
Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethics Statement
2.3. HIV-1 Nef Sequencing
2.4. Plasmids
2.5. Cell Lines
2.6. Virus Production
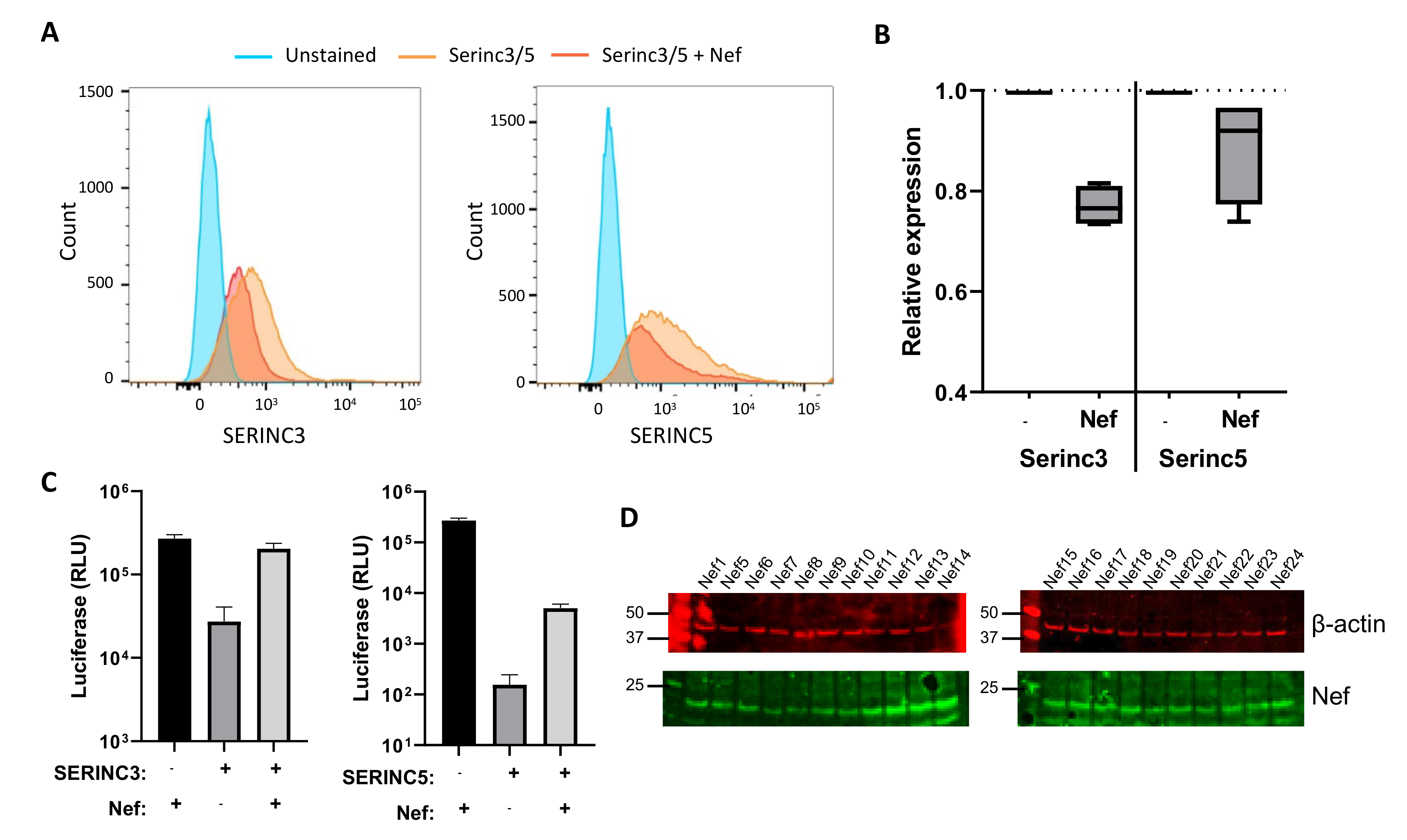
2.7. Flow Cytometry
2.8. Western Blot Analysis
2.9. Viral Infection
2.10. Statistical Analysis
3. Results
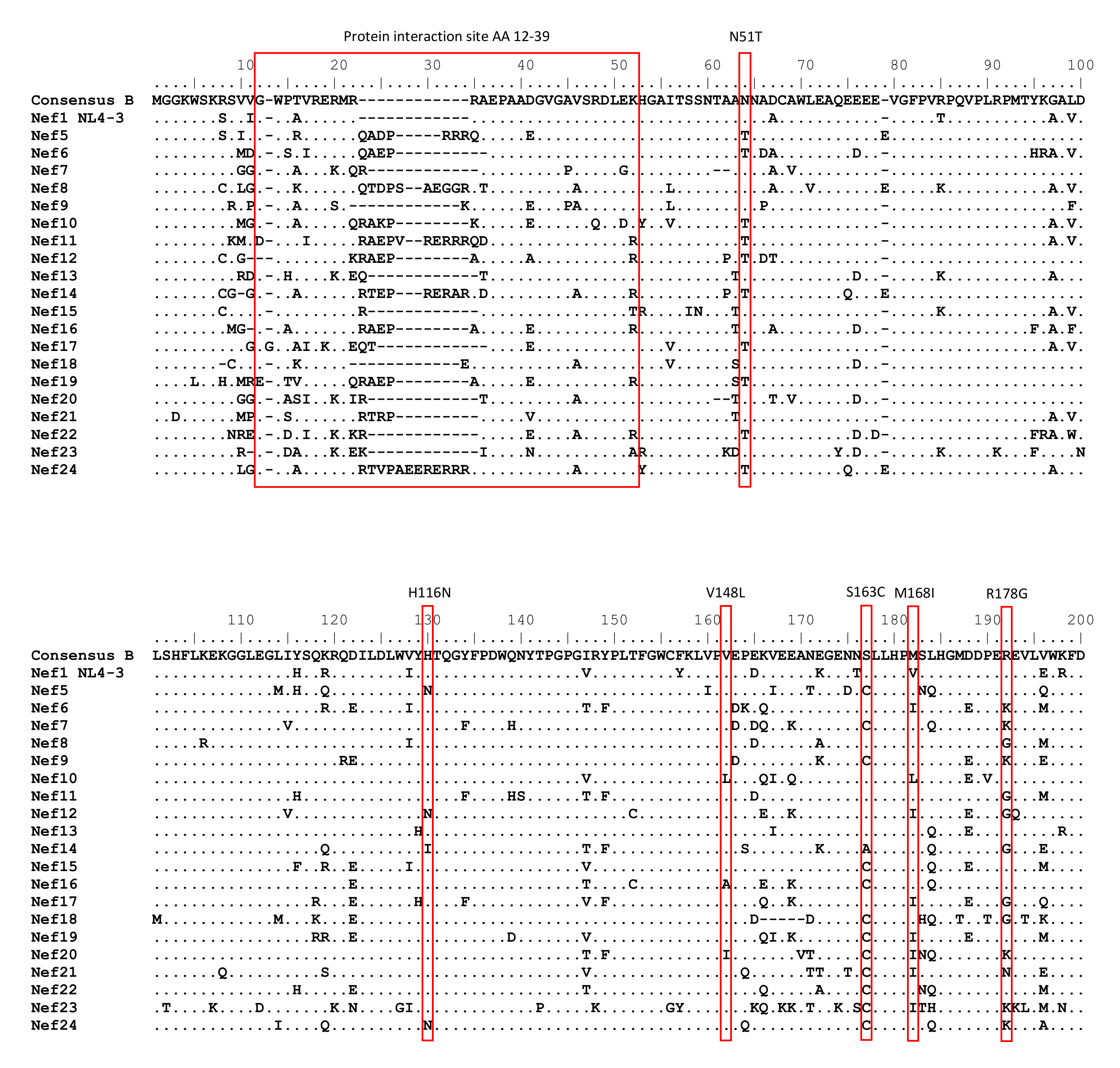
3.1. Naturally Occurring Variation in Nef
3.2. Effect of Naturally Occurring Amino Acid Variations on Nef Activity to Counteract SERINC3 and SERINC5
3.3. Association between Naturally Occurring Mutations in Nef and the Ability to Counteract SERINC3 and SERINC5
3.4. Effect of Naturally Occurring Mutations in Nef on Disease Progression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Landi, A.; Iannucci, V.; Nuffel, A.V.; Meuwissen, P.; Verhasselt, B. One protein to rule them all: Modulation of cell surface receptors and molecules by HIV Nef. Curr. HIV Res. 2011, 9, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Alsahafi, N.; Ding, S.; Richard, J.; Markle, T.; Brassard, N.; Walker, B.; Lewis, G.K.; Kaufmann, D.E.; Brockman, M.A.; Finzi, A. Nef proteins from HIV-1 elite controllers are inefficient at preventing antibody-dependent cellular cytotoxicity. J. Virol. 2015, 90, 2993–3002. [Google Scholar] [CrossRef] [Green Version]
- Collins, K.L.; Chen, B.K.; Kalams, S.A.; Walker, B.D.; Baltimore, D. HIV-1 Nef protein protects infected primary cells against killing by cytotoxic T lymphocytes. Nature 1998, 391, 397–401. [Google Scholar] [CrossRef]
- Le Gall, S.; Erdtmann, L.; Benichou, S.; Berlioz-Torrent, C.; Liu, L.; Benarous, R.; Heard, J.M.; Schwartz, O. Nef interacts with the mu subunit of clathrin adaptor complexes and reveals a cryptic sorting signal in MHC I molecules. Immunity 1998, 8, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Veillette, M.; Coutu, M.; Richard, J.; Batraville, L.A.; Dagher, O.; Bernard, N.; Tremblay, C.; Kaufmann, D.E.; Roger, M.; Finzi, A. The HIV-1 GP120 CD4-bound conformation is preferentially targeted by antibody-dependent cellular cytotoxicity-mediating antibodies in sera from HIV-1-infected individuals. J. Virol. 2015, 89, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, L.; Du, Z.; Rosenzweig, M.; Jung, J.U.; Desrosiers, R.C. A role for natural simian immunodeficiency virus and human immunodeficiency virus type 1 nef alleles in lymphocyte activation. J. Virol. 1997, 71, 6094–6099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baur, A.S.; Sass, G.; Laffert, B.; Willbold, D.; Cheng-Mayer, C.; Peterlin, B.M. The N-terminus of Nef from HIV-1/SIV associates with a protein complex containing Lck and a serine kinase. Immunity 1997, 6, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Stolp, B.; Reichman-Fried, M.; Abraham, L.; Pan, X.; Giese, S.I.; Hannemann, S.; Goulimari, P.; Raz, E.; Grosse, R.; Fackler, O.T. HIV-1 Nef interferes with host cell motility by deregulation of Cofilin. Cell Host Microbe 2009, 6, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Rosa, A.; Chande, A.; Ziglio, S.; De Sanctis, V.; Bertorelli, R.; Goh, S.L.; McCauley, S.M.; Nowosielska, A.; Antonarakis, S.E.; Luban, J.; et al. HIV-1 Nef promotes infection by excluding SERINC5 from virion incorporation. Nature 2015, 526, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Usami, Y.; Wu, Y.; Gottlinger, H.G. SERINC3 and SERINC5 restrict HIV-1 infectivity and are counteracted by Nef. Nature 2015, 526, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Sood, C.; Marin, M.; Chande, A.; Pizzato, M.; Melikyan, G.B. SERINC5 protein inhibits HIV-1 fusion pore formation by promoting functional inactivation of envelope glycoproteins. J. Biol. Chem. 2017, 292, 6014–6026. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Xiong, R.; Zhou, T.; Su, P.; Zhang, X.; Qiu, X.; Li, H.; Li, S.; Yu, C.; Wang, B.; et al. HIV-1 Nef Antagonizes SERINC5 Restriction by downregulation of SERINC5 via the endosome/lysosome system. J. Virol. 2018, 92, e00196. [Google Scholar] [CrossRef] [Green Version]
- Foster, J.L.; Denial, S.J.; Temple, B.R.; Garcia, J.V. Mechanisms of HIV-1 Nef function and intracellular signaling. J. Neuroimmune Pharmacol. 2011, 6, 230–246. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.W.; Alsahafi, N.; Kuang, X.T.; Swann, S.A.; Toyoda, M.; Gottlinger, H.; Walker, B.D.; Ueno, T.; Finzi, A.; Brumme, Z.L.; et al. Natural HIV-1 nef polymorphisms impair SERINC5 downregulation activity. Cell Rep. 2019, 29, 1449–1457.e45. [Google Scholar] [CrossRef] [PubMed]
- Sauter, D.; Hotter, D.; Van Driessche, B.; Sturzel, C.M.; Kluge, S.F.; Wildum, S.; Yu, H.; Baumann, B.; Wirth, T.; Plantier, J.C.; et al. Differential regulation of NF-kappaB-mediated proviral and antiviral host gene expression by primate lentiviral Nef and Vpu proteins. Cell Rep. 2015, 10, 586–599. [Google Scholar] [CrossRef]
- Lindwasser, O.W.; Smith, W.J.; Chaudhuri, R.; Yang, P.; Hurley, J.H.; Bonifacino, J.S. A diacidic motif in human immunodeficiency virus type 1 Nef is a novel determinant of binding to AP-2. J. Virol. 2008, 82, 1166–1174. [Google Scholar] [CrossRef] [Green Version]
- Kootstra, N.A.; Navis, M.; Beugeling, C.; van Dort, K.A.; Schuitemaker, H. The presence of the Trim5alpha escape mutation H87Q in the capsid of late stage HIV-1 variants is preceded by a prolonged asymptomatic infection phase. AIDS 2007, 21, 2015–2023. [Google Scholar] [CrossRef]
- Schuitemaker, H.; Koot, M.; Kootstra, N.A.; Dercksen, M.W.; de Goede, R.E.; van Steenwijk, R.P.; Lange, J.M.; Schattenkerk, J.K.; Miedema, F.; Tersmette, M. Biological phenotype of human immunodeficiency virus type 1 clones at different stages of infection: Progression of disease is associated with a shift from monocytotropic to T-cell-tropic virus population. J. Virol. 1992, 66, 1354–1360. [Google Scholar] [CrossRef] [Green Version]
- Bjorndal, A.; Deng, H.; Jansson, M.; Fiore, J.R.; Colognesi, C.; Karlsson, A.; Albert, J.; Scarlatti, G.; Littman, D.R.; Fenyo, E.M. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J. Virol. 1997, 71, 7478–7487. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.K.; Saksela, K.; Andino, R.; Baltimore, D. Distinct modes of human immunodeficiency virus type 1 proviral latency revealed by superinfection of nonproductively infected cell lines with recombinant luciferase-encoding viruses. J. Virol. 1994, 68, 654–660. [Google Scholar] [CrossRef] [Green Version]
- Connor, R.I.; Chen, B.K.; Choe, S.; Landau, N.R. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology 1995, 206, 935–944. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Choe, S.; Walker, R.; Di Marzio, P.; Morgan, D.O.; Landau, N.R. Human immunodeficiency virus type 1 viral protein R (VPR) arrests cells in the G2 phase of the cell cycle by inhibiting P34cdc2 activity. J. Virol. 1995, 69, 6705–6711. [Google Scholar] [CrossRef] [Green Version]
- Tersmette, M.; Winkel, I.N.; Groenink, M.; Gruters, R.A.; Spence, R.P.; Saman, E.; Van Der Groen, G.; Miedema, F.; Huisman, J.G. Detection and subtyping of HIV-1 isolates with a panel of characterized monoclonal antibodies to HIV p24gag. Virology 1989, 171, 149–155. [Google Scholar] [CrossRef]
- Ananth, S.; Morath, K.; Trautz, B.; Tibroni, N.; Shytaj, I.L.; Obermaier, B.; Stolp, B.; Lusic, M.; Fackler, O.T. Multifunctional roles of the N-terminal region of HIV-1SF2Nef are mediated by three independent protein interaction sites. J. Virol. 2019, 94, e01398. [Google Scholar] [CrossRef]
- Sudderuddin, H.; Kinloch, N.N.; Jin, S.W.; Miller, R.L.; Jones, B.R.; Brumme, C.J.; Joy, J.B.; Brockman, M.A.; Brumme, Z.L. Longitudinal within-host evolution of HIV Nef-mediated CD4, HLA and SERINC5 downregulation activity: A case study. Retrovirology 2020, 17, 3. [Google Scholar] [CrossRef]
- Jin, S.W.; Mwimanzi, F.M.; Mann, J.K.; Bwana, M.B.; Lee, G.Q.; Brumme, C.J.; Hunt, P.W.; Martin, J.N.; Bangsberg, D.R.; Ndung’u, T.; et al. Variation in HIV-1 Nef function within and among viral subtypes reveals genetically separable antagonism of SERINC3 and SERINC5. PLoS Pathog. 2020, 16, e1008813. [Google Scholar] [CrossRef]
- Homann, S.; Tibroni, N.; Baumann, I.; Sertel, S.; Keppler, O.T.; Fackler, O.T. Determinants in HIV-1 Nef for enhancement of virus replication and depletion of CD4+ T lymphocytes in human lymphoid tissue ex vivo. Retrovirology 2009, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Schierer, S.; Blume, K.; Dindorf, J.; Wittki, S.; Xiang, W.; Ostalecki, C.; Koliha, N.; Wild, S.; Schuler, G.; et al. HIV-Nef and ADAM17-containing plasma extracellular vesicles induce and correlate with immune pathogenesis in chronic HIV infection. EBioMedicine 2016, 6, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Ostalecki, C.; Wittki, S.; Lee, J.H.; Geist, M.M.; Tibroni, N.; Harrer, T.; Schuler, G.; Fackler, O.T.; Baur, A.S. HIV Nef- and Notch1-dependent endocytosis of ADAM17 induces vesicular TNF secretion in chronic HIV infection. EBioMedicine 2016, 13, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Fackler, O.T.; Moris, A.; Tibroni, N.; Giese, S.I.; Glass, B.; Schwartz, O.; Krausslich, H.G. Functional characterization of HIV-1 Nef mutants in the context of viral infection. Virology 2006, 351, 322–339. [Google Scholar] [CrossRef]
- Trautz, B.; Pierini, V.; Wombacher, R.; Stolp, B.; Chase, A.J.; Pizzato, M.; Fackler, O.T. The antagonism of HIV-1 Nef to SERINC5 particle infectivity restriction involves the counteraction of virion-associated pools of the restriction factor. J. Virol. 2016, 90, 10915–10927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.A.; Huang, M.B.; Campbell, P.E.; Roth, W.W.; Campbell, T.; Khan, M.; Newman, G.; Villinger, F.; Powell, M.D.; Bond, V.C. Genetic characterization of HIV type 1 Nef-induced vesicle secretion. AIDS Res. Hum. Retroviruses 2010, 26, 173–192. [Google Scholar] [CrossRef]
- Bentham, M.; Mazaleyrat, S.; Harris, M. Role of myristoylation and N-terminal basic residues in membrane association of the human immunodeficiency virus type 1 Nef protein. J. Gen. Virol. 2006, 87, 563–571. [Google Scholar] [CrossRef]
- Welker, R.; Harris, M.; Cardel, B.; Krausslich, H.G. Virion incorporation of human immunodeficiency virus type 1 Nef is mediated by a bipartite membrane-targeting signal: Analysis of its role in enhancement of viral infectivity. J. Virol. 1998, 72, 8833–8840. [Google Scholar] [CrossRef]
- Cruz, N.V.; Amorim, R.; Oliveira, F.E.; Speranza, F.A.; Costa, L.J. Mutations in the nef and vif genes associated with progression to AIDS in elite controller and slow-progressor patients. J. Med. Virol. 2013, 85, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Burtey, A.; Rappoport, J.Z.; Bouchet, J.; Basmaciogullari, S.; Guatelli, J.; Simon, S.M.; Benichou, S.; Benmerah, A. Dynamic interaction of HIV-1 Nef with the clathrin-mediated endocytic pathway at the plasma membrane. Traffic 2007, 8, 61–76. [Google Scholar] [CrossRef]
- Johannes, L.; Pezo, V.; Mallard, F.; Tenza, D.; Wiltz, A.; Saint-Pol, A.; Helft, J.; Antony, C.; Benaroch, P. Effects of HIV-1 Nef on retrograde transport from the plasma membrane to the endoplasmic reticulum. Traffic 2003, 4, 323–332. [Google Scholar] [CrossRef]
- Pizzato, M.; Helander, A.; Popova, E.; Calistri, A.; Zamborlini, A.; Palu, G.; Gottlinger, H.G. Dynamin 2 is required for the enhancement of HIV-1 infectivity by Nef. Proc. Natl. Acad. Sci. USA 2007, 104, 6812–6817. [Google Scholar] [CrossRef] [Green Version]
- Noviello, C.M.; Benichou, S.; Guatelli, J.C. Cooperative binding of the class I major histocompatibility complex cytoplasmic domain and human immunodeficiency virus type 1 Nef to the endosomal AP-1 complex via its mu subunit. J. Virol. 2008, 82, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Roeth, J.F.; Williams, M.; Kasper, M.R.; Filzen, T.M.; Collins, K.L. HIV-1 Nef disrupts MHC-I trafficking by recruiting AP-1 to the MHC-I cytoplasmic tail. J. Cell Biol. 2004, 167, 903–913. [Google Scholar] [CrossRef]
- Singh, R.K.; Lau, D.; Noviello, C.M.; Ghosh, P.; Guatelli, J.C. An MHC-I cytoplasmic domain/HIV-1 Nef fusion protein binds directly to the mu subunit of the AP-1 endosomal coat complex. PLoS ONE 2009, 4, e8364. [Google Scholar] [CrossRef] [Green Version]
- Aiken, C.; Krause, L.; Chen, Y.L.; Trono, D. Mutational analysis of HIV-1 Nef: Identification of two mutants that are temperature-sensitive for CD4 downregulation. Virology 1996, 217, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Roeth, J.F.; Kasper, M.R.; Filzen, T.M.; Collins, K.L. Human immunodeficiency virus type 1 Nef domains required for disruption of major histocompatibility complex class I trafficking are also necessary for coprecipitation of Nef with HLA-A2. J. Virol. 2005, 79, 632–636. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.; Roeth, J.F.; Kasper, M.R.; Fleis, R.I.; Przybycin, C.G.; Collins, K.L. Direct binding of human immunodeficiency virus type 1 Nef to the major histocompatibility complex class I (MHC-I) cytoplasmic tail disrupts MHC-I trafficking. J. Virol. 2002, 76, 12173–12184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wonderlich, E.R.; Williams, M.; Collins, K.L. The tyrosine binding pocket in the adaptor protein 1 (AP-1) mu1 subunit is necessary for Nef to recruit AP-1 to the major histocompatibility complex class I cytoplasmic tail. J. Biol. Chem. 2008, 283, 3011–3022. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Rosales, T.; Rose, J.J.; Chowdhury, B.; Knutson, J.R.; Venkatesan, S. HIV-1 Nef binds a subpopulation of MHC-I throughout its trafficking itinerary and down-regulates MHC-I by perturbing both anterograde and retrograde trafficking. J. Biol. Chem. 2010, 285, 30884–30905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, J.H.; Bonifacino, J.S. Nef-arious goings-on at the Golgi. Nat. Struct. Mol. Biol. 2012, 19, 661–662. [Google Scholar] [CrossRef]
- Jia, X.; Singh, R.; Homann, S.; Yang, H.; Guatelli, J.; Xiong, Y. Structural basis of evasion of cellular adaptive immunity by HIV-1 Nef. Nat. Struct. Mol. Biol. 2012, 19, 701–706. [Google Scholar] [CrossRef]
- Schaefer, M.R.; Wonderlich, E.R.; Roeth, J.F.; Leonard, J.A.; Collins, K.L. HIV-1 Nef targets MHC-I and CD4 for degradation via a final common beta-COP-dependent pathway in T cells. PLoS Pathog. 2008, 4, e1000131. [Google Scholar] [CrossRef] [Green Version]
- Bresnahan, P.A.; Yonemoto, W.; Ferrell, S.; Williams-Herman, D.; Geleziunas, R.; Greene, W.C. A dileucine motif in HIV-1 Nef acts as an internalization signal for CD4 downregulation and binds the AP-1 clathrin adaptor. Curr. Biol. 1998, 8, 1235–1238. [Google Scholar] [CrossRef] [Green Version]
- Craig, H.M.; Pandori, M.W.; Guatelli, J.C. Interaction of HIV-1 Nef with the cellular dileucine-based sorting pathway is required for CD4 down-regulation and optimal viral infectivity. Proc. Natl. Acad. Sci. USA 1998, 95, 11229–11234. [Google Scholar] [CrossRef] [Green Version]
- Macchi, B.; Graziani, G.; Zhang, J.; Mastino, A. Emergence of double-positive CD4/CD8 cells from adult peripheral blood mononuclear cells infected with human T cell leukemia virus type I (HTLV-I). Cell Immunol. 1993, 149, 376–389. [Google Scholar] [CrossRef]
- Piguet, V.; Gu, F.; Foti, M.; Demaurex, N.; Gruenberg, J.; Carpentier, J.L.; Trono, D. Nef-induced CD4 degradation: A diacidic-based motif in Nef functions as a lysosomal targeting signal through the binding of beta-COP in endosomes. Cell 1999, 97, 63–73. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Mattera, R.; Lindwasser, O.W.; Robinson, M.S.; Bonifacino, J.S. A basic patch on alpha-adaptin is required for binding of human immunodeficiency virus type 1 Nef and cooperative assembly of a CD4-Nef-AP-2 complex. J. Virol. 2009, 83, 2518–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.J.; Cai, C.Y.; Zhang, X.; Zhang, H.T.; Hirst, J.A.; Burakoff, S.J. HIV Nef-mediated CD4 down-regulation is adaptor protein complex 2 dependent. J. Immunol. 2005, 175, 3157–3164. [Google Scholar] [CrossRef] [Green Version]
- Stove, V.; Van de Walle, I.; Naessens, E.; Coene, E.; Stove, C.; Plum, J.; Verhasselt, B. Human immunodeficiency virus Nef induces rapid internalization of the T-cell coreceptor CD8alphabeta. J. Virol. 2005, 79, 11422–11433. [Google Scholar] [CrossRef] [Green Version]
- Leonard, J.A.; Filzen, T.; Carter, C.C.; Schaefer, M.; Collins, K.L. HIV-1 Nef disrupts intracellular trafficking of major histocompatibility complex class I, CD4, CD8, and CD28 by distinct pathways that share common elements. J. Virol. 2011, 85, 6867–6881. [Google Scholar] [CrossRef] [Green Version]
- Swigut, T.; Shohdy, N.; Skowronski, J. Mechanism for down-regulation of CD28 by Nef. EMBO J. 2001, 20, 1593–1604. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.A.; da Silva, L.L. HIV-1 Nef: Taking control of protein trafficking. Traffic 2016, 17, 976–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, H.M.; Reddy, T.R.; Riggs, N.L.; Dao, P.P.; Guatelli, J.C. Interactions of HIV-1 nef with the mu subunits of adaptor protein complexes 1, 2, and 3: Role of the dileucine-based sorting motif. Virology 2000, 271, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, M.; DeTulleo, L.; Rapoport, I.; Skowronski, J.; Kirchhausen, T. A dileucine motif in HIV-1 Nef is essential for sorting into clathrin-coated pits and for downregulation of CD4. Curr. Biol. 1998, 8, 1239–1242. [Google Scholar] [CrossRef] [Green Version]
- Obermaier, B.; Ananth, S.; Tibroni, N.; Pierini, V.; Shytaj, I.L.; Diaz, R.S.; Lusic, M.; Fackler, O.T. Patient-derived HIV-1 Nef alleles reveal uncoupling of CD4 downregulation and SERINC5 antagonism functions of the viral pathogenesis factor. J. Acquir. Immune Defic. Syndr. 2020, 85, e23–e26. [Google Scholar] [CrossRef]
- Heigele, A.; Kmiec, D.; Regensburger, K.; Langer, S.; Peiffer, L.; Sturzel, C.M.; Sauter, D.; Peeters, M.; Pizzato, M.; Learn, G.H.; et al. The potency of Nef-mediated SERINC5 antagonism correlates with the prevalence of primate lentiviruses in the wild. Cell Host Microbe 2016, 20, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Mwimanzi, P.; Markle, T.J.; Martin, E.; Ogata, Y.; Kuang, X.T.; Tokunaga, M.; Mahiti, M.; Pereyra, F.; Miura, T.; Walker, B.D.; et al. Attenuation of multiple Nef functions in HIV-1 elite controllers. Retrovirology 2013, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Park, S.Y.; Bonifacino, J.S.; Hurley, J.H. How HIV-1 Nef hijacks the AP-2 clathrin adaptor to downregulate CD4. eLife 2014, 3, e01754. [Google Scholar] [CrossRef]
- Little, S.J.; Riggs, N.L.; Chowers, M.Y.; Fitch, N.J.; Richman, D.D.; Spina, C.A.; Guatelli, J.C. Cell surface CD4 downregulation and resistance to superinfection induced by a defective provirus of HIV-1. Virology 1994, 205, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ueno, T.; Nakahara, T.; Sasaki, K.; Ishimoto, A.; Sakai, H. Downregulation of CD4 is required for maintenance of viral infectivity of HIV-1. Virology 2003, 311, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Volsky, D.J.; Simm, M.; Shahabuddin, M.; Li, G.; Chao, W.; Potash, M.J. Interference to human immunodeficiency virus type 1 infection in the absence of downmodulation of the principal virus receptor, CD4. J. Virol. 1996, 70, 3823–3833. [Google Scholar] [CrossRef] [Green Version]
- Benson, R.E.; Sanfridson, A.; Ottinger, J.S.; Doyle, C.; Cullen, B.R. Downregulation of cell-surface CD4 expression by simian immunodeficiency virus Nef prevents viral super infection. J. Exp. Med. 1993, 177, 1561–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, M.; Meloche, S.; Gratton, S.; Wainberg, M.A.; Sekaly, R.P. Association of p56lck with the cytoplasmic domain of CD4 modulates HIV-1 expression. EMBO J. 1994, 13, 774–783. [Google Scholar] [CrossRef]
- Chun, T.W.; Nickle, D.C.; Justement, J.S.; Meyers, J.H.; Roby, G.; Hallahan, C.W.; Kottilil, S.; Moir, S.; Mican, J.M.; Mullins, J.I.; et al. Persistence of HIV in gut-associated lymphoid tissue despite long-term antiretroviral therapy. J. Infect. Dis. 2008, 197, 714–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klatt, N.R.; Chomont, N.; Douek, D.C.; Deeks, S.G. Immune activation and HIV persistence: Implications for curative approaches to HIV infection. Immunol. Rev. 2013, 254, 326–342. [Google Scholar] [CrossRef] [Green Version]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nef Variation | % of Patients (n) n = 123 | ||
|---|---|---|---|
| Protein interaction site (AA 12–39): | |||
| Extra length | 46.3% (57) | ||
| Change of charge | 55.3% (68) | ||
| Amino acid polymorphisms: | |||
| 8R | 52.0% (64) | 81F | 10.6% (13) |
| 9S | 63.4% (78) | H116N | 17.9% (22) |
| 11P | 5.7% (7) | 120F | 6.5% (8) |
| 12G | 82.1% (101) | V148L/V148X | 10.6% (13)/13.8% (17) |
| 14A/14P | 5.7% (7)/25.2% (31) | 157N | 85.4% (105) |
| 15A | 52.8% (65) | 158K | 11.4% (14) |
| 21K/21R | 11.4% (14)/64.2% (79) | 161N | 93.5% (115) |
| 28E | 35.8% (44) | S163C | 39.8% (49) |
| 43I | 82.1% (101) | M168I/M168X | 22.0% (27)/27.6% (34) |
| N51T | 61.8% (76) | R178G | 17.9% (22) |
| 54D | 78.0% (96) | 182E | 19.5% (24) |
| 63E | 79.7% (98) | 188S | 6.5% (8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruize, Z.; van Nuenen, A.C.; van Wijk, S.W.; Girigorie, A.F.; van Dort, K.A.; Booiman, T.; Kootstra, N.A. Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction. Viruses 2021, 13, 423. https://doi.org/10.3390/v13030423
Kruize Z, van Nuenen AC, van Wijk SW, Girigorie AF, van Dort KA, Booiman T, Kootstra NA. Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction. Viruses. 2021; 13(3):423. https://doi.org/10.3390/v13030423
Chicago/Turabian StyleKruize, Zita, Ad C. van Nuenen, Stan W. van Wijk, Arginell F. Girigorie, Karel A. van Dort, Thijs Booiman, and Neeltje A. Kootstra. 2021. "Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction" Viruses 13, no. 3: 423. https://doi.org/10.3390/v13030423
APA StyleKruize, Z., van Nuenen, A. C., van Wijk, S. W., Girigorie, A. F., van Dort, K. A., Booiman, T., & Kootstra, N. A. (2021). Nef Obtained from Individuals with HIV-1 Vary in Their Ability to Antagonize SERINC3- and SERINC5-Mediated HIV-1 Restriction. Viruses, 13(3), 423. https://doi.org/10.3390/v13030423