
Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Chickens
2.3. Experimental Procedures
2.3.1. Grouping and Viral Challenge
2.3.2. Clinical Observations and Swab Collection
2.3.3. Postmortem Examination and Sample Collection
2.3.4. Assessment of the Development of the Ovary and Oviduct
2.4. Techniques
2.4.1. IBV Genome Load Quantification
2.4.2. Immunofluorescence Assay
2.4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4.4. Histopathology
2.4.5. Statistical Analysis
3. Results
3.1. Clinical Findings
3.2. Viral Shedding
3.3. Serology
3.4. IBV Genome Load in Tissues
3.5. Immunofluorescence Staining
3.6. Gross Pathology and Development of the Reproductive Tract
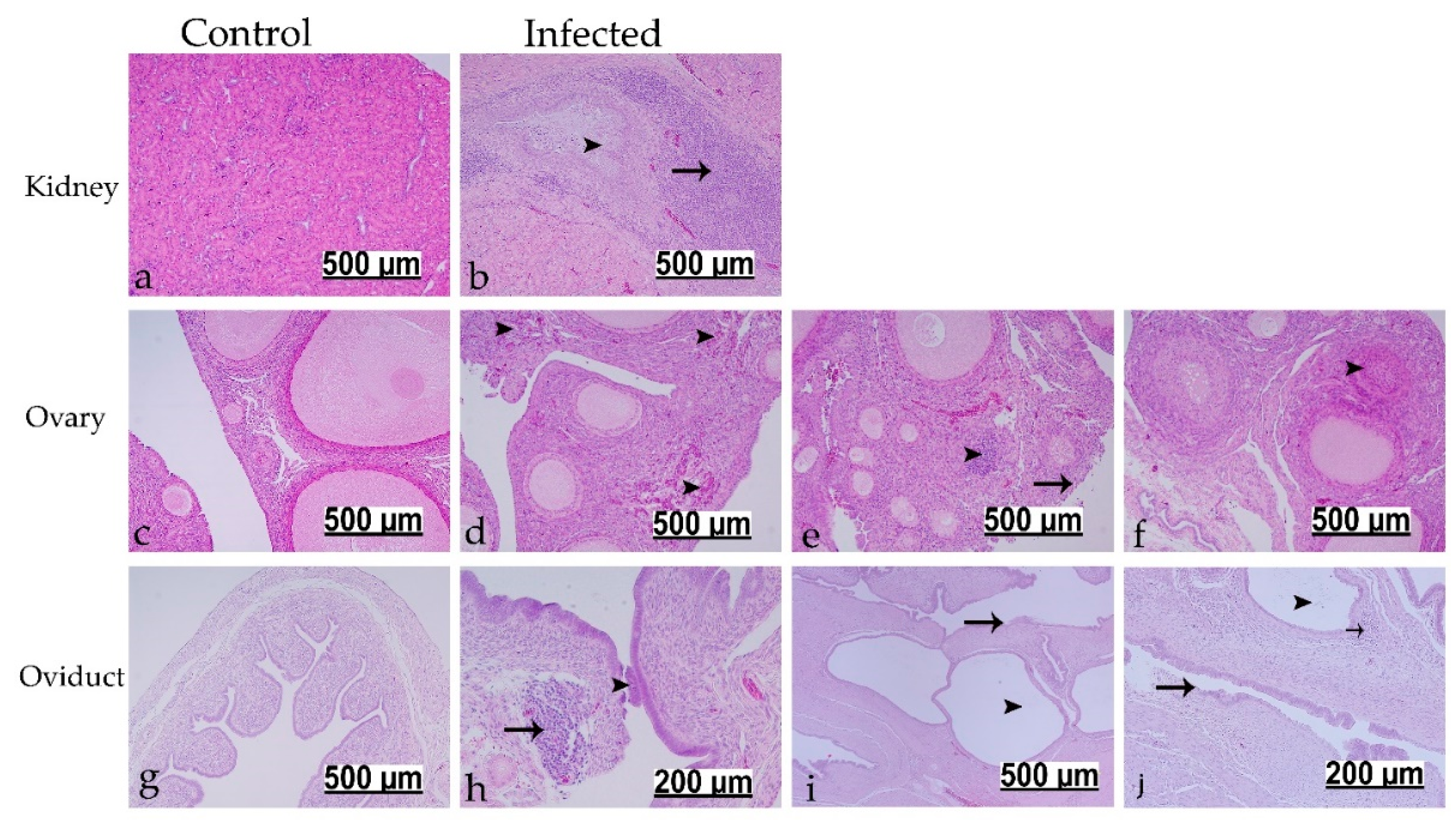
3.7. Histopathological Findings
3.7.1. Histopathological Findings at 7 dpi
3.7.2. Histopathological Findings at 112 dpi
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Hair-Bejo, M.; Mahmuda, A.; Nair, V. Global distributions and strain diversity of avian infectious bronchitis virus: A review. Anim. Health Res. Rev. 2017, 18, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Raj, G.D.; Jones, R.C. Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol. 1997, 26, 677–706. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Abdul-Cader, M.S.; Nazir, S.; De Silva Senapathi, U.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Infectious bronchitis corona virus establishes productive infection in avian macrophages interfering with selected antimicrobial functions. PLoS ONE 2017, 12, e0181801. [Google Scholar] [CrossRef]
- Reddy, V.R.; Trus, I.; Desmarets, L.M.; Li, Y.; Theuns, S.; Nauwynck, H.J. Productive replication of nephropathogenic infectious bronchitis virus in peripheral blood monocytic cells, a strategy for viral dissemination and kidney infection in chickens. Vet. Res. 2016, 47, 70. [Google Scholar] [CrossRef] [Green Version]
- Najimudeen, S.M.; Hassan, M.S.H.; Cork, S.C.; Abdul-Careem, M.F. Infectious Bronchitis Coronavirus Infection in Chickens: Multiple System Disease with Immune Suppression. Pathogens 2020, 9, 779. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef]
- Jackwood, M.W.; Hall, D.; Handel, A. Molecular evolution and emergence of avian gammacoronaviruses. Infect. Genet. Evol. 2012, 12, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; van Santen, V.L.; Jackwood, M.W. Genetic diversity and selection regulates evolution of infectious bronchitis virus. Avian Dis. 2012, 56, 449–455. [Google Scholar] [CrossRef]
- van Roeckel, H.; Bullis, K.L.; Flint, O.S.; Clarke, M.K. Poultry Disease Control Service; Annual Report 1942, Bulletin 388; Massachusetts Agricultural Experiment Station: Amherst, MA, USA; pp. 99–103.
- Jordan, B. Vaccination against infectious bronchitis virus: A continuous challenge. Vet. Microbiol. 2017, 206, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, J.; Han, Z.; Zhang, Q.; Shao, Y.; Kong, X.; Tong, G. Infectious bronchitis virus: S1 gene characteristics of vaccines used in China and efficacy of vaccination against heterologous strains from China. Avian Pathol. 2006, 35, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Beato, M.S.; De Battisti, C.; Terregino, C.; Drago, A.; Capua, I.; Ortali, G. Evidence of circulation of a Chinese strain of infectious bronchitis virus (QXIBV) in Italy. Vet. Rec. 2005, 156, 720. [Google Scholar] [CrossRef]
- Toffan, A.; Monne, I.; Terregino, C.; Cattoli, G.; Hodobo, C.T.; Gadaga, B.; Makaya, P.V.; Mdlongwa, E.; Swiswa, S. QX-like infectious bronchitis virus in Africa. Vet. Rec. 2011, 169, 589. [Google Scholar] [CrossRef]
- Winterfield, R.W.; Hitchner, S.B. Etiology of an infectious nephritis-nephrosis syndrome of chickens. Am. J. Vet. Res. 1962, 23, 1273–1279. [Google Scholar]
- Ziegler, A.F.; Ladman, B.S.; Dunn, P.A.; Schneider, A.; Davison, S.; Miller, P.G.; Lu, H.; Weinstock, D.; Salem, M.; Eckroade, R.J.; et al. Nephropathogenic infectious bronchitis in Pennsylvania chickens 1997–2000. Avian Dis. 2002, 46, 847–858. [Google Scholar] [CrossRef]
- Gelb, J., Jr.; Ladman, B.S.; Pope, C.R.; Ruano, J.M.; Brannick, E.M.; Bautista, D.A.; Coughlin, C.M.; Preskenis, L.A. Characterization of nephropathogenic infectious bronchitis virus DMV/1639/11 recovered from Delmarva broiler chickens in 2011. Avian Dis. 2013, 57, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.B. THE ETIOLOGY OF “URAEMIA” OF CHICKENS. Aust. Vet. J. 1962, 38, 554. [Google Scholar] [CrossRef]
- Chong, K.T.; Apostolov, K. The pathogenesis of nephritis in chickens induced by infectious bronchitis virus. J. Comp. Pathol. 1982, 92, 199–211. [Google Scholar] [CrossRef]
- Sevoian, M.; Levine, P.P. Effects of Infectious Bronchitis on the Reproductive Tracts, Egg Production, and Egg Quality of Laying Chickens. Avian Dis. 1957, 1, 136–164. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Popowich, S.; De Silva Senapathi, U.; Abdul-Cader, M.S.; Marshall, F.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Shell-Less Egg Syndrome (SES) Widespread in Western Canadian Layer Operations Is Linked to a Massachusetts (Mass) Type Infectious Bronchitis Virus (IBV) Isolate. Viruses 2018, 10, 437. [Google Scholar] [CrossRef] [Green Version]
- Chousalkar, K.K.; Roberts, J.R. Effects of Australian strains of infectious bronchitis virus on internal and external quality of hen eggs. Anim. Prod. Sci. 2009, 49, 162–169. [Google Scholar] [CrossRef]
- Crinion, R.A.; Ball, R.A.; Hofstad, M.S. Abnormalities in laying chickens following exposure to infectious bronchitis virus at one day old. Avian Dis. 1971, 15, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Crinion, R.A.; Hofstad, M.S. Pathogenicity of four serotypes of avian infectious bronchitis virus for the oviduct of young chickens of various ages. Avian Dis. 1972, 16, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Benyeda, Z.; Mato, T.; Suveges, T.; Szabo, E.; Kardi, V.; Abonyi-Toth, Z.; Rusvai, M.; Palya, V. Comparison of the pathogenicity of QX-like, M41 and 793/B infectious bronchitis strains from different pathological conditions. Avian Pathol. 2009, 38, 449–456. [Google Scholar] [CrossRef] [Green Version]
- de Wit, J.J.; Nieuwenhuisen-van Wilgen, J.; Hoogkamer, A.; van de Sande, H.; Zuidam, G.J.; Fabri, T.H. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Broadfoot, D.I.; Pomeroy, B.S.; Smith, W.M. Effects of Infectious Bronchitis in Baby Chicks. Poult. Sci. 1956, 35, 757–762. [Google Scholar] [CrossRef]
- Crinion, R.A.; Hofstad, M.S. Pathogenicity of two embryo-passage levels of avian infectious bronchitis virus for the oviduct of young chickens of various ages. Avian Dis. 1972, 16, 967–973. [Google Scholar] [CrossRef]
- Martin, E.; Brash, M.; Stalker, M.; Ojkic, D. Using phylogenetic analysis to examine the changing strains of infectious bronchitis virus infections in Ontario over time. In Proceedings of the 16th Annual Meeting of the Canadian Animal Health Laboratorians Network, Guelph, ON, Canada, 4–7 June 2017. [Google Scholar]
- Hassan, M.S.H.; Ojkic, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Abdul-Careem, M.F. Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Variants Isolated in Eastern Canada Show Evidence of Recombination. Viruses 2019, 11, 1054. [Google Scholar] [CrossRef] [Green Version]
- Petrik, M. Clinical Presentation of False Layer Syndrome Caused by Infectious Bronchitis. In Proceedings of the AVMA/American Association of Avian Pathology Annual Meeting, Denver, CO, USA, 13–17 July 2018. [Google Scholar]
- Boulianne, M.; Buczinski, S.; Lanthier, B. Use of Ultrasound to Diagnose Cystic Oviducts in Infectious Bronchitis Virus (DMV Strain) Positive Immature Leghorns. In Proceedings of the AVMA/American Association of Avian Pathology Annual Meeting, Denver, CO, USA, 13–17 July 2018. [Google Scholar]
- Steimling, C.A.; Wallner-Pendleton, E.A.; Sharafeldin, T.A.; Lu, H.; Williams, J.L.; Dunn, P.A.; Owen, R.L.; Slater, M.; Ladman, B.; Roh, H. Infectious Bronchitis in Immunologically Naïve Breeding Chickens and Their Progeny. In Proceedings of the AVMA/American Association of Avian Pathology Annual Meeting, Washington, DC, USA, 2–5 August 2019. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Cheng, J.; Huo, C.; Zhao, J.; Liu, T.; Li, X.; Yan, S.; Wang, Z.; Hu, Y.; Zhang, G. Pathogenicity differences between QX-like and Mass-type infectious bronchitis viruses. Vet. Microbiol. 2018, 213, 129–135. [Google Scholar] [CrossRef] [PubMed]
- De Silva Senapathi, U.; Abdul-Cader, M.S.; Amarasinghe, A.; van Marle, G.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. The In Ovo Delivery of CpG Oligonucleotides Protects against Infectious Bronchitis with the Recruitment of Immune Cells into the Respiratory Tract of Chickens. Viruses 2018, 10, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.M.; An, S.H.; Lee, C.Y.; Song, C.S.; Choi, K.S.; Kim, J.H.; Kwon, H.J. Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII). Viruses 2018, 10, 652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis corona virus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Q.; Hu, Y.X.; Jin, J.H.; Zhao, Y.; Zhao, J.; Zhang, G.Z. Pathogenicity of virulent infectious bronchitis virus isolate YN on hen ovary and oviduct. Vet. Microbiol. 2016, 193, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liao, K.; Chen, S.; Yan, K.; Du, X.; Zhang, C.; Guo, M.; Wu, Y. Evaluation of the reproductive system development and egg-laying performance of hens infected with TW I-type infectious bronchitis virus. Vet. Res. 2020, 51, 95. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, L.; Zhao, W.; Liu, L.; Zhao, Y.; Shao, Y.; Li, H.; Han, Z.; Liu, S. Identification and molecular characterization of a novel serotype infectious bronchitis virus (GI-28) in China. Vet. Microbiol. 2017, 198, 108–115. [Google Scholar] [CrossRef]
- Boroomand, Z.; Asasi, K.; Mohammadi, A. Pathogenesis and tissue distribution of avian infectious bronchitis virus isolate IRFIBV32 (793/B serotype) in experimentally infected broiler chickens. Sci. World J. 2012, 2012, 402537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J. Detection of infectious bronchitis virus. Avian Pathol. 2000, 29, 71–93. [Google Scholar] [CrossRef] [Green Version]
- da Silva, A.P.; Hauck, R.; Nociti, S.R.C.; Kern, C.; Shivaprasad, H.L.; Zhou, H.; Gallardo, R.A. Molecular Biology and Pathological Process of an Infectious Bronchitis Virus with Enteric Tropism in Commercial Broilers. Viruses 2021, 13, 1477. [Google Scholar] [CrossRef]
- Mast, J.; Goddeeris, B.M. Development of immunocompetence of broiler chickens. Vet. Immunol. Immunopathol. 1999, 70, 245–256. [Google Scholar] [CrossRef]
- Bhattacharjee, P.S.; Carter, S.D.; Savage, C.E.; Jones, R.C. Re-excretion of infectious bronchitis virus in chickens induced by cyclosporin. Avian Pathol. 1995, 24, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Gough, R.E. A long-term study of the pathogenesis of infection of fowls with three strains of avian infectious bronchitis virus. Res. Vet. Sci. 1978, 24, 228–233. [Google Scholar] [CrossRef]
- Naqi, S.; Gay, K.; Patalla, P.; Mondal, S.; Liu, R. Establishment of persistent avian infectious bronchitis virus infection in antibody-free and antibody-positive chickens. Avian Dis. 2003, 47, 594–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, N.A.; Alessi, A.C.; Montassier, H.J.; Pereira, R.J.G.; Taniwaki, S.A.; Botosso, V.F.; Rui, B.R.; Richtzenhain, L.J. Gonadal pathogenicity of an infectious bronchitis virus strain from the Massachusetts genotype. Braz. J. Microbiol. 2019, 50, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crinion, R.A.; Ball, R.A.; Hofstad, M.S. Pathogenesis of oviduct lesions in immature chickens following exposure to infectious bronchitis virus at one day old. Avian Dis. 1971, 15, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Benyeda, Z.; Szeredi, L.; Mató, T.; Süveges, T.; Balka, G.; Abonyi-Tóth, Z.; Rusvai, M.; Palya, V. Comparative histopathology and immunohistochemistry of QX-like, Massachusetts and 793/B serotypes of infectious bronchitis virus infection in chickens. J. Comp. Pathol. 2010, 143, 276–283. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Chickens | Lesion Score (Cystic Formation) in Oviduct A | |||
|---|---|---|---|---|---|
| − | + | ++ | +++ | ||
| Infected | 13 | 7 | 1 | 4 * | 1 |
| Control | 15 | 15 * | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.S.H.; Ali, A.; Buharideen, S.M.; Goldsmith, D.; Coffin, C.S.; Cork, S.C.; van der Meer, F.; Boulianne, M.; Abdul-Careem, M.F. Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens. Viruses 2021, 13, 2488. https://doi.org/10.3390/v13122488
Hassan MSH, Ali A, Buharideen SM, Goldsmith D, Coffin CS, Cork SC, van der Meer F, Boulianne M, Abdul-Careem MF. Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens. Viruses. 2021; 13(12):2488. https://doi.org/10.3390/v13122488
Chicago/Turabian StyleHassan, Mohamed S. H., Ahmed Ali, Sabrina M. Buharideen, Dayna Goldsmith, Carla S. Coffin, Susan C. Cork, Frank van der Meer, Martine Boulianne, and Mohamed Faizal Abdul-Careem. 2021. "Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens" Viruses 13, no. 12: 2488. https://doi.org/10.3390/v13122488
APA StyleHassan, M. S. H., Ali, A., Buharideen, S. M., Goldsmith, D., Coffin, C. S., Cork, S. C., van der Meer, F., Boulianne, M., & Abdul-Careem, M. F. (2021). Pathogenicity of the Canadian Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) on Female Reproductive Tract of Chickens. Viruses, 13(12), 2488. https://doi.org/10.3390/v13122488