
High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia

 ,
,  and
and 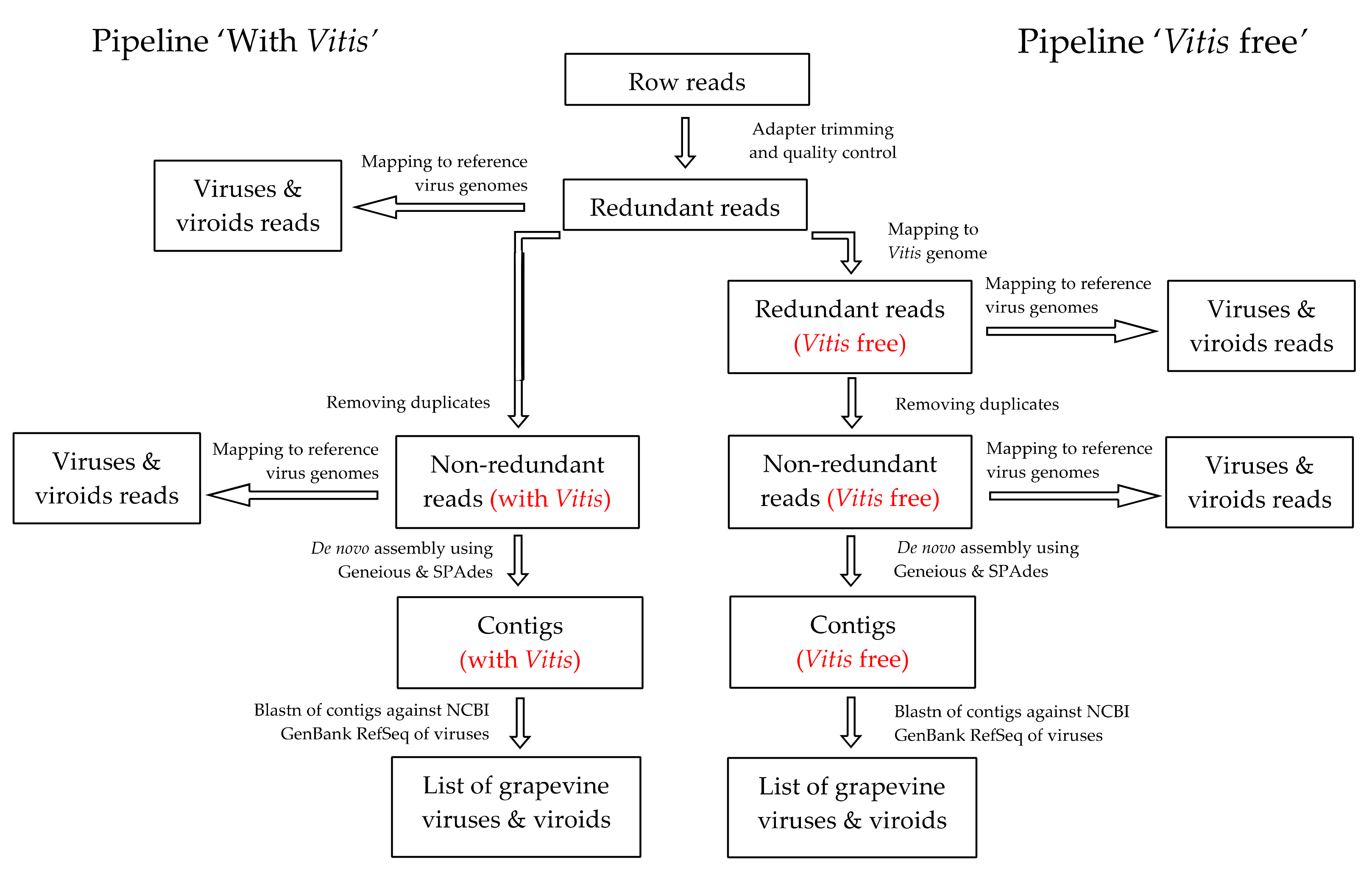{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sources and Preparation of sRNA Sequencing Libraries
- Phytosanitary monitoring of commercial vineyards in Krasnodar Krai was carried out from July 2017 to September 2018 The exact coordinates of the vineyard are available in Google Earth (https://earth.google.com/earth/d/1KymmhGwAdv4Wns6VikJ2W_R736-UeR-U?usp=sharing) (accessed on 30 November 2021). For the analysis, samples were collected from grapevine plants with symptoms of viral disease (Supplementary Table S1, Supplementary Photographic Materials). To isolate total RNA, we used 1 g of vine and leaves from each of the 38 plants in accordance with the protocol described by Morante-Carriel [24]. RNA weighing 2–5 μg was extracted from 7–9 plants and combined into one pool, and sRNA sequencing libraries were prepared using the TruSeq Small RNA Library Preparation Kit (Illumina, San Diego, CA, USA) in accordance with the manufacturer’s protocol with modifications following Czotter et al. (2018) [21]. The resulting five libraries were sequenced using a HiScanSQ System (Illumina, USA) at a read length of 1 × 50 bp. Fastq files of the sequenced libraries were deposed in the SRA and can be accessed at accession number PRJNA771403.
2.2. Bioinformatics Analysis of HTS Results
2.3. Data Validation by RT-PCR
2.4. Phylogenetic Analysis
2.5. Recombination Analysis
3. Results and Discussion
3.1. Phytosanitary Monitoring
3.2. Results of the sRNA HTS Bioinformatics Analysis
3.3. Validation of the Results of sRNA HTS by RT-PCR and the Phylogenetic Analysis of Detected Viruses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meng, B.; Martelli, G.P.; Golino, D.A.; Fuchs, M. (Eds.) The Grapevine, Viticulture, and Winemaking: A Brief Introduction. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Mannini, F.; Digiaro, M. The Effects of Viruses and Viral Diseases on Grapes and Wine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 453–482. [Google Scholar] [CrossRef]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids, and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 31–46. [Google Scholar] [CrossRef]
- Martelli, G. Where Grapevine Virology Is Heading to. In Proceedings of the 19th Congress of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine (ICVG), Santiago, Chile, 9–12 April 2018; pp. 10–15. [Google Scholar]
- Fuchs, M. Grapevine Viruses: A Multitude of Diverse Species with Simple but Overall Poorly Adopted Management Solutions in the Vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Grapevine Fanleaf Virus and Other Old World Nepoviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 47–82. [Google Scholar] [CrossRef]
- Naidu, R.A. Grapevine Leafroll-Associated Virus 1. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 127–139. [Google Scholar] [CrossRef] [Green Version]
- Angelini, E.; Aboughanem-Sabanadzovic, N.; Dolja, V.V.; Meng, B. Grapevine Leafroll-Associated Virus 2. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 141–165. [Google Scholar] [CrossRef]
- Burger, J.T.; Maree, H.J.; Gouveia, P.; Naidu, R.A. Grapevine Leafroll-Associated Virus 3. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 167–195. [Google Scholar] [CrossRef] [Green Version]
- Minafra, A.; Mawassi, M.; Goszczynski, D.; Saldarelli, P. Grapevine Vitiviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 229–256. [Google Scholar] [CrossRef]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in Virus Diagnostics: From ELISA to next Generation Sequencing. Virus Res. 2014, 186, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current Impact and Future Directions of High Throughput Sequencing in Plant Virus Diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of Next-Generation Sequencing Versus Biological Indexing for the Optimal Detection of Viral Pathogens in Grapevine. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Roossinck, M.J. Deep Sequencing for Discovery and Evolutionary Analysis of Plant Viruses. Virus Res. 2017, 239, 82–86. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant Virus Metagenomics: Advances in Virus Discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [Green Version]
- Giampetruzzi, A.; Roumi, V.; Roberto, R.; Malossini, U.; Yoshikawa, N.; La Notte, P.; Terlizzi, F.; Credi, R.; Saldarelli, P. A New Grapevine Virus Discovered by Deep Sequencing of Virus- and Viroid-Derived Small RNAs in Cv Pinot Gris. Virus Res. 2012, 163, 262–268. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Candresse, T.; Katis, N.I.; Avgelis, A.D. A Novel Grapevine Badnavirus Is Associated with the Roditis Leaf Discoloration Disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Singh, K.; Kaur, R.; Qiu, W. Association of a Novel DNA Virus with the Grapevine Vein-Clearing and Vine Decline Syndrome. Phytopathology 2011, 101, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Blouin, A.G.; Keenan, S.; Napier, K.R.; Barrero, R.A.; MacDiarmid, R.M. Identification of a Novel Vitivirus from Grapevines in New Zealand. Arch. Virol. 2018, 163, 281–284. [Google Scholar] [CrossRef]
- Blouin, A.G.; Chooi, K.M.; Warren, B.; Napier, K.R.; Barrero, R.A.; MacDiarmid, R.M. Grapevine Virus I, a Putative New Vitivirus Detected in Co-Infection with Grapevine Virus G in New Zealand. Arch. Virol. 2018, 163, 1371–1374. [Google Scholar] [CrossRef]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deak, T.; Bisztray, G.; et al. NGS of Virus-Derived Small RNAs as a Diagnostic Method Used to Determine Viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Demian, E.; Jaksa-Czotter, N.; Molnar, J.; Tusnady, G.E.; Kocsis, L.; Varallyay, E. Grapevine Rootstocks Can Be a Source of Infection with Non-Regulated Viruses. Eur. J. Plant Pathol. 2020, 156, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Pooggin, M.M. Small RNA-Omics for Plant Virus Identification, Virome Reconstruction, and Antiviral Defense Characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Morante-Carriel, J.; Sellés-Marchart, S.; Martínez-Márquez, A.; Martínez-Esteso, M.J.; Luque, I.; Bru-Martínez, R. RNA Isolation from Loquat and Other Recalcitrant Woody Plants with High Quality and Yield. Anal. Biochem. 2014, 452, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Geneious Bioinformatics Software for Sequence Data Analysis. Available online: https://www.geneious.com/ (accessed on 30 April 2021).
- FinchTV; Version 1.4.0; Geospiza, Inc.: Seattle, WA, USA, 2004–2012.
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2020, 48, D84–D86. [Google Scholar] [CrossRef] [Green Version]
- Bertazzon, N.; Angelini, E. Advances in the Detection of Grapevine Leafroll-Associated Virus 2 Variants. J. Plant Pathol. 2004, 86, 283–290. [Google Scholar]
- Porotikova, E.; Terehova, U.; Volodin, V.; Yurchenko, E.; Vinogradova, S. Distribution and Genetic Diversity of Grapevine Viruses in Russia. Plants 2021, 10, 1080. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substituions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Mekuria, T.A.; Gutha, L.R.; Martin, R.R.; Naidu, R.A. Genome Diversity and Intra- and Interspecies Recombination Events in Grapevine Fanleaf Virus. Phytopathology 2009, 99, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Alkowni, R.; Zhang, Y.-P.; Rowhani, A.; Uyemoto, J.K.; Minafra, A. Biological, Molecular, and Serological Studies of a Novel Strain of Grapevine Leafroll-Associated Virus 2. Virus Genes 2011, 43, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Ghebremeskel, S.; Martelli, G.P. Grapevine Deformation Virus: Completion of the Sequence and Evidence on Its Origin from Recombination Events between Grapevine Fanleaf Virus and Arabis Mosaic Virus. Virus Res. 2012, 166, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Vigne, E.; Marmonier, A.; Fuchs, M. Multiple Interspecies Recombination Events within RNA2 of Grapevine Fanleaf Virus and Arabis Mosaic Virus. Arch. Virol. 2008, 153, 1771–1776. [Google Scholar] [CrossRef]
- Cretazzo, E.; Padilla, C.V.; Velasco, L. First Report of Grapevine Red Globe Virus in Grapevine in Spain. Available online: https://www.cabi.org/ISC/abstract/20173050622 (accessed on 28 April 2021).
- Sabanadzovic, S.; Abou-Ghanem, N.; Castellano, M.A.; Digiaro, M.; Martelli, G.P. Grapevine Fleck Virus-like Viruses in Vitis. Arch. Virol. 2000, 145, 553–565. [Google Scholar] [CrossRef]
- Chiumenti, M.; Giampetruzzi, A.; Pirolo, C.; Morelli, M.; Saldarelli, P.; Minafra, A.; Bottalico, G.; La Notte, P.; Campanale, A.; Savino, V. Approaches of next Generation Sequencing to Investigate Grapevine Diseases of Unknown Etiology. In Proceedings of the 17th Congress of ICVG, Davis, CA, USA, 7–14 October 2012; pp. 116–117. [Google Scholar]
- Eichmeier, A.; Komínková, M.; Komínek, P.; Baránek, M. Comprehensive Virus Detection Using Next Generation Sequencing in Grapevine Vascular Tissues of Plants Obtained from the Wine Regions of Bohemia and Moravia (Czech Republic). PLoS ONE 2016, 11, e0167966. [Google Scholar] [CrossRef]
- Sidharthan, V.K.; Sevanthi, A.M.; Jaiswal, S.; Baranwal, V.K. Robust Virome Profiling and Whole Genome Reconstruction of Viruses and Viroids Enabled by Use of Available MRNA and SRNA-Seq Datasets in Grapevine (Vitis vinifera L.). Front. Microbiol. 2020, 11, 1232. [Google Scholar] [CrossRef]
- Demian, E.; Holczbauer, A.; Galbacs, Z.N.; Jaksa-Czotter, N.; Turcsan, M.; Olah, R.; Varallyay, E. Variable Populations of Grapevine Virus T Are Present in Vineyards of Hungary. Viruses 2021, 13, 1119. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Rowhani, A. Deep Sequencing Analysis of RNAs from a Grapevine Showing Syrah Decline Symptoms Reveals a Multiple Virus Infection That Includes a Novel Virus. Virology 2009, 387, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Komar, V.; Vigne, E.; Demangeat, G.; Fuchs, M. Beneficial Effect of Selective Virus Elimination on the Performance of Vitis vinifera Cv. Chardonnay. Am. J. Enol. Vitic. 2007, 58, 202–210. [Google Scholar]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine Leafroll-Associated Virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Porotikova, E.V.; Risovannaya, V.I.; Volkov, Y.A.; Dmitrenko, U.D.; Volodin, V.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Agranovsky, A.A.; Kamionskaya, A.M.; Vinogradova, S.V. Occurrence of Grapevine Leafroll-Associated Viruses-1 and -3 in Crimea. Moscow Univ. Biol. Sci. Bull. 2016, 71, 76–79. [Google Scholar] [CrossRef] [Green Version]
- Porotikova, E.V.; Dmitrenko, U.D.; Yurchenko, E.G.; Vinogradova, S.V. First Report of Grapevine Leafroll-Associated Virus 2 in Russian Grapevines (Vitis vinifera). Plant Dis. 2019, 103, 164. [Google Scholar] [CrossRef]
- Vinogradova, S.V.; Navrotskaya, E.V.; Porotikova, E.V.; Massart, S.; Varallyay, E. The First Virome of Russian Vineyard. Plants 2021, in press. [Google Scholar]
- Lehad, A.; Selmi, I.; Louanchi, M.; Aitoua, M.; Mahfoudhi, N. Occurrence and Diversity of Grapevine Leafroll—Associated Virus 1 in Algeria. Phytopathol. Mediterr. 2019, 58, 277–281. [Google Scholar]
- Fonseca, F.; Esteves, F.; Teixeira Santos, M.; Brazao, J.; Eiras-Dias, J.E. Genetic Variants of Grapevine Leafroll-Associated Virus 2 Infecting Portuguese Grapevine Cultivars. Phytopathol. Mediterr. 2016, 55, 73–88. [Google Scholar]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y.; et al. Characterization of Grapevine Leafroll-Associated Virus 3 Genetic Variants and Application towards RT-QPCR Assay Design. PLoS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [Green Version]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus Taxonomy: The Database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [Green Version]
- Adiputra, J.; Jarugula, S.; Naidu, R.A. Intra-Species Recombination among Strains of the Ampelovirus Grapevine Leafroll-Associated Virus 4. Virol. J. 2019, 16, 139. [Google Scholar] [CrossRef]
- Andret-Link, P.; Laporte, C.; Valat, L.; Ritzenthaler, C.; Demangeat, G.; Vigne, E.; Laval, V.; Pfeiffer, P.; Stussi-Garaud, C.; Fuchs, M. Grapevine fanleaf virus: Still a major threat to the grapevine industry. J. Plant Pathol. 2004, 86, 183–195. [Google Scholar]
- Volodin, V.; Gorislavets, S.; Risovannaya, V.; Stranishevskaya, E.; Shadura, N.; Volkov, Y.; Matveykina, E. Detection of a Viral Infection Complex (GLRAV-1, -3 and GFLV) in Vineyards of Crimea. Vitic. Winemak. 2020, 49, 124–126. [Google Scholar]
- Ebrahimi Ghomi, M.; Shamsbakhsh, M.; Pourrahim, R. Study on the status of three grapevine viruses in North-Eastern vineyards of Iran. Appl. Entomol. Phytopathol. 2007, 75, 109–121. [Google Scholar]
- Rakhshandehroo, F.; Pourrahim, R.; Zadeh, H.Z.; Rezaee, S.; Mohammadi, M. Incidence and Distribution of Viruses Infecting Iranian Vineyards. J. Phytopathol. 2005, 153, 480–484. [Google Scholar] [CrossRef]
- Porotikova, E.V.; Dmitrenko, U.D.; Volodin, V.A.; Volkov, Y.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Risovannaya, V.I.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine Virus A in Russian Grapevines. Plant Dis. 2016, 100, 2541. [Google Scholar] [CrossRef]
- Alabi, O.J.; Rwahnih, M.A.; Mekuria, T.A.; Naidu, R.A. Genetic Diversity of Grapevine Virus A in Washington and California Vineyards. Phytopathology 2014, 104, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G. (Ed.) Rugose Wood Complex. Graft-Transmissible Disease of Grapevines, Handbook for Detection and Diagnosis; Food and Agriculture Organization of The United Nations: Rome, Italy, 1993; pp. 45–53. [Google Scholar]
- Zhang, Y.-P.; Uyemoto, J.K.; Golino, D.A.; Rowhani, A. Nucleotide Sequence and RT-PCR Detection of a Virus Associated with Grapevine Rupestris Stem-Pitting Disease. Phytopathology 1998, 88, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Dmitrenko, U.D.; Porotikova, E.V.; Gorislavets, S.M.; Risovannaya, V.I.; Volkov, Y.A.; Stranishevskaya, E.P.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine Rupestris Stem Pitting-Associated Virus in Russia. Plant Dis. 2016, 100, 2542. [Google Scholar] [CrossRef]
- Komorowska, B.; Berniak, H.; Golis, T. Detection of Grapevine Viruses in Poland. J. Phytopathol. 2014, 162, 326–331. [Google Scholar] [CrossRef]
- Pacifico, D.; Stigliano, E.; Sposito, L.; Spinelli, P.; Garfì, G.; Gristina, A.S.; Fontana, I.; Carimi, F. Survey of Viral Infections in Spontaneous Grapevines from Natural Environments in Sicily. Eur. J. Plant Pathol. 2016, 145, 189–197. [Google Scholar] [CrossRef]
- Katarina, H.; Saldarelli, P.; Čarija, M.; Černi, S.; Goran, Z.; Mucalo, A.; Tomislav, R. Predominance and Diversity of GLRaV-3 in Native Vines of Mediterranean Croatia. Plants 2020, 10, 17. [Google Scholar] [CrossRef]
- Hily, J.-M.; Candresse, T.; Garcia, S.; Vigne, E.; Tannière, M.; Komar, V.; Barnabé, G.; Alliaume, A.; Gilg, S.; Hommay, G.; et al. High-Throughput Sequencing and the Viromic Study of Grapevine Leaves: From the Detection of Grapevine-Infecting Viruses to the Description of a New Environmental Tymovirales Member. Front. Microbiol. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Selmi, I.; Pacifico, D.; Lehad, A.; Stigliano, E.; Crucitti, D.; Carimi, F.; Naima, M. Genetic Diversity of Grapevine Rupestris Stem Pitting-Associated Virus Isolates from Tunisian Grapevine Germplasm—Selmi—2020—Plant Pathology—Wiley Online Library. Available online: https://bsppjournals.onlinelibrary.wiley.com/doi/full/10.1111/ppa.13183?casa_token=QfyTerWj6LwAAAAA%3AK3ug7fUFAs91pMxBeulOZ4YFpR7LKLdCsPYdlycRc0fK8AvOk8lFHyRIx3ouoH7zhJCG3_99W2j9ci8vMaw (accessed on 15 July 2021).
- Glasa, M.; Predajna, L.; Soltys, K.; Sihelska, N.; Nagyova, A.; Wetzel, T.; Sabanadzovic, S. Analysis of Grapevine Rupestris Stem Pitting-Associated Virus in Slovakia Reveals Differences in Intra-Host Population Diversity and Naturally Occurring Recombination Events. Plant Pathol. J. 2017, 33, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Beuve, M.; Candresse, T.; Tannières, M.; Lemaire, O. First Report of Grapevine Pinot Gris Virus (GPGV) in Grapevine in France. Plant Dis. 2015, 99, 293. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Golino, D.; Rowhani, A. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in the United States. Plant Dis. 2015, 100, 1030. [Google Scholar] [CrossRef]
- Cho, I.S.; Jung, S.M.; Cho, J.D.; Choi, G.S.; Lim, H.S. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Korea. N. Dis. Rep. 2013, 27, 10. [Google Scholar] [CrossRef] [Green Version]
- Gazel, M.; Caglayan, K.; Elçi, E.; Öztürk, L. First Report of Grapevine Pinot Gris Virus in Grapevine in Turkey. Plant Dis. 2015, 100, 657. [Google Scholar] [CrossRef]
- Fan, X.D.; Dong, Y.F.; Zhang, Z.P.; Ren, F.; Hu, G.J.; Li, Z.N.; Zhou, J. First Report of Grapevine Pinot Gris Virus in Grapevines in China. Plant Dis. 2015, 100, 540. [Google Scholar] [CrossRef]
- Xiao, H.; Shabanian, M.; McFadden-Smith, W.; Meng, B. First Report of Grapevine Pinot Gris Virus in Commercial Grapes in Canada. Plant Dis. 2015, 100, 1030. [Google Scholar] [CrossRef]
- Reynard, J.-S.; Schumacher, S.; Menzel, W.; Fuchs, J.; Bohnert, P.; Glasa, M.; Wetzel, T.; Fuchs, R. First Report of Grapevine Pinot Gris Virus in German Vineyards. Plant Dis. 2016, 100, 2545. [Google Scholar] [CrossRef]
- Rasool, S.; Naz, S.; Rowhani, A.; Golino, D.A.; Westrick, N.M.; Farrar, K.D.; Al Rwahnih, M. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Pakistan. Plant Dis. 2017, 101, 1958. [Google Scholar] [CrossRef]
- Ruiz-García, A.B.; Olmos, A. First Report of Grapevine Pinot Gris Virus in Grapevine in Spain. Plant Dis. 2017, 101, 1070. [Google Scholar] [CrossRef]
- Fajardo, T.V.M.; Eiras, M.; Nickel, O. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Brazil. Australas. Plant Dis. Notes 2017, 12, 45. [Google Scholar] [CrossRef]
- Zamorano, A.; Medina, G.; Fernández, C.; Cui, W.; Quiroga, N.; Fiore, N. First Report of Grapevine Pinot Gris Virus in Grapevine in Chile. Plant Dis. 2019, 103, 1438. [Google Scholar] [CrossRef]
- Abou Kubaa, R.; Choueiri, E.; Jreijiri, F.; El Khoury, Y.; Saldarelli, P. First Report of Grapevine Pinot Gris Virus in Lebanon and the Middle East. J. Plant Pathol. 2020, 102, 565. [Google Scholar] [CrossRef] [Green Version]
- Abou Kubaa, R.; Lanotte, P.; Saldarelli, P. First Report of Grapevine Pinot Gris Virus in Grapevine in Moldavia. J. Plant Pathol. 2019, 101, 441. [Google Scholar] [CrossRef] [Green Version]
- Casati, P.; Maghradze, D.; Quaglino, F.; Ravasio, A.; Failla, O.; Bianco, P.A. First Report of Grapevine Pinot Gris Virus in Georgia. J. Plant Pathol. 2015, 97, 67. [Google Scholar]
- Tokhmechi, K.; Koolivand, D. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Iran. J. Plant Pathol. 2020, 102, 549. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Habili, N. The Recent Importation of Grapevine Pinot Gris Virus into Australia. Virus Genes 2017, 53, 935–938. [Google Scholar] [CrossRef]
- Eichmeier, A.; Peňázová, E.; Pavelková, R.; Mynarzová, Z.; Saldarelli, P. Detection of Grapevine Pinot Gris Virus in Certified Grapevine Stocks in Moravia, Czech Republic. J. Plant Pathol. 2016, 98, 155–157. [Google Scholar]
- Bertazzon, N.; Rahali, M.; Angelini, E.; Crespan, M.; Migliaro, D. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Algeria. Plant Dis. 2020, 105, 234. [Google Scholar] [CrossRef]
- Silva, G.; Lecourt, J.; Clover, G.R.G.; Seal, S.E. First Record of Grapevine Pinot Gris Virus Infecting Vitis vinifera in the United Kingdom. N. Dis. Rep. 2018, 38, 7. [Google Scholar] [CrossRef] [Green Version]
- Eichmeier, A.; Penazova, E.; Nebish, A. First Report of Grapevine Pinot Gris Virus on Grapevines in Armenia. Plant Dis. 2019, 104, 1000. [Google Scholar] [CrossRef]
- Bertazzon, N.; Angelini, E.; Signorotto, M.; Genov, N. First Report of Grapevine Pinot Gris Virus and Grapevine Leafroll-Associated Virus 2 in Bulgarian Vineyards. J. Plant Dis. Prot. 2021, 128, 597–599. [Google Scholar] [CrossRef]
- Debat, H.; Luna, F.; Moyano, S.; Zavallo, D.; Asurmendi, S.; Gomez-Talquenca, S. First Report of Grapevine Pinot Gris Virus Infecting Grapevine in Argentina. J. Plant Pathol. 2020, 102, 1321. [Google Scholar] [CrossRef]
- Rwahnih, M.A.; Diaz-Lara, A.; Arnold, K.; Cooper, M.L.; Smith, R.J.; Zhuang, G.; Battany, M.C.; Bettiga, L.J.; Rowhani, A.; Golino, D. Incidence and Genetic Diversity of Grapevine Pinot Gris Virus in California. Am. J. Enol. Vitic. 2021, 72, 164–169. [Google Scholar] [CrossRef]
- Abe, J.; Tomoyuki, N. First Report of Grapevine Pinot Gris Virus in Wild Grapevines (Vitis coignetiae) in Japan. J. Plant Pathol. 2021, 103, 725. [Google Scholar] [CrossRef]
- Massart, S.; Vankerkoven, L.; Blouin, A.G.; Nourinejhad Zarghani, S.; Wetzel, T. First Report of Grapevine Pinot Gris Virus and Grapevine Rupestris Stem Pitting-Associated Virus in Grapevine in Belgium. Plant Dis. 2020, 104, 1879. [Google Scholar] [CrossRef]
- Guta, I.-C.; Buciumeanu, E.-C. Grapevine Pinot Gris Virus Infecting Grapevines in Romania–Short Communicaiton. Hortic. Sci. 2021, 48, 47–50. [Google Scholar] [CrossRef]
- Tarquini, G.; Zaina, G.; Ermacola, P.; De Amicis, F.; Franco-Orozco, B.; Loi, N.; Martini, M.; Luca Bianchi, G.; Pagliari, L.; Firrao, G.; et al. Agroinoculation of Grapevine Pinot Gris Virus in Tobacco and Grapevine Provides Insights on Viral Pathogenesis. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0214010 (accessed on 28 September 2021).
- Sabanadzovic, S.; Aboughanem-Sabanadzovic, N.; Martelli, G.P. Grapevine Fleck and Similar Viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 331–349. [Google Scholar] [CrossRef]
- Crnogorac, A.; Gašpar, M.; Davino, S.; Mandić, A.; Matić, S. First Report of Grapevine Fleck Virus in Vineyards of Bosnia and Herzegovina. J. Plant Pathol. 2020, 102, 1299. [Google Scholar] [CrossRef]
- Kanuya, E.; Clayton, L.A.; Naidu, R.A.; Karasev, A.V. First Report of Grapevine Fleck Virus in Idaho Grapevines. Plant Dis. 2012, 96, 1705. [Google Scholar] [CrossRef]
- Poojari, S.; Lowery, T.; Rott, M.; Schmidt, A.-M.; DeLury, N.; Boulé, J.; Úrbez-Torres, J.R. First Report and Prevalence of Grapevine Fleck Virus in Grapevines (Vitis vinifera) in Canada. Plant Dis. 2016, 100, 1028. [Google Scholar] [CrossRef]
- Kostadinovska, E.; Mitrev, S.; Bianco, P.A.; Casati, P.; Bulgari, D. First Report of Grapevine Virus A and Grapevine Fleck Virus in the Former Yugoslav Republic of Macedonia. Plant Dis. 2014, 98, 1747. [Google Scholar] [CrossRef]
- Jo, Y.; Song, M.K.; Choi, H.; Park, J.S.; Lee, J.W.; Cho, W.K. First Report of Grapevine Fleck Virus and Grapevine Virus E in Grapevine in Korea. Plant Dis. 2017, 101, 1069. [Google Scholar] [CrossRef]
- Immanuel, T.M.; Delmiglio, C.; Ward, L.I.; Denton, J.O.; Clover, G.R.G. First Reports of Grapevine Virus A, Grapevine Fleck Virus, and Grapevine Leafroll-Associated Virus 1 in the United Kingdom. Plant Dis. 2015, 99, 898. [Google Scholar] [CrossRef]
- Ma, Y.X.; Li, S.F.; Zhang, Z.X. First Report of Grapevine Rupestris Vein Feathering Virus in an Old Grapevine in China. Plant Dis. 2017, 101, 848. [Google Scholar] [CrossRef]
- Wu, Q.; Kehoe, M.A.; Kinoti, W.M.; Wang, C.P.; Rinaldo, A.; Tyerman, S.; Habili, N.; Constable, F.E. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Australia. Plant Dis. 2021, 105, 515. [Google Scholar] [CrossRef]
- Xiao, H.; Meng, B. First Report of Grapevine Asteroid Mosaic-Associated Virus and Grapevine Rupestris Vein Feathering Virus in Grapevines in Canada. Plant Dis. 2016, 100, 2175. [Google Scholar] [CrossRef]
- Mahmood, M.; Gentili, A.; Naz, S.; Faggioli, F. First Report of Grapevine Rupestris Vein Feathering Virus in Pakistan. J. Plant Pathol. 2019, 101, 1261. [Google Scholar] [CrossRef] [Green Version]
- Khalili, M.; Zarghani, S.N.; Massart, S.; Dizadji, A.; Olmos, A.; Wetzel, T.; Ruiz-García, A.B. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Iran. J. Plant Pathol. 2020, 102, 1313. [Google Scholar] [CrossRef]
- Aoki, Y.; Suzuki, S. First Report of Grapevine Rupestris Vein Feathering Virus in Vitis vinifera L. from Japan. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Cho, I.S.; Chung, B.N.; Hammond, J.; Moon, J.S.; Lim, H.S. First Report of Grapevine Rupestris Vein Feathering Virus Infecting Grapevines in Korea. Plant Dis. 2018, 102, 1471. [Google Scholar] [CrossRef]
- Mekuria, T.A.; Naidu, R.A. First Report of Grapevine Virus Sequences Highly Similar to Grapevine Syrah Virus-1 from Washington Vineyards. Plant Dis. 2010, 94, 787. [Google Scholar] [CrossRef]
- Engel, E.A.; Rivera, P.A.; Valenzuela, P.D.T. First Report of Grapevine Syrah Virus-1 in Chilean Grapevines. Plant Dis. 2010, 94, 633. [Google Scholar] [CrossRef]
- Ahmed, I.; Fan, X.D.; Zhang, Z.P.; Ren, F.; Hu, G.J.; Li, Z.N.; Khaskheli, M.I.; Dong, Y.F. First Report of Grapevine Syrah Virus-1 in Grapevines in China. Plant Dis. 2017, 102, 466. [Google Scholar] [CrossRef]
- Ruiz-García, A.B.; Sabaté, J.; Lloria, O.; Laviña, A.; Batlle, A.; Olmos, A. First Report of Grapevine Syrah Virus-1 in Grapevine in Spain. Plant Dis. 2017, 101, 1830. [Google Scholar] [CrossRef]
- Cho, I.S.; Yang, C.Y.; Kwon, S.J.; Yoon, J.Y.; Kim, D.H.; Choi, G.S.; Hammond, J.; Moon, J.S.; Lim, H.S. First Report of Grapevine Syrah Virus 1 Infecting Grapevines in Korea. Plant Dis. 2019, 103, 2970. [Google Scholar] [CrossRef]
- Czotter, N.; Szabó, E.; Molnar, J.; Kocsis, L.; Deák, T.; Bisztray, G.; Tusnády, G.E.; Burgyán, J.; Várallyay, É. First Description of Grapevine Syrah Virus 1 in Vineyards of Hungary. J. Plant Pathol. 2015, 97. [Google Scholar] [CrossRef]
- Oosthuizen, K.; Coetzee, B.; Maree, H.J.; Burger, J.T. First Report of Grapevine Syrah Virus 1 in South African Grapevines. Plant Dis. 2016, 100, 1252. [Google Scholar] [CrossRef]
- Caglayan, K.; Gazel, M.; Kocabag, H.D. First Report of Grapevine Syrah Virus 1 in Grapevine in Turkey. J. Plant Pathol. 2017, 99, 303. [Google Scholar]
- Glasa, M.; Predajňa, L.; Šoltys, K.; Sabanadzovic, S.; Olmos, A. Detection and Molecular Characterisation of Grapevine Syrah Virus-1 Isolates from Central Europe. Virus Genes 2015, 51, 112. [Google Scholar] [CrossRef] [PubMed]
- Mavrič Pleško, I.; Lamovšek, J.; Lešnik, A.; Viršček Marn, M. Raspberry Bushy Dwarf Virus in Slovenia—Geographic Distribution, Genetic Diversity and Population Structure. Eur. J. Plant Pathol. 2020, 158, 1033–1042. [Google Scholar] [CrossRef]
- Mavrič, I.; Marn, M.V.; Koron, D.; Žežlina, I. First Report of Raspberry Bushy Dwarf Virus on Red Raspberry and Grapevine in Slovenia. Plant Dis. 2003, 87, 1148. [Google Scholar] [CrossRef]
- Pleško, I.M.; Marn, M.V.; Nyerges, K.; Lázár, J. First Report of Raspberry Bushy Dwarf Virus Infecting Grapevine in Hungary. Plant Dis. 2012, 96, 1582. [Google Scholar] [CrossRef]
- Jevremovic, D.; Paunovic, S. Raspberry Bushy Dwarf Virus: A Grapevine Pathogen in Serbia. Pesticidi i Fitomedicina 2011, 26, 55–60. [Google Scholar] [CrossRef]
- Polivka, H.; Staub, U.; Gross, H.J. Variation of Viroid Profiles in Individual Grapevine Plants: Novel Grapevine Yellow Speckle Viroid 1 Mutants Show Alterations of Hairpin I. J. Gen. Virol. 1996, 77, 155–161. [Google Scholar] [CrossRef]
- Di Serio, F.; Izadpanah, K.; Hajizadeh, M.; Navarro, B. Viroids Infecting the Grapevine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 373–392. [Google Scholar] [CrossRef]
- Cañizares, M.C.; Marcos, J.F.; Pallás, V. Molecular Characterization of an Almond Isolate of Hop Stunt Viroid (HSVd) and Conditions for Eliminating Spurious Hybridization in Its Diagnosis in Almond Samples. Eur. J. Plant Pathol. 1999, 105, 553–558. [Google Scholar] [CrossRef]
- Marquez-Molins, J.; Gomez, G.; Pallas, V. Hop Stunt Viroid: A Polyphagous Pathogenic RNA That Has Shed Light on Viroid–Host Interactions. Mol. Plant Pathol. 2021, 22, 153–162. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, Y.; Guo, R.; Mu, L.; Yang, Y.; Li, S.; Wang, H. Molecular Characterization of Chinese Hop Stunt Viroid Isolates Reveals a New Phylogenetic Group and Possible Cross Transmission between Grapevine and Stone Fruits. Eur. J. Plant Pathol. 2012, 134, 217–225. [Google Scholar] [CrossRef]
- Hataya, T.; Tsushima, T.; Sano, T. Chapter 19—Hop Stunt Viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 199–210. [Google Scholar] [CrossRef]
- Rezaian, M.A. Australian Grapevine Viroid—Evidence for Extensive Recombination between Viroids. Nucleic Acids Res. 1990, 18, 1813–1818. [Google Scholar] [CrossRef] [Green Version]
- Elleuch, A.; Fakhfakh, H.; Pelchat, M.; Landry, P.; Marrakchi, M.; Perreault, J.-P. Sequencing of Australian Grapevine Viroid and Yellow Speckle Viroid Isolated from a Tunisian Grapevine without Passage in an Indicator Plant. Eur. J. Plant Pathol. 2002, 108, 815–820. [Google Scholar] [CrossRef]
- Elleuch, A.; Marrakchi, M.; Perreault, J.P.; Fakhfakh, H. First Report of Australian Grapevine Viroid from the Mediterranean Region. J. Plant Pathol. 2003, 85, 53–57. [Google Scholar]
- Jiang, D.; Peng, S.; Wu, Z.; Cheng, Z.; Li, S. Genetic Diversity and Phylogenetic Analysis of Australian Grapevine Viroid (AGVd) Isolated from Different Grapevines in China. Virus Genes 2009, 38, 178–183. [Google Scholar] [CrossRef]
- Zaki-Aghl, M.; Izadpanah, K.; Niazi, A.; Behjatnia, A.; Afsharifar, A. Molecular and Biological Characterization of the Iranian Isolate of the Australian Grapevine Viroid. J. Agric. Sci. Technol. 2013, 15, 855–865. [Google Scholar]
- Gambino, G.; Navarro, B.; La Notte, P.; Mannini, F.; Di Serio, F. Survey on Viroids Infecting Grapevine in Italy: Identification and Characterization of Australian Grapevine Viroid and Grapevine Yellow Speckle Viroid 2. Available online: https://link.springer.com/article/10.1007%2Fs10658-014-0458-x (accessed on 28 July 2021).
- Adkar-Purushothama, C.R.; Kanchepalli, P.R.; Yanjarappa, S.M.; Zhang, Z.; Sano, T. Detection, Distribution, and Genetic Diversity of Australian Grapevine Viroid in Grapevines in India. Virus Genes 2014, 49, 304–311. [Google Scholar] [CrossRef]
- Saengmanee, P.; Burns, P.; Wetzel, T. First Report of Australian Grapevine Viroid in Grapevine in Thailand. Plant Dis. 2018, 102, 2668. [Google Scholar] [CrossRef]
- Kryovrysanaki, N.; Katsarou, K.; Olmos, A.; Ruiz-García, A.B.; Kalantidis, K.; Pappi, P. First Report of Australian Grapevine Viroid in Grapevine in Greece. J. Plant Pathol. 2021, 103, 1023–1024. [Google Scholar] [CrossRef]
- Barbara, D.J.; Morton, A.; Ramcharan, S.; Cole, I.W.; Phillips, A.; Knight, V.H. Occurrence and Distribution of Raspberry Bushy Dwarf Virus in Commercial Rubus Plantations in England and Wales. Plant Pathol. 2001, 50, 747–754. [Google Scholar] [CrossRef]
- Čepin, U.; Gutiérrez-Aguirre, I.; Ravnikar, M.; Pompe-Novak, M. Frequency of Occurrence and Genetic Variability of Grapevine Fanleaf Virus Satellite RNA. Plant Pathol. 2016, 65, 510–520. [Google Scholar] [CrossRef]
- Cigsar, I.; Digiaro, M.; Gokalp, K.; Ghanem-Sabanadzovic, N.A.; Stradis, A.D.; Boscia, D.; Martell, G.P. Grapevine deformation virus, a novel nepovirus from Turkey. J. Plant Pathol. 2003, 85, 183–191. [Google Scholar] [CrossRef]
- Daldoul, S.; Massart, S.; Ruiz-García, A.B.; Olmos, A.; Wetzel, T. First Report of Grapevine Rupestris Vein Feathering Virus in Grapevine in Germany. Plant Dis. 2018, 102, 2053. [Google Scholar] [CrossRef]
- Gambino, G.; Gribaudo, I. Simultaneous Detection of Nine Grapevine Viruses by Multiplex Reverse Transcription-Polymerase Chain Reaction with Coamplification of a Plant RNA as Internal Control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasa, M.; Predajňa, L.; Komínek, P.; Nagyová, A.; Candresse, T.; Olmos, A. Molecular Characterization of Divergent Grapevine Pinot Gris Virus Isolates and Their Detection in Slovak and Czech Grapevines. Arch. Virol. 2014, 159, 2103–2107. [Google Scholar] [CrossRef] [PubMed]
- Gottula, J.; Lapato, D.; Cantilina, K.; Saito, S.; Bartlett, B.; Fuchs, M. Genetic Variability, Evolution, and Biological Effects of Grapevine Fanleaf Virus Satellite RNAs. Phytopathology 2013, 103, 1180–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajizadeh, M.; Navarro, B.; Bashir, N.S.; Torchetti, E.M.; Di Serio, F. Development and Validation of a Multiplex RT-PCR Method for the Simultaneous Detection of Five Grapevine Viroids. Sci. Direct 2012, 179, 62–69. [Google Scholar] [CrossRef]
- Jiang, D.; Zhang, Z.; Wu, Z.; Guo, R.; Wang, H.; Fan, P.; Li, S. Molecular Characterization of Grapevine Yellow Speckle Viroid-2 (GYSVd-2). Virus Genes 2009, 38, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Osman, F.; Rowhani, A. Application of a Spotting Sample Preparation Technique for the Detection of Pathogens in Woody Plants by RT-PCR and Real-Time PCR (TaqMan). Sci. Direct 2006, 133, 130–136. [Google Scholar] [CrossRef]
- Padilla, C.V.; Cretazzo, E.; Hita, I.; López, N.; Padilla, V.; Velasco, L. First Report of Grapevine Leafroll-Associated Virus 5 in Spain. Plant Dis. 2010, 94, 1507. [Google Scholar] [CrossRef]
- Rowhani, A.; Golino, D.A.; Uyemoto, J.K.; Zhang, Y.P. Isolation and Partial Characterization of Two New Viruses from Grapevine. In Proceedings of the 13th Meeting of the International Council for the Study of Viruses and Virus-Like Diseases of the Grapevine (ICVG), Adelaide, SA, Australia, 12–17 March 2000. [Google Scholar]
- Sano, T.; Mimura, R.; Ohshima, K. Phylogenetic Analysis of Hop and Grapevine Isolates of Hop Stunt Viroid Supports a Grapevine Origin for Hop Stunt Disease. Virus Genes 2001, 22, 53–59. [Google Scholar] [CrossRef]
- Wetzel, T.; Bassler, A.; Golino, D.A.; Uyemoto, J.K. A RT/PCR-Partial Restriction Enzymatic Mapping (PREM) Method for the Molecular Characterisation of the Large Satellite RNAs of Arabis Mosaic Virus Isolates. Sci. Direct 2006, 132, 97–103. [Google Scholar] [CrossRef]
- Cretazzo, E.; Velasco, L. High-throughput Sequencing Allowed the Completion of the Genome of Grapevine Red Globe Virus and Revealed Recurring Co-infection with Other Tymoviruses in Grapevine. Plant Pathol. 2017, 66, 1202–1213. [Google Scholar] [CrossRef]
- Ghanem-Sabanadzovic, N.A.; Sabanadzovic, S.; Castellano, M.A.; Boscia, D.; Martelli, G.P. Properties of a New Isolate of Grapevine Leafroll-Associated Virus 2. Vitis 2000, 39, 119–121. [Google Scholar]
- Naidu, R.A.; Mekuria, T.A. First Report of Grapevine Fleck Virus from Washington Vineyards. Plant Dis. 2010, 94, 784. [Google Scholar] [CrossRef]
- Glasa, M.; Predajňa, L.; Sihelská, N.; Šoltys, K.; Ruiz-García, A.B.; Olmos, A.; Wetzel, T.; Sabanadzovic, S. Grapevine Virus T Is Relatively Widespread in Slovakia and Czech Republic and Genetically Diverse. Virus Genes 2018, 54, 737–741. [Google Scholar] [CrossRef]
- Wetzel, T.; Jardak, R.; Meunier, L.; Ghorbel, A.; Reustle, G.M.; Krczal, G. Simultaneous RT/PCR Detection and Differentiation of Arabis Mosaic and Grapevine Fanleaf Nepoviruses in Grapevines with a Single Pair of Primers. J. Virol. Methods 2002, 101, 63–69. [Google Scholar] [CrossRef]
- Reynard, J.-S.; Brodard, J.; Dubuis, N.; Yobregat, O.; Kominek, P.; Schumpp, O.; Schaerer, S. First Report of Grapevine Rupestris Vein Feathering Virus in Swiss Grapevines. Plant Dis. 2017, 101, 1062. [Google Scholar] [CrossRef]
- Minafra, A.; Hadidi, A. Sensitive Detection of Grapevine Virus A, B, or Leafroll-Associated III from Viruliferous Mealybugs and Infected Tissue by CDNA Amplification. J. Virol. Methods 1994, 47, 175–187. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navrotskaya, E.; Porotikova, E.; Yurchenko, E.; Galbacs, Z.N.; Varallyay, E.; Vinogradova, S. High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses 2021, 13, 2432. https://doi.org/10.3390/v13122432
Navrotskaya E, Porotikova E, Yurchenko E, Galbacs ZN, Varallyay E, Vinogradova S. High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses. 2021; 13(12):2432. https://doi.org/10.3390/v13122432
Chicago/Turabian StyleNavrotskaya, Emiliya, Elena Porotikova, Eugeniya Yurchenko, Zsuzsanna Nagyne Galbacs, Eva Varallyay, and Svetlana Vinogradova. 2021. "High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia" Viruses 13, no. 12: 2432. https://doi.org/10.3390/v13122432
APA StyleNavrotskaya, E., Porotikova, E., Yurchenko, E., Galbacs, Z. N., Varallyay, E., & Vinogradova, S. (2021). High-Throughput Sequencing of Small RNAs for Diagnostics of Grapevine Viruses and Viroids in Russia. Viruses, 13(12), 2432. https://doi.org/10.3390/v13122432