1. Introduction
Muscodor albus is a novel endophytic fungus, isolated from
Cinnamomum zeylanicum, that produces over two dozen volatile organic compounds (VOCs) that demonstrate potent antimicrobial activity against a wide range of both human and plant pathogens [
1,
2]. These VOCs were characterized using gas chromatography/mass spectrometry (GC/MS) to determine the identities and amounts of each individual compound. It was established that the small molecular weight organic acid, isobutyric acid, was central to the bioactivity of the VOCs [
2,
3]. However, when isobutyric acid was placed in combination with one or more complex esters that had little or no activity alone (depending on the target organism), antimicrobial activity was substantially increased and resulted in a synergistic inhibitory response [
4]. Of note, when isobutyric acid was combined in the laboratory with the esters in the same relative molar ratio that they were measured via GC/MS, the “artificial” mixture virtually mimicked the activity of the fungus itself [
2,
3]. This antimicrobial activity was predominantly characterized utilizing the
M. albus compounds in the volatile state [
1,
2].
Subsequently, for utility and intellectual property purposes, and after extensive testing, it was learned that propionic acid could be substituted for isobutyric acid and isoamyl hexanoates (two esters of the 2 and 3 isomers of the alcohol) could serve as a substitute for other esters, aldehydes and alcohols in the
M. albus mixture without compromising biological activity. Interestingly, neither the propionic acid nor the isoamyl hexanoates have ever been noted as products of
Muscodor sp. In the gas phase as well as in MIC (minimum inhibitory concentration) tests, the propionic acid/isoamyl hexanoate mixture, now named Propylamylatin formula, was biologically active against a wide range of both human and plant pathogenic fungi and bacteria, including such microbes as
Listeria sp.,
Salmonella sp., drug-resistant
Staphyloccus aureus,
Clostridium sp.,
Xanthomonas sp.
Candida albicans,
Pythium ultimum, Rhizoctonia solani, and many others [
5]. The Propylamylatin formula also targeted many other bacteria and fungi that are relatively resistant to antimicrobial interventions, including
Trichoderma sp and
Lactobacillus sp. [
5]. Unfortunately, the exact mode of action of the formula is unknown, but it has been suggested that one or more physio-chemical mechanisms involving membrane proteins might be involved.
Despite the well-documented antibacterial and antifungal properties of the Propylamylatin formula, antiviral activity had yet to be noted [
5]; that is, until the 2013 to 2014 outbreak of porcine epidemic diarrhea (PED) caused by the porcine epidemic diseases virus (PEDv) in the United States. PED spread quickly throughout the pig population; one grower in Montana lost 850 piglets to PEDv infection within weeks of the disease reaching his locale. The Propylamylatin formula solution containing the propionic acid/isoamyl hexanoate electrolyte mixture was administered to tens of animals suffering with PED to assist in rehydration and electrolyte balance. Many of the animals that received the solution improved rapidly, showing evidence of decreased clinical signs after 5 h post-administration, and all animals completely recovered by 24 h (Strobel, unpublished record of Ecoplanet Environmental L.L.C). Interestingly, PEDv is a coronavirus and this fact led us to investigate the distinct possibility that the Propylamylatin formula may be helpful in other coronavirus infections [
6].
Thus, the immediate purpose of this report is to show, via gas and direct injection assays with virus suspensions, that the Propylamylatin formula is active against SARS-CoV-2.
2. Materials and Methods
2.1. Cells, Viruses, and the Propylamylatin Formula
Vero E6 cells were purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA) and cultured in Minimum Essential Medium (MEM; Corning Cellgro, Tewksbury, MA, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan City, UT, USA) and 1% penicillin-streptomycin solution (Hyclone). Cells were maintained at 37 °C, 5% CO2 and sub-cultured twice weekly to ensure sub-confluency.
The USA-WA1/2020 strain of SARS-CoV-2 was deposited by the Centers for Disease Control and Prevention and obtained through BEI Resources, NIAID, NIH (NR-52281). Viral stocks were propagated in Vero E6 cells for two days. Supernatants were collected, clarified by high-speed centrifugation, aliquoted and frozen at −80 °C. Viral stock titers were determined by plaque assay on Vero E6 cells. All work with SARS-CoV-2 was conducted under Biosafety Level 3 plus conditions at the Institute for Therapeutic Innovation (University of Florida, Orlando, FL, USA) using safety protocols approved by the University of Florida Institutional Biosafety Committee (BIO5525 approved 11 March 2020).
The Propylamylatin formula consists of a mixture of propionic acid and isoamyl hexanoates at a ratio of 7:2 (v/v) or with a molar ratio of 10.2:1. The isoamyl hexanoates are present in the mixture of esters at a ratio of 99:1 of the 3 to the 2 isomer. These compounds were obtained from Eccelentia International Co. (Fairfield, NJ, USA). Other chemicals were obtained from Sigma Aldrich (St. Louis, MO, USA).
2.2. The Propylamylatin Formula Gaseous and Direct Injection Evaluations against SARS-CoV-2
For vapor (gas phase) evaluations of the Propylamylatin formula, SARS-CoV-2 was diluted to a concentration equivalent to 10
5 plaque forming units (PFU)/mL in MEM. Aliquots containing 1 mL of the virus suspension (10
5 PFU) were placed into plastic caps and secured to the surface of 100 mm × 15 mm polystyrene petri dishes with double sided tape directly in the middle of the petri dishes. Small plastic caps (the lid from a standard microcentrifuge tube) containing 100 µls of the Propylamylatin formula were placed 2.5 cm away from the virus-containing caps. The top of the petri dish was sealed with parafilm and plates were incubated at both room temperature (~18 °C) and 37 °C for various lengths of time (
Figure 1). Since the removal of the top cover of the petri dish immediately destroys the gas atmosphere of that dish, one petri dish for each experimental arm was harvested at each time point. Time-matched control plates were included in the experimental design and set up exactly as described above with the exception that 100 µls of PBS was placed inside the white small plastic cap. At the time of sampling, viral supernatant was transferred from the plastic cap into a microfuge tube and frozen at −80 °C until the end of the study. The viral burden was quantified by plaque assay on Vero E6 cells for all samples simultaneously. Each experimental arm was assayed in duplicate to determine between sample variability and at least two independent assays were conducted on different days to address between-day variability.
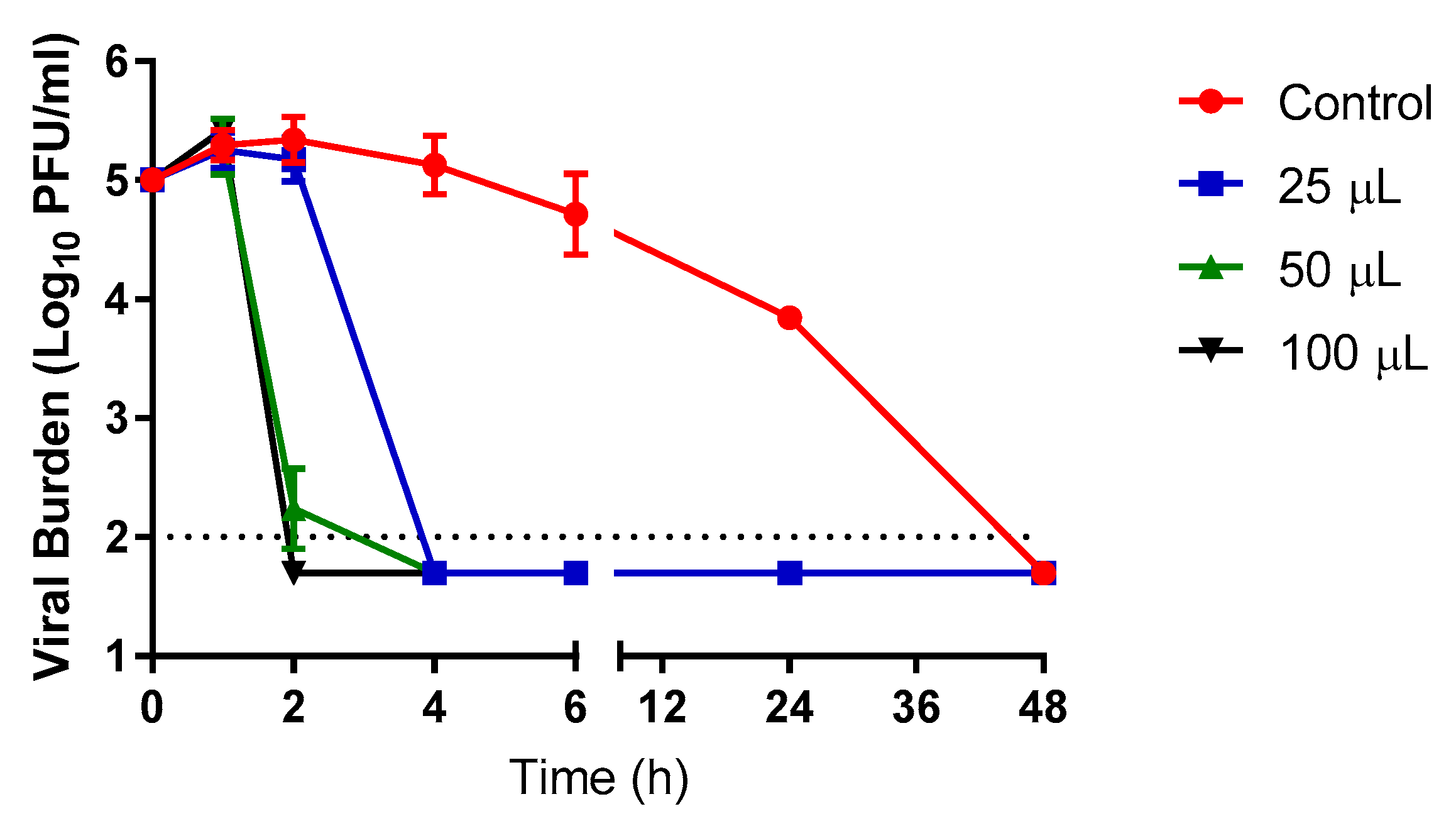
Additional vapor (gas phase) evaluations were conducted with various volumes of the Propylamylatin formula against SARS-CoV-2 at 37 °C. Studies were conducted as described above and shown in
Figure 1, but for these experiments different volumes ranging from 25 µls to 100 µls of the Propylamylatin formula were placed in the white small plastic cap. Time- and volume-matched PBS controls were included in this experiment. All experimental arms were sampled in duplicate and processed as described above. The viral burden was determined by plaque assay on Vero E6 cells and two independent studies were conducted on two different days.
The ability of the Propylamylatin formula to inactivate SARS-CoV-2 when directly injected into a viral suspension was also evaluated. For these studies, various volumes of the Propylamylatin formula were directly inoculated into 1 mL aliquots of SARS-CoV-2 diluted to a concentration of 105 PFU/mL in MEM and incubated at 37 °C. At various times post-inoculation, aliquots from each experimental arm were harvested, in duplicate, and frozen at −80 °C until the end of the study. The viral burden was quantified by plaque assay on Vero E6 cells for all samples simultaneously. At least two independent studies were conducted on different days to address between-day variability.
The anti-SARS-CoV-2 activity of the individual components making up the Propylamylatin formula (i.e., propionic acid alone and esters alone) was evaluated in the amounts in which they are present in the formula at the 7:2 (v/v) ratio, using the direct injection methods described above. Isobutyric acid (the original acid produced by M. albus) was substituted for the propionic acid in the mixture and also assessed via direct injection.
2.3. SARS-CoV-2 Plaque Assay
Vero E6 cells were seeded into six-well plates and incubated until confluency at 37 °C, 5% CO2. Viral samples were thawed on ice, serially diluted 10-fold in MEM, and 100 µls of each dilution was inoculated onto Vero E6 monolayers. Plates were incubated for 1 h to allow for viral attachment and were shaken every 15 min to maintain an even distribution of the viral inoculum on the cell monolayer. Monolayers were then overlaid with MEM containing a final concentration of 0.5% agar (w/v) and 5% FBS (v/v). Two days later, a secondary overlay containing MEM with a final concentration of 0.5% agar, 1% FBS, and 0.007% neutral red solution was added to each well of the six-well plate. Plaques were counted on the following day (72 h post-infection) with the naked eye.
2.4. Gas Chromatography/Mass Spectroscopy (GC/MS) and Gas Chromatography/Flame Ionization Detection (GC/FID) Plus Colorimetric Titration of the Virus Wells
We estimated the amount of dissolved propionic acid and isoamyl hexanoates in MEM when the Propylamylatin formula was administered via gas phase, and the resulting concentrations served as a guide for the design of the direct injection experiments. For these assays, petri dishes were set up exactly as described above for the Propylamylatin formula gas phase evaluations with the exception that the virus was not included in these studies. Propionic acid was quantified using two different methods. The first method was via titration of the MEM (which contains phenol red, a pH indicator) with a 1 M NaOH solution. The second method was by GC/FID in which samples, taken from the wells, were diluted 20-fold, separated by GC, and the amounts measured by FID. The samples were injected into a Hewlett Packard 6890 gas chromatograph containing a 30 m × 0.25 mm inner diameter ZB Wax capillary column with a film thickness of 0.50 mm. A thermal program of 30 °C for 2 min followed by an increase to 220 °C at 5 °C min−1 was applied. Ultrahigh-purity helium gas was used as the carrier gas and the initial column head pressure was 50 KPa. The quantification for both methods was conducted in triplicate.
The presence of the esters was determined both qualitatively and quantitatively by GC/MS using a Hewlett Packard 6890 gas chromatograph. The samples were diluted two-fold and the initial identification of the esters found in the analysis was made via library comparison using the National Institute of Standards and Technology (NIST) database. For GC/MS analyses, authentic samples of isoamyl hexanoates were used to generate standard curves to interpolate the concentrations of each compound in the media samples.
2.5. pH Determination of Virus Diluent Media
Various volumes of the Propylamylatin formula were added to 1 mL of MEM. The pH of the media was measured using MColorpHast™ pH test strips and indicator papers (MilliporeSigma, Burlington, MA, USA) at various times ranging from 15 min to 24 h post-Propylamylatin-formula addition.
2.6. Statistical Analyses
The volume of the Propylamylatin formula that reduces viral burden by 90% (EC90) was determined on individual time points as well as over the entire duration of the experiment. For analyses at individual time points, an inhibitory Sigmoid-Emax model (Hill model) was fit to the viral burden data from each experimental arm generated at that time point. Analyses of the entire data set was determined by first calculating the area under the viral burden time curve (AUCVB-Time) for each experimental condition. An inhibitory sigmoid-Emax model (Hill model) was then fit to the AUCVB-time values. All inhibitory sigmoid-Emax models were fit to the data using GraphPad Prism software (version 7.02).
4. Discussion
The COVID-19 pandemic continues to threaten global public health, spreading readily from person to person and resulting in significant illness and an unacceptably high number of deaths. Although numerous preclinical and clinical investigations have been conducted to identify effective drug candidates for the treatment and prevention of COVID-19, there are currently no antiviral agents approved for this serious disease. Natural products derived from plant, fungi, and marine sources have been, and are still, frequently used to treat viral infections, particularly those that cause respiratory illnesses such as rhinovirus and influenza virus [
2,
5,
7,
8,
9,
10,
11]. Natural products are attractive therapies for viral as well as other microbial infections because they are often accessible in nature (making them low in cost), amenable to mass production, and have generally good safety profiles [
9]. Moreover, the lack of available antiviral drugs for many ubiquitous viral diseases makes natural-based remedies a logical source of treatment for these infections, especially in developing countries [
12,
13]. For these studies, we aimed to evaluate the Propylamylatin formula against SARS-CoV-2, in an attempt to identify a potentially effective treatment strategy to combat COVID-19.
Because the antimicrobial activity of the natural fungus-derived VOCs was originally observed in the gas phase, we first evaluated the Propylamylatin formula in the gaseous state against SARS-CoV-2 at room temperature (18 °C) and body temperature (37 °C). This initial study was conducted in order to determine if the Propylamylatin formula might possess any antiviral activity and if so, at what concentrations. Ultimately, this information would serve as a guide for more comprehensive and better-controlled studies using a direct injection method of the Propylamylatin formula into viral suspension. As previously described by others, infectious SARS-CoV-2 virus is naturally more stable at cooler temperatures with the viral burden persisting longer at room temperature compared to body temperature [
14,
15]. The Propylamylatin formula vapors were effective against SARS-CoV-2 at both temperatures, resulting in undetectable levels of virus after exposure. However, viral inactivation was substantially faster at higher temperatures. This is likely due to the fact that the Propylamylatin formula volatizes more rapidly at higher temperatures. The impressive ability of the formula vapors to inactivate high levels (~10
5 PFU/mL) of SARS-CoV-2 at room temperature has important implications, as this agent may serve as a way to inactivate the virus on different surfaces including instruments and personal protective equipment. Studies evaluating the disinfecting potential of the Propylamylatin formula vapors against SARS-CoV-2 on surfaces made of plastic, glass, metal, etc. are currently ongoing.
Measuring the levels of dissolved propionic acid and isoamyl hexanoates in the tissue culture medium post-exposure to the vapors volatizing from the Propylamylatin formula at 37 °C revealed that propionic acid was readily detected but the esters (isoamyl hexanoates) were only present (at very low levels) when high starting volumes of compound were employed. These findings indicate that the majority of anti-SARS-CoV-2 activity was likely due to the propionic acid under this route of administration.
Per our hypothesis, our initial interest in the Propylamylatin formula was as a potential antiviral strategy against the virus causing COVID-19; thus, we conducted direct injection experiments where the Propylamylatin formula was directly added to a SARS-CoV-2 suspension at 37 °C to simulate body temperature. The addition of at least 10 µls of the Propylamylatin formula into 1 mL suspension of 10
5 PFU of SARS-CoV-2 completely inactivated SARS-CoV-2 to levels below the limit of detection after only 15 min of exposure. Assessing the individual components of the Propylamylatin formula revealed that indeed, as identified from the gas phase experiments, the propionic acid provided the majority of antiviral effect compared to the isoamyl hexanoates. However, the degree of inhibition was markedly greater when the propionic acid and the isoamyl hexanoates were combined, indicating synergy. These findings for antiviral activity are in concordance with what has been reported for the antimicrobial activity of the Propylamylatin formula against bacteria and fungi [
2,
3,
4,
5].
Isobutyric acid is the small weight molecular acid identified in the original VOCs made by the fungus. Isobutyric acid in combination with the isoamyl hexanoates was slightly more potent overall than the Propylamylatin formula (which contains propionic acid with the isoamyl hexanoates) yielding EC90 values of 2.58 µls and 4.28 µls, respectively. This difference in EC90 is mainly attributed to the 5 µL volume of the isobutyric mixture which completely inhibited SARS-CoV-2 after 15 min of exposure, yielding inhibitory kinetics that were identical to the 10 µL and 20 µls volumes. A 5 µL volume of the Propylamylatin formula provided complete inhibition after 60 min of exposure. These differences in antiviral activity are relatively negligible. Moreover, there are several benefits to using propionic acid over isobutyric acid, the most obvious being the unattractive odor of isobutyric acid, which smells like rancid butter. Propionic acid also has an unpleasant odor, but it is less pungent than that of isobutyric acid.
The degree and rapidity of viral inactivation via the Propylamylatin formula is impressive. It is hypothesized that the agent may inactivate the virus via interaction with the receptor binding domain (RBD) of the SARS-CoV-2 spike protein, which plays a critical role in viral attachment to the cellular angiotensin-converting enzyme 2 (ACE2) receptor [
16]. A polybasic cleavage site that is 10 nm from the RBD has been identified on the spike protein and shown to enhance the binding affinity of RBD to ACE2 [
17]. Blocking this positively charged site (arginine) with negatively charged anions in the presence of a strong lipophilic agent should effectively neutralize this critical site and render it ineffective. This phenomenon was shown to be true by Qiao et al., as the addition of a synthetic peptide GluGluLeuGlu to SARS-CoV-2 reduced the binding affinity of RBD to ACE2 by 34% [
17]. The reduction of binding strength would feasibly reduce the infectivity of the virus. Thus, this may be one mechanism by which the Propylamylatin formula inactivates SARS-CoV-2. A second mechanism by which this agent may act is through its ability to create an acidic environment. The pH was inversely proportional to antiviral activity, with lower pH values resulting in a more rapid decline in viral burden. Others have reported that the infectivity of coronaviruses including SARS-CoV (causative agent of the 2002 to 2003 SARS outbreak) are very sensitive to pH extremes, as lower pH values were able to completely inactivate SARS-CoV [
18]. Thus, the Propylamylatin formula may rapidly inactivate infectious SARS-CoV-2 by blocking the polybasic cleavage site on the spike protein and/or creating an acidic environment that renders the virus non-infectious.
Our studies illustrate the therapeutic potential of the Propylamylatin formula for the treatment of COVID-19. As shown above, a volume of only 10 µls was required to completely inactivate infectious SARS-CoV-2 after only 15 min of contact time. This amount of formula is equivalent to 1% of the total volume of liquid in the suspension (10 µls of the Propylamylatin formula into 1000 µls of virus suspension) and represents the exact amount of the Propylamylatin formula that was present in the electrolyte solution originally administered to piglets suffering with PEDv. These findings warrant further investigations to determine if the level of the Propylamylatin formula required to inactivate SARS-CoV-2 is physiologically achievable in humans and whether it will have a direct application to treat human COVID-19.
Gastrointestinal symptoms have been reported in patients with COVID-19 [
19,
20] and SARS-CoV-2 has been shown to infect human gut enterocytes [
21]. Therefore, a 1% solution of Propylamylatin formula may be sufficient to inhibit viral replication in the digestive system, thereby alleviating gastrointestinal symptoms. Respiratory symptoms may also be relieved with the formula. The Propylamylatin formula could possibly be applied as a topical agent in the nasal cavities to inactivate infectious virus in the nose, which is the initial site of infection. It has been proposed that aspiration from the nasal cavity into the oral cavity may be a route for the virus to transverse from the upper respiratory system into the lower respiratory system, resulting in more severe disease outcomes [
22,
23]. Thus, a 1% solution of the Propylamylatin formula as a mouthwash may inhibit any virus in the oral cavity from reaching the lungs and preventing pneumonia as well as other more severe disease outcomes. Finally, Propylamylatin formula when administered via an atomizer, nebulizer, or intubation directly into lung tissue may be helpful in reducing viral load in these tissues. Additional studies are required to address these hypotheses.
There are some weaknesses associated with our study. First, our experiments were conducted with a single strain of SARS-CoV-2. Studies will be repeated with new viral variants, including the mutated United Kingdom and South African strains, to ensure that the Propylamylatin formula is effective against a variety of SARS-CoV-2 strains. Secondly, we plan to evaluate toxicities associated with the 1% solution in the lung tissue using small rodent models.
In conclusion, we have demonstrated that the Propylamylatin formula holds promise as a therapeutic strategy to be further investigated for patients afflicted with COVID-19, a disease for which there are no approved antiviral therapies. The potent direct inhibition of infectious SARS-CoV-2 suggest that further preclinical (in vitro and in vivo) studies are warranted for this virus and other coronaviruses.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}