Rodriguez, Y.I.; Pantoja, P.; Sariol, C.A. Dengue virus subverts the interferon induction pathway via NS2B/3 protease-IkappaB kinase epsilon interaction. Clin. Vaccine Immunol. 2014, 21, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Dalrymple, N.A.; Cimica, V.; Mackow, E.R. Dengue virus NS proteins inhibit RIG-I/MAVS signaling by blocking TBK1/IRF3 phosphorylation: Dengue virus serotype 1 NS4A is a unique interferon-regulating virulence determinant. mBio 2015, 6, e00553-15. [Google Scholar] [CrossRef] [Green Version]
- Su, H.L.; Lin, Y.L.; Yu, H.P.; Tsao, C.H.; Chen, L.K.; Liu, Y.T.; Liao, C.L. The effect of human bcl-2 and bcl-X genes on dengue virus-induced apoptosis in cultured cells. Virology 2001, 282, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.R.; Taylor, S.; Fraser, T.S.; Kucharski, C.A.; Dawson, J.L.; Fraser, M.J., Jr. Suppression of the arboviruses dengue and chikungunya using a dual-acting group-I intron coupled with conditional expression of the bax C-terminal domain. PLoS ONE 2015, 10, e0139899. [Google Scholar] [CrossRef]
- Carter, J.R.; Keith, J.H.; Fraser, T.S.; Dawson, J.L.; Kucharski, C.A.; Horne, K.M.; Higgs, S.; Fraser, M.J., Jr. Effective suppression of dengue virus using a novel group-I intron that induces apoptotic cell death upon infection through conditional expression of the Bax C-terminal domain. Virol. J. 2014, 11, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, K.S.M.; Her, Z.; Chen, Q. Humanized mice as unique tools for human-specific studies. Arch. Immunol. Ther. Exp. 2018, 66, 245–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta, P.L.; Byrne, A.B.; Hijano, D.R.; Talarico, L.B. Human type I interferon antiviral effects in respiratory and reemerging viral infections. J. Immunol. Res. 2020, 2020, 1372494. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, J. Review of diagnostic plaque reduction neutralization tests for flavivirus infection. Vet. J. 2013, 195, 33–40. [Google Scholar] [CrossRef]
- Khine, P.K.; Schneider, H. Data on pteridophyte species diversity and status of the International Union for Conservation of Nature in each political unit of Myanmar. Data Brief 2020, 30, 105503. [Google Scholar] [CrossRef]
- Sollmann, R.; Eaton, M.J.; Link, W.A.; Mulondo, P.; Ayebare, S.; Prinsloo, S.; Plumptre, A.J.; Johnson, D.S. A Bayesian Dirichlet process community occupancy model to estimate community structure and species similarity. Ecol. Appl. 2021, 31, e02249. [Google Scholar] [CrossRef]
- Harfoot, M.B.J.; Johnston, A.; Balmford, A.; Burgess, N.D.; Butchart, S.H.M.; Dias, M.P.; Hazin, C.; Hilton-Taylor, C.; Hoffmann, M.; Isaac, N.J.B.; et al. Using the IUCN Red List to map threats to terrestrial vertebrates at global scale. Nat. Ecol. Evol. 2021. [Google Scholar] [CrossRef]
- Hardy, J.M.; Newton, N.D.; Modhiran, N.; Scott, C.A.P.; Venugopal, H.; Vet, L.J.; Young, P.R.; Hall, R.A.; Hobson-Peters, J.; Coulibaly, F.; et al. A unified route for flavivirus structures uncovers essential pocket factors conserved across pathogenic viruses. Nat. Commun. 2021, 12, 3266. [Google Scholar] [CrossRef] [PubMed]
- Starolis, M.W.; Perez, O.; Powell, E.A. Clinical features and laboratory diagnosis of emerging arthropod-transmitted viruses: A Report from the Pan American Society for Clinical Virology Clinical Practice Committee. J. Clin. Virol. 2020, 132, 104651. [Google Scholar] [CrossRef] [PubMed]
- Daep, C.A.; Munoz-Jordan, J.L.; Eugenin, E.A. Flaviviruses, an expanding threat in public health: Focus on dengue, West Nile, and Japanese encephalitis virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
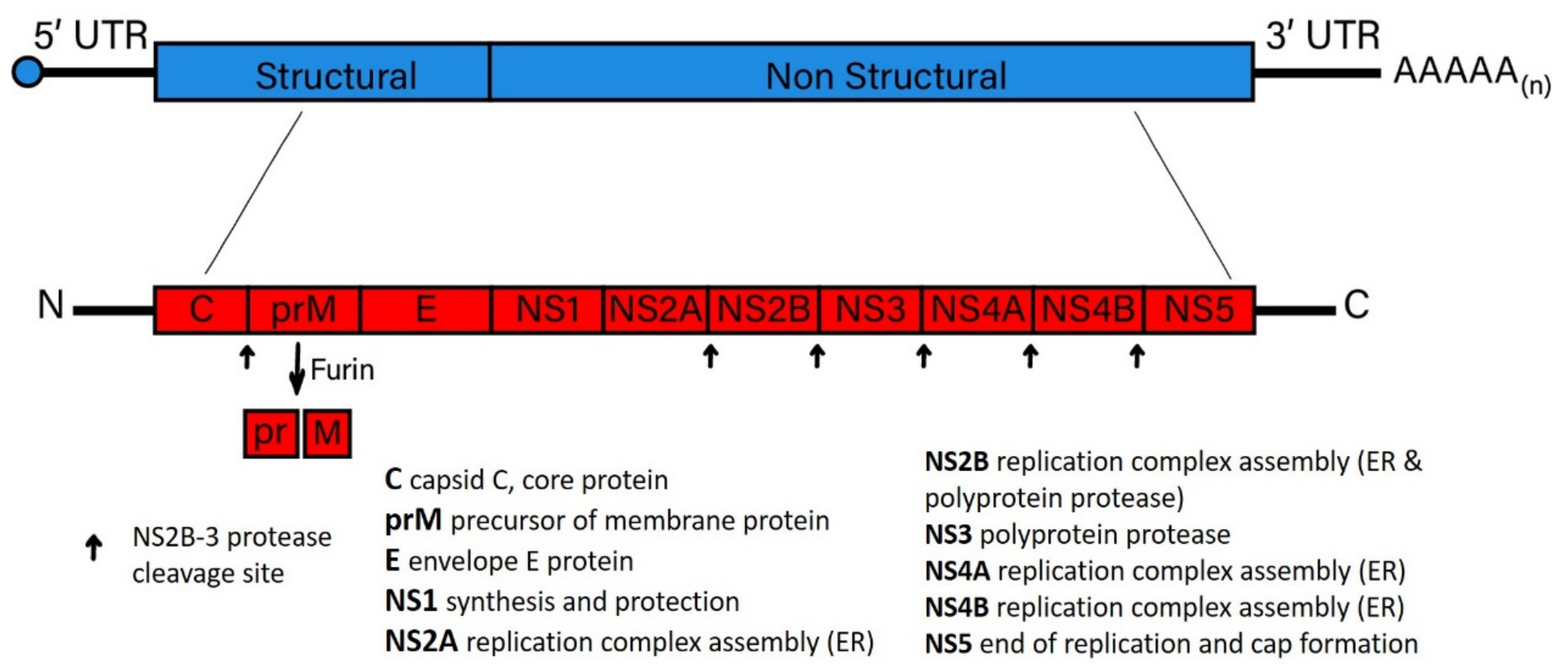
| Flavivirus Non-Structural Proteins | Mosquito-Borne Flavivirus | Tick-Borne Flavivirus ** |
|---|---|---|
| NS1 | NS1-mediated immune evasion leads to the activation of human complement by the specific inhibition of the classical and lectin pathways of complement activation through a direct interaction with complement components C4 and C1s [88]. For example, DENV, WNV, and YFV NS1 proteins have been shown to confine C4b expression and C3 convertase activity by increasing the digestion of C4 through the recruitment of the complement-specific protease C1s [88,89,90]. This effect might explain some of the clinical manifestations of dengue hemorrhagic fever and dengue shock syndrome. Each subunit forms a homodimer located in the ER lumen and co-localizes with viral dsRNA. Secreted and cell surface-associated NS1 is immunogenic and induces an antibody response that serves as a marker of flavivirus infection [91]. | Degradation of TBEV-NS1 proteins also restricts TBEV replication [92]. Tick-borne encephalitis virus Far Eastern subtype (Sofjin virus) is associated with a 39-kDa truncated form of NS1 with acutely and persistently infected cells [93]. |
| NS2A | Mutations within a pseudo-knot of NS2A RNA that is characteristic of the JEV subgroup abolish NS2A interaction with NS1, suggesting a role in viral neuroinvasiveness and attenuation in mice [94], and also suggests a potential link between neuroinvasiveness and flavivirus persistence in wildlife species. | |
| NS2B | NS2B is a hydrophobic protein that behaves as a cofactor for NS3. Together, they form a serine protease complex essential for processing the flavivirus polyprotein [95]. | |
| NS3 | NS3 protein is a critical member of the Replication Complex (RC) and is activated with NS5 to bind the genomic RNA prior to replication [96,97]. Mosquito-borne flavivirus NS3 can also potentially induce [98,99,100,101] INFα/β signaling [102]. For example, studies on neurovirulence associated with DENV-1 NS3 have shown that mutations in NS3 induce cell death in DENV-1-infected cells [103]. | The protease domain of LGTV NS3 associates with caspase 8 and induces apoptosis [104]. |
| NS4A | NS4A, in concert with NS3 and NS4B flavivirus proteins, is responsible for promoting the reorganization of host ER membranes, resulting in the development of virus-induced membranous spherules and vesicles enclosing the dsRNA and RC, potentially diminishing the exposure of actively replicating flavivirus RNA to innate immune sensor proteins, such as melanoma differentiation-associated gene 5 (MDA5) and retinoic acid-inducible gene I (RIG-I) [96,105,106], potentially aiding in the establishment of flavivirus persistence. The mature form of NS4A also induces PI3K-dependent autophagy signaling, leading to protection from antiviral-induced cell death [107]. | |
| NS4B | The NS4B protein of DENV, JEV, and WNV, inhibits type I interferon (IFN-α/β) response through the inhibition of STAT1 phosphorylation [108,109,110]. The same is true for the WNV strain, NY99, which disrupts the host immune responses by blocking IFN-α/β/λ pathways through the disruption of the phosphoactivation of STAT1/STAT2, which is essential for nuclear translocation [108]. | |
| NS5 | The largest and most conserved among the vector-borne flavivirus proteins. NS5 primarily functions as the RNA-dependent RNA polymerase (RdRp) [97]. For WNV, NS5 is involved in different cellular pathways and has a crucial role in the escape from the IFN-α/β signaling pathway, typically through inhibiting phosphorylation of protein members of this pathway. DENV NS5-mediated inhibition of host TYK2 and STAT2 phosphorylation thwarts JAK-STAT signaling pathway activation [111]. | Supplemental to functions described for the viral RdRp, NS5 described for the mosquito-borne viruses, and the NS5 of tick-borne flaviviruses were the first to be shown to disrupt innate immune signaling. For example, the suppression of critical host responses is shown through LGTV NS5 interactions with IFNAR2 and IFNGR2 (IFN receptor subunits) and antagonizes IFN-dependent responses via JAK-STAT signal transduction suppression [112,113]. The TBEV NS5 interacts with TRIM79-α, an IFN-inducible protein, leading to the inhibition of TRIM79-mediated degradation of TBEV-encoded proteins [86]. The ability of host cells to suppress tick-borne flaviviruses is compromised by the interaction of TBEV NS5 with a cellular scaffold protein, named Scribble, which blocks STAT1 phosphorylation and disrupts JAK-STAT-mediated signaling [114]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blahove, M.R.; Carter, J.R. Flavivirus Persistence in Wildlife Populations. Viruses 2021, 13, 2099. https://doi.org/10.3390/v13102099
Blahove MR, Carter JR. Flavivirus Persistence in Wildlife Populations. Viruses. 2021; 13(10):2099. https://doi.org/10.3390/v13102099
Chicago/Turabian StyleBlahove, Maria Raisa, and James Richard Carter. 2021. "Flavivirus Persistence in Wildlife Populations" Viruses 13, no. 10: 2099. https://doi.org/10.3390/v13102099
APA StyleBlahove, M. R., & Carter, J. R. (2021). Flavivirus Persistence in Wildlife Populations. Viruses, 13(10), 2099. https://doi.org/10.3390/v13102099