
A Review of the Emerging White Chick Hatchery Disease



Abstract
:1. Introduction
2. Determination of the Causative Agent of White Chick Hatchery Disease
3. Clinical Signs and Disease Progression
4. Histopathology
5. Economic Impact
6. Differential Diagnosis
7. Transmission
8. Prevention, Control and Treatment
9. Methods of Detection
10. Classification of CAstV
11. Genomic Organisation of CAstV
12. Genetic Diversity of CAstV
13. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazaheri, A.; Prusas, C.; Voß, M.; Hess, M. Vertical transmission of fowl adenovirus serotype 4 investigated in specified patho-gen-free birds after experimental infection. Arch. Fur Geflugelkd. 2003, 67, 6–10. [Google Scholar]
- Todd, D.; Wilkinson, D.S.; Jewhurst, H.L.; Wylie, M.; Gordon, A.W.; Adair, B.M. A seroprevalence investigation of chicken astrovirus infections. Avian Pathol. 2009, 38, 301–309. [Google Scholar] [CrossRef]
- Linden, J. Vertically Transmitted Health Issues in Poultry. Available online: https://thepoultrysite.com/articles/vertically-transmitted-health-issues-in-poultry. (accessed on 6 May 2021).
- Smyth, V.; Trudgett, J.; Wylie, M.; Jewhurst, H.; Conway, B.; Welsh, M.; Kaukonen, E.; Perko-Mäkelä, P. Chicken astrovirus detected in hatchability problems associated with ‘white chicks’. Vet. Rec. 2013, 173, 403–404. [Google Scholar] [CrossRef] [PubMed]
- Long, K.E.; Hastie, G.M.; Ojkić, D.; Brash, M.L. Economic impacts of white chick syndrome in Ontario, Canada. Avian Dis. 2017, 61, 402–408. [Google Scholar] [CrossRef]
- Bosch, A.; Guix, S.; Krishna, N.; Al, E. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses, 9th ed.; King, A., Adams, M., Carstens, E., Lefkowitz, E., Eds.; Elsevier: San Diego, CA, USA, 2012. [Google Scholar]
- Nuñez, L.; Parra, S.H.S.; Carranza, C.; Astolfi-Ferreira, C.; Buim, M.; Ferreira, A.P. Detection and molecular characterization of chicken astrovirus associated with chicks that have an unusual condition known as “white chicks” in Brazil. Poult. Sci. 2016, 95, 1262–1270. [Google Scholar] [CrossRef]
- Sajewicz-Krukowska, J.; Pać, K.; Lisowska, A.; Pikuła, A.; Minta, Z.; Króliczewska, B.; Domańska-Blicharz, K. Astrovirus-induced “white chicks” condition—Field observation, virus detection and preliminary characterization. Avian Pathol. 2016, 45, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stayer, P.A. Incursion and recursion of ‘white chicks’ in U.S. commercial broiler production. In Proceedings of the American Association of Avian Pathologists Symposium and Scientific Program, San Antonion, TX, USA, 6–9 August 2016. [Google Scholar]
- Smyth, V.; Kaukonen, E.; Trudgett, J.; Welsh, M.; Todd, D. Chicken astrovirus detected in hatchability problems associated with ‘white chicks’. In Proceeding of the XVIIth Congress 2013 WVPA (World Veterinary Poultry Association), Nantes, France, 19–23 August 2013. [Google Scholar]
- Long, K.E.; Ouckama, R.M.; Weisz, A.; Brash, M.L.; Ojkic, D. White chick syndrome associated with chicken astrovirus in Ontario, Canada. Avian Dis. 2018, 62, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Baxendale, W.; Mebatsion, T. The isolation and characterisation of astroviruses from chickens. Avian Pathol. 2004, 33, 364–370. [Google Scholar] [CrossRef]
- Hoerr, F.; Lockaby, S.B.; Bickford, A.A. Pathology of Avian Mycoplasmosis. In Poultry Mycoplasma Workshop; California Veterinary Diagnostic Laboratory System: Davis, CA, USA, 1994; pp. 89–100. [Google Scholar]
- Abdisa, T.; Tagesu, T. Review on Newcastle disease of poultry and its public health importance. J. Vet. Sci. Technol. 2017, 8, 441. [Google Scholar] [CrossRef]
- Miller, P.J.; Koch, G. Newcastle Disease. Dis. Poult. 2013, 13, 89–138. [Google Scholar]
- Grgić, H.; Philippe, C.; Ojkić, D.; Nagy, E. Study of vertical transmission of fowl adenoviruses. Can. J. Vet. Res. Rev. Can. de Rech. Vet. 2006, 70, 230–233. [Google Scholar]
- van der Heide, L.; Kalbac, M. Infectious tenosynovitis (viral arthritis): Characterization of a connecticut viral isolant as a reovi-rus and evidence of viral egg transmission by reovirus-infected broiler breeders. Avian Dis. 1975, 19, 683–688. [Google Scholar] [CrossRef]
- Al-Muffarej, S.I.; Savage, C.E.; Jones, R.C. Egg transmission of avian reoviruses in chickens: Comparison of a trypsin-sensitive and a trypsin-resistant strain. Avian Pathol. 1996, 25, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.A.; Deloach, J.R.; Corrier, D.E.; Nisbet, D.J.; Stanker, L.H. Evaluation of Salmonella serotype distributions from commercial broiler hatcheries and grower houses. Avian Dis. 1999, 43, 39–47. [Google Scholar] [CrossRef]
- Bygrave, A.C.; Gallagher, J. Transmission of Salmonella enteritidis in poultry. Vet. Rec. 1989, 124, 571. [Google Scholar] [CrossRef] [PubMed]
- Kouwenhoven, B.; Vertommen, M.; Van Eck, J.H.H. Runting and leg weakness in broilers; Involvement of infectious factors. Vet. Res. Commun. 1978, 2, 253–259. [Google Scholar] [CrossRef]
- Tung, H.-T.; Hamilton, P.B. Decreased plasma carotenoids during aflatoxicosis. Poult. Sci. 1973, 52, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Lilburn, M.S.; Edwards, H.M., Jr.; Jensen, L.S. Impaired nutrient utilization associated with pale bird syndrome in broiler chicks. Poult. Science 1982, 61, 608–609. [Google Scholar]
- Nelson, T.S.; Kirby, L.K.; Johnson, Z.B.; Halley, J.T.; Beasley, J.N. Digestion of feed by chicks with pale bird syndrome. Avian Dis. 1982, 26, 696. [Google Scholar] [CrossRef]
- Schaeffer, J.L.; Tyczkowski, J.K.; Riviere, J.E.; Hamilton, P.B. Aflatoxin-impaired ability to accumulate oxycarotenoid pigments during restoration in young chickens. Poult. Sci. 1988, 67, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Tyczkowski, J.K.; Schaeffer, J.L.; Hamilton, P.B. Measurement of malabsorption of carotenoids in chickens with pale-bird syndrome. Poult. Sci. 1991, 70, 2275–2279. [Google Scholar] [CrossRef]
- Osborne, D.J.; Huff, W.E.; Hamilton, P.B.; Burmeister, H.R. Comparison of ochratoxin, aflatoxin, and T-2 toxin for their effects on selected parameters related to digestion and evidence for specific metabolism of carotenoids in chickens. Poult. Sci. 1982, 61, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.D. The Role of Coccidia in Malabsorption Syndrome. In Poultry Nutrition Short Course; University of Delaware, Cooperative Extension Service: Newark, DE, USA, 1984; pp. 6–16. [Google Scholar]
- Van Loon, A.; Koopman, H.; Kosman, W.; Mumczur, J.; Szeleszczuk, O.; Karpinska, E.; Kosowska, G.; Lütticken, D. Virology: Isolation of a new serotype of avian reovirus associated with malabsorption syndrome in chickens. Vet. Q. 2001, 23, 129–133. [Google Scholar] [CrossRef]
- Kouwenhoven, B.; Vertommen, M.; Goren, E. Investigations into the role of reovirus in the malabsorption syndrome. Avian Pathol. 1988, 17, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Huff, W.E.; Hamilton, P.B. Decreased plasma carotenoids during ochratoxicosis. Poult. Sci. 1975, 54, 1308–1310. [Google Scholar] [CrossRef] [PubMed]
- Mettifogo, E.; Nuñez, L.F.N.; Chacón, J.L.; Parra, S.H.S.; Astolfi-Ferreira, C.; Jerez, J.A.; Jones, R.C.; Ferreira, A.J.P. Emergence of enteric viruses in production chickens is a concern for avian health. Sci. World J. 2014, 2014, 450423. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.; Lee, T. Astroviruses: Human and animal. Novartis Found. Symp. 2007, 128, 92–107. [Google Scholar] [CrossRef]
- Smyth, V.J. A Review of the strain diversity and pathogenesis of chicken astrovirus. Viruses 2017, 9, 29. [Google Scholar] [CrossRef]
- Adebiyi, A.I.; Tregaskis, P.L.; Oluwayelu, D.O.; Smyth, V.J. Investigation of Enteric Viruses Associated with Runting and Stunting in Day-Old Chicks and Older Broilers in Southwest Nigeria. Front. Veter- Sci. 2019, 6, 239. [Google Scholar] [CrossRef] [Green Version]
- Koci, M.D.; Schultz-Cherry, S. Avian astroviruses. Avian Pathol. 2002, 31, 213–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazeleger, W.C.; Bolder, N.M.; Beumer, R.R.; Jacobs-Reitsma, W.F. Darkling beetles (Alphitobius diaperinus) and their larvae as potential vectors for the transfer of Campylobacter jejuni and Salmonella enterica Serovar Paratyphi B variant java between Successive broiler flocks. Appl. Environ. Microbiol. 2008, 74, 6887–6891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skibińska, A.; Lee, A.; Wylie, M.; Smyth, V.J.; Welsh, M.D.; Todd, D. Development of an indirect ELISA test for detecting antibodies to chicken astrovirus in chicken sera. Avian Pathol. 2015, 44, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Madeley, C.; Cosgrove, B. 28 nm particles in faeces in infantile gastroenteritis. Lancet 1975, 306, 451–452. [Google Scholar] [CrossRef]
- Todd, D.; Smyth, V.; Ball, N.W.; Donnelly, B.M.; Wylie, M.; Knowles, N.J.; Adair, B.M. Identification of chicken enterovirus-like viruses, duck hepatitis virus type 2 and duck hepatitis virus type 3 as astroviruses. Avian Pathol. 2009, 38, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Smyth, V.; Jewhurst, H.L.; Wilkinson, D.S.; Adair, B.M.; Gordon, A.W.; Todd, D. Development and evaluation of real-time TaqMan®RT-PCR assays for the detection of avian nephritis virus and chicken astrovirus in chickens. Avian Pathol. 2010, 39, 467–474. [Google Scholar] [CrossRef]
- Palomino-Tapia, V.; Mitevski, D.; Inglis, T.; Van Der Meer, F.; Martin, E.; Brash, M.; Provost, C.; Gagnon, C.A.; Abdul-Careem, M.F. Chicken Astrovirus (CAstV) molecular studies reveal evidence of multiple past recombination events in sequences originated from clinical samples of White Chick Syndrome (WCS) in western Canada. Viruses 2020, 12, 1096. [Google Scholar] [CrossRef] [PubMed]
- Sajewicz-Krukowska, J.; Domanska-Blicharz, K. Nearly full-length genome sequence of a novel astrovirus isolated from chickens with ‘white chicks’ condition. Arch. Virol. 2016, 161, 2581–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mutairy, B.; Walter, J.E.; Pothen, A.; Mitchell, D.K. Genome prediction of putative genome-linked Viral Protein (VPg) of astroviruses. Virus Genes 2005, 31, 21–30. [Google Scholar] [CrossRef]
- Lewis, T.L.; Matsui, S.M. An astrovirus frameshift signal induces ribosomal frameshifting in vitro. Arch. Virol. 1995, 140, 1127–1135. [Google Scholar] [CrossRef]
- Jiang, B.; Monroe, S.; Koonin, E.V.; Stine, S.E.; Glass, R.I. RNA sequence of astrovirus: Distinctive genomic organization and a putative retrovirus-like ribosomal frameshifting signal that directs the viral replicase synthesis. Proc. Natl. Acad. Sci. USA 1993, 90, 10539–10543. [Google Scholar] [CrossRef] [Green Version]
- Monroe, S.S.; Jiang, B.; Stine, S.E.; Koopmans, M.; Glass, R.I. Subgenomic RNA sequence of human astrovirus supports classification of astroviridae as a new family of RNA viruses. J. Virol. 1993, 67, 3611–3614. [Google Scholar] [CrossRef] [Green Version]
- Smyth, V.J.; Jewhurst, H.L.; Adair, B.M.; Todd, D. Detection of chicken astrovirus by reverse transcriptase-polymerase chain reaction. Avian Pathol. 2009, 38, 293–299. [Google Scholar] [CrossRef]
- Smyth, V.J.; Todd, D.; Trudgett, J.; Lee, A.; Welsh, M.D. Capsid protein sequence diversity of chicken astrovirus. Avian Pathol. 2012, 41, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulbule, N.R.; Mandakhalikar, K.D.; Kapgate, S.S.; Deshmukh, V.V.; Schat, K.A.; Chawak, M.M. Role of chicken astrovirus as a causative agent of gout in commercial broilers in India. Avian Pathol. 2013, 42, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, F.; Connor, T.J.; Calvert, V.M.; Smyth, J.A.; Curran, W.L.; Morley, A.J.; Thompson, D.; Singh, S.; McFerran, J.B.; Adair, B.M.; et al. Studies on a new enterovirus-like virus isolated from chickens. Avian Pathol. 1994, 23, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Spackman, D.; Gough, R.E.; Collins, M.S.; Lanning, D. Isolation of an enterovirus-like agent from the meconium of dead-in-shell chicken embryos. Vet. Rec. 1984, 114, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-I.; Icard, A.H.; Linnemann, E.; Sellers, H.S.; Mundt, E. Determination of the full length sequence of a chicken astrovirus suggests a different replication mechanism. Virus Genes 2012, 44, 45–50. [Google Scholar] [CrossRef]
- Kang, K.-I.; Linnemann, E.; Icard, A.H.; Durairaj, V.; Mundt, E.; Sellers, H.S. Chicken astrovirus as an aetiological agent of runting-stunting syndrome in broiler chickens. J. Gen. Virol. 2018, 99, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef]
- Martin, D.; Posada, D.; Crandall, K.; Williamson, C. A modified bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. AIDS Res. Hum. Retroviruses 2005, 21, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuBois, R.; Freiden, P.; Marvin, S.; Reddivari, M.; Heath, R.J.; White, S.W.; Schultz-Cherry, S. Crystal structure of the avian astrovirus capsid spike. J. Virol. 2013, 87, 7853–7863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jindal, N.; Patnayak, D.P.; Chander, Y.; Ziegler, A.F.; Goyal, S.M. Comparison of capsid gene sequences of turkey astrovirus-2 from poult-enteritis-syndrome-affected and apparently healthy turkeys. Arch. Virol. 2011, 156, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, L.L.C. The {PyMOL} Molecular Graphics System. Version 1.8. 2015. Available online: https://pymol.org/2/support.html?#citing (accessed on 3 December 2021).






{kind=link}
{kind=link}
{kind=link}
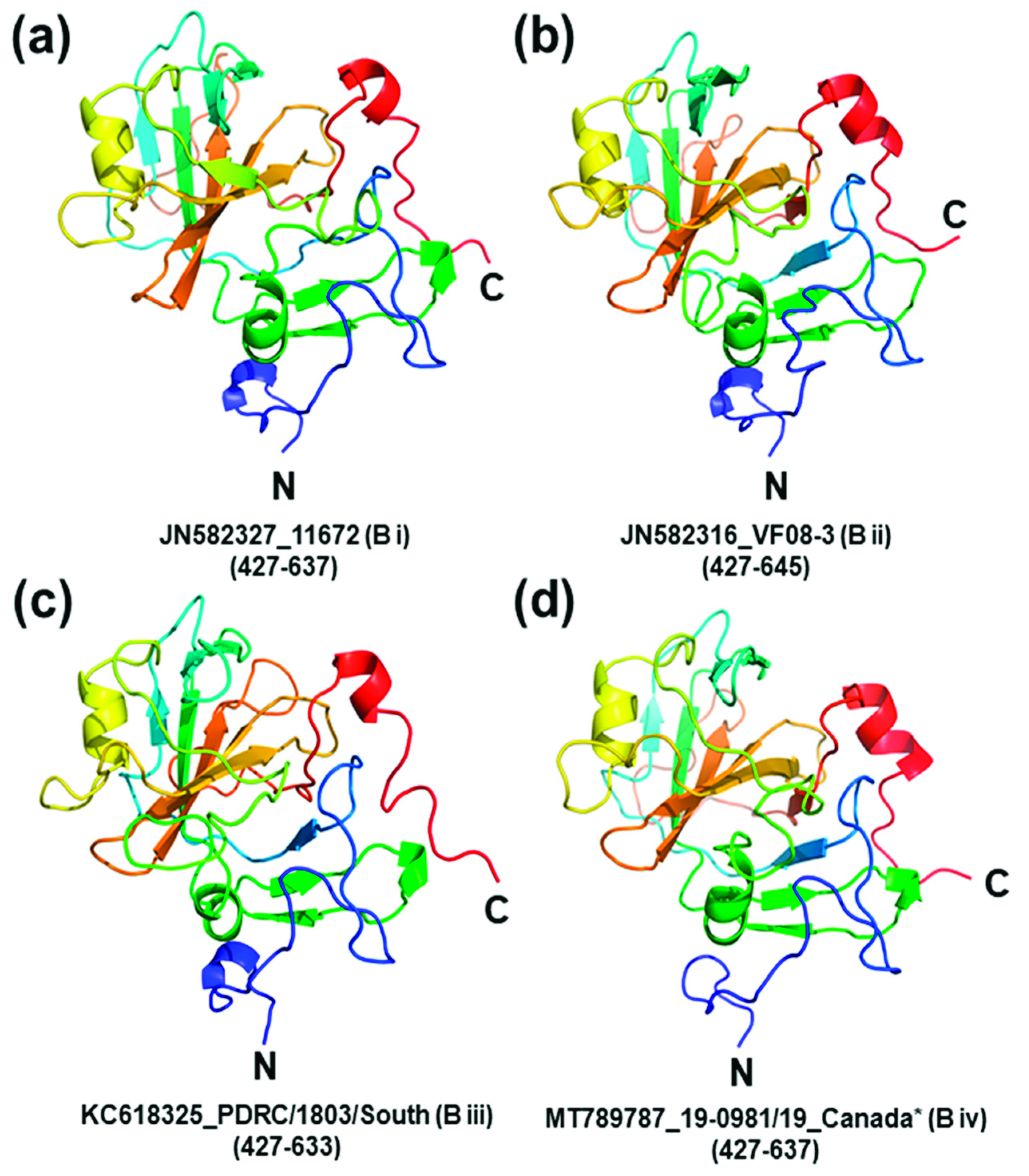{kind=link}
{kind=link}
| CAstV Subgroup | Strain | Accession Number | Associated Disease | Clinical Signs and/or Experimental Findings | Organs Infected | References |
|---|---|---|---|---|---|---|
| Ai | 612 | EU669001 | Originally isolated from broilers suffering from respiratory distress | Growth retardation | Duodenum, jejunum and kidney. Isolated from birds suffering from respiratory distress | [40,52] |
| Aii | VF08-54 | JN582323 | N/A | Underperformance | Intestines | [49] |
| VF08-36 | JN582325 | |||||
| Aiii | VF08-46 | JN582321 | N/A | Underperformance | Intestines | [49] |
| PL/G059/2014 | KR052479 | White chick hatchery disease | Adult birds—no apparent effect Chicks—runted, weak chicks, white plumage, decreased life expectancy and a decrease in egg hatchability | Liver, kidneys, pancreas, spleen | [8] Sajewicz-Krukowska et al., 2016 | |
| Bi | NGR_CAstV_Ch1 | MK509014 | Hatchery disease | Growth retardation, weakness, dullness, ruffled/wet feathers, splayed legs and hatchability problems | Intestines | [35] |
| FP3 | JN582328 | Hatchery disease | Dead-in-shell embryos and weak chicks | Small intestine, kidney, pancreas | [53] | |
| Bii | VF08-29 | JN582315 | N/A | Underperformance | Intestines | [49] |
| GA2011 | JF414802 | Runting stunting syndrome (RSS) | Ruffled feathers, growth retardation and diarrhoea | Small intestine | [54] | |
| Biii | PDRC 588 | JX945866 | Visceral gout and kidney disease | Embryo stunting, swollen, pale kidneys, liver necrosis, leading to kidney disease and visceral gout | Kidneys and liver | [50] |
| PDRC 1804 | KC618324 | |||||
| PDRC 579 | JX945864 | |||||
| Biv | 16-028568-0005 a | KY635984 a | White chick hatchery disease | Adult birds—The health status of adults is normal other than occasional reduction in egg-laying. Birds test positive for antibodies against the CAstV B group Chicks and embryos—runted, weak chicks, white plumage, decreased life expectancy, decrease in hatchability, yolk sac remnant, oedema on the neck and head, subcutaneous oedema and enlarged, green, mottled livers | Adults—Cloacal cells Chicks—Liver, kidneys, intestines, pancreas, bursa of Fabricius spleen, yolk, gizzards, heart, brain, lungs and proventriculus Embryos—liver, kidneys, intestines and yolk | [11] |
| 18-0942/18 | MT789785 | [42] | ||||
| USP541-15 a,b | KR013249 a,b | Additional findings in chicks—Pale kidneys, pancreas, beaks and legs | [7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McIlwaine, K.; Law, C.J.; Lemon, K.; Grant, I.R.; Smyth, V.J. A Review of the Emerging White Chick Hatchery Disease. Viruses 2021, 13, 2435. https://doi.org/10.3390/v13122435
McIlwaine K, Law CJ, Lemon K, Grant IR, Smyth VJ. A Review of the Emerging White Chick Hatchery Disease. Viruses. 2021; 13(12):2435. https://doi.org/10.3390/v13122435
Chicago/Turabian StyleMcIlwaine, Kerry, Christopher J. Law, Ken Lemon, Irene R. Grant, and Victoria J. Smyth. 2021. "A Review of the Emerging White Chick Hatchery Disease" Viruses 13, no. 12: 2435. https://doi.org/10.3390/v13122435
APA StyleMcIlwaine, K., Law, C. J., Lemon, K., Grant, I. R., & Smyth, V. J. (2021). A Review of the Emerging White Chick Hatchery Disease. Viruses, 13(12), 2435. https://doi.org/10.3390/v13122435