
Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe?


 and
and 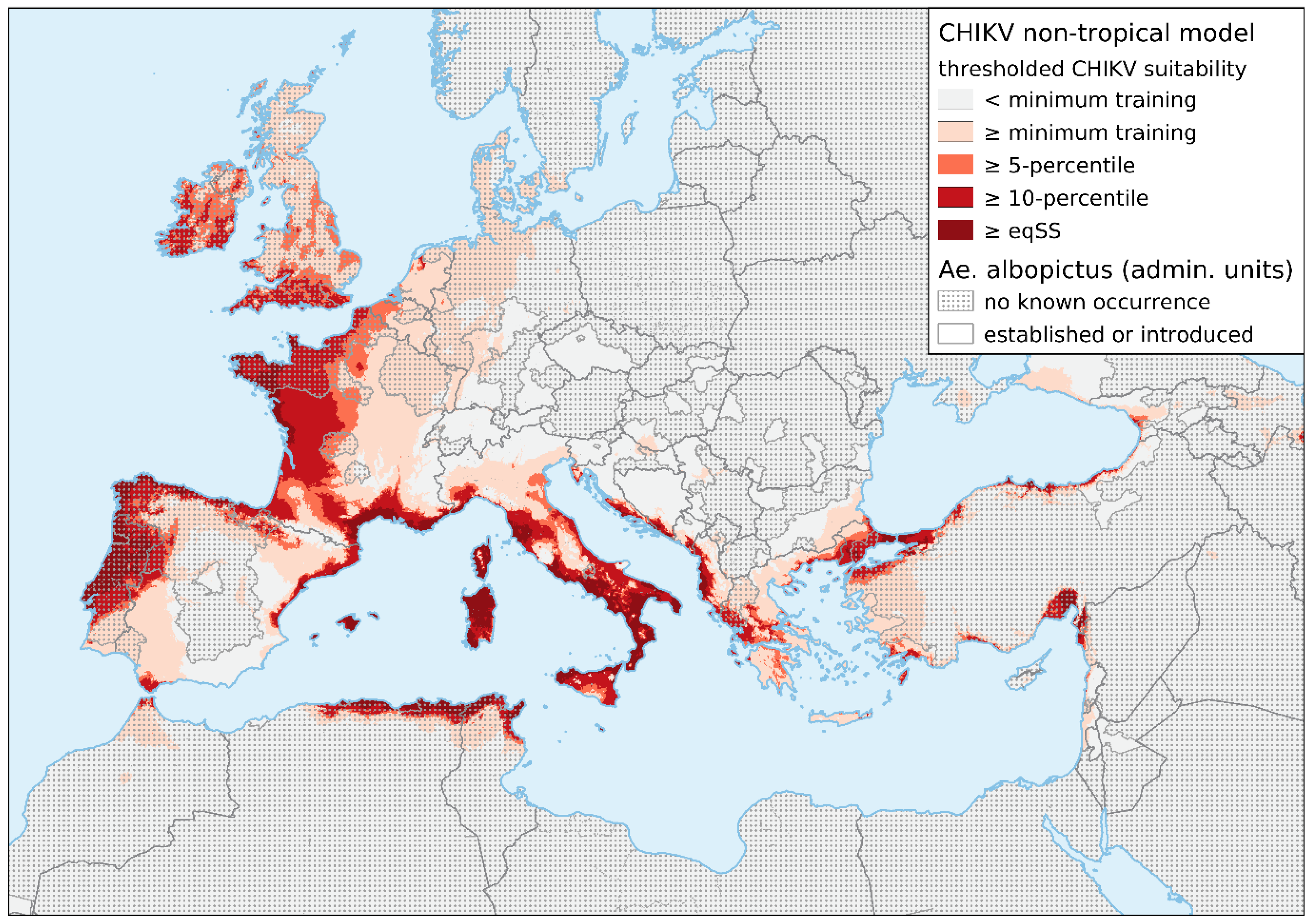{kind=link}
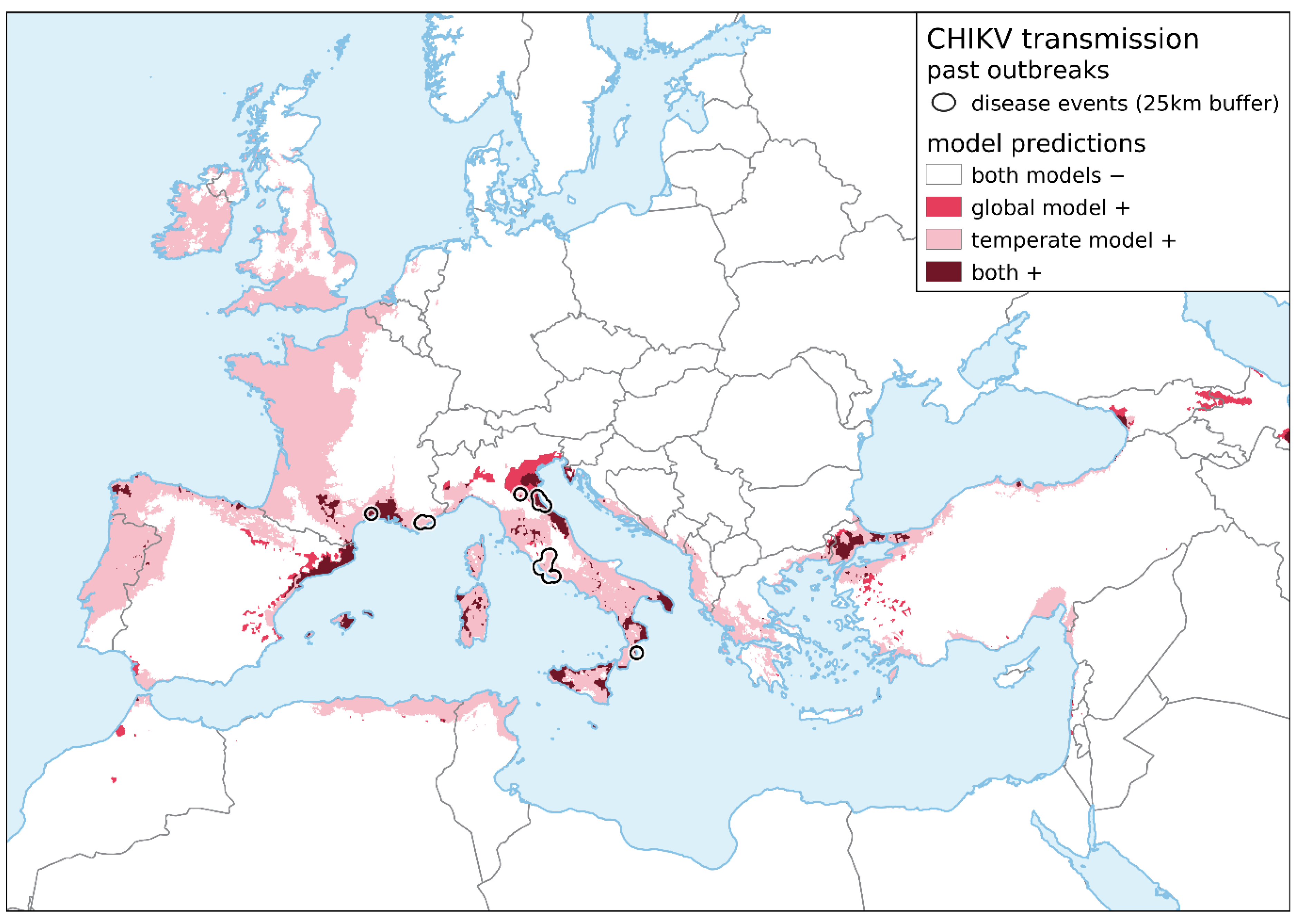{kind=link}
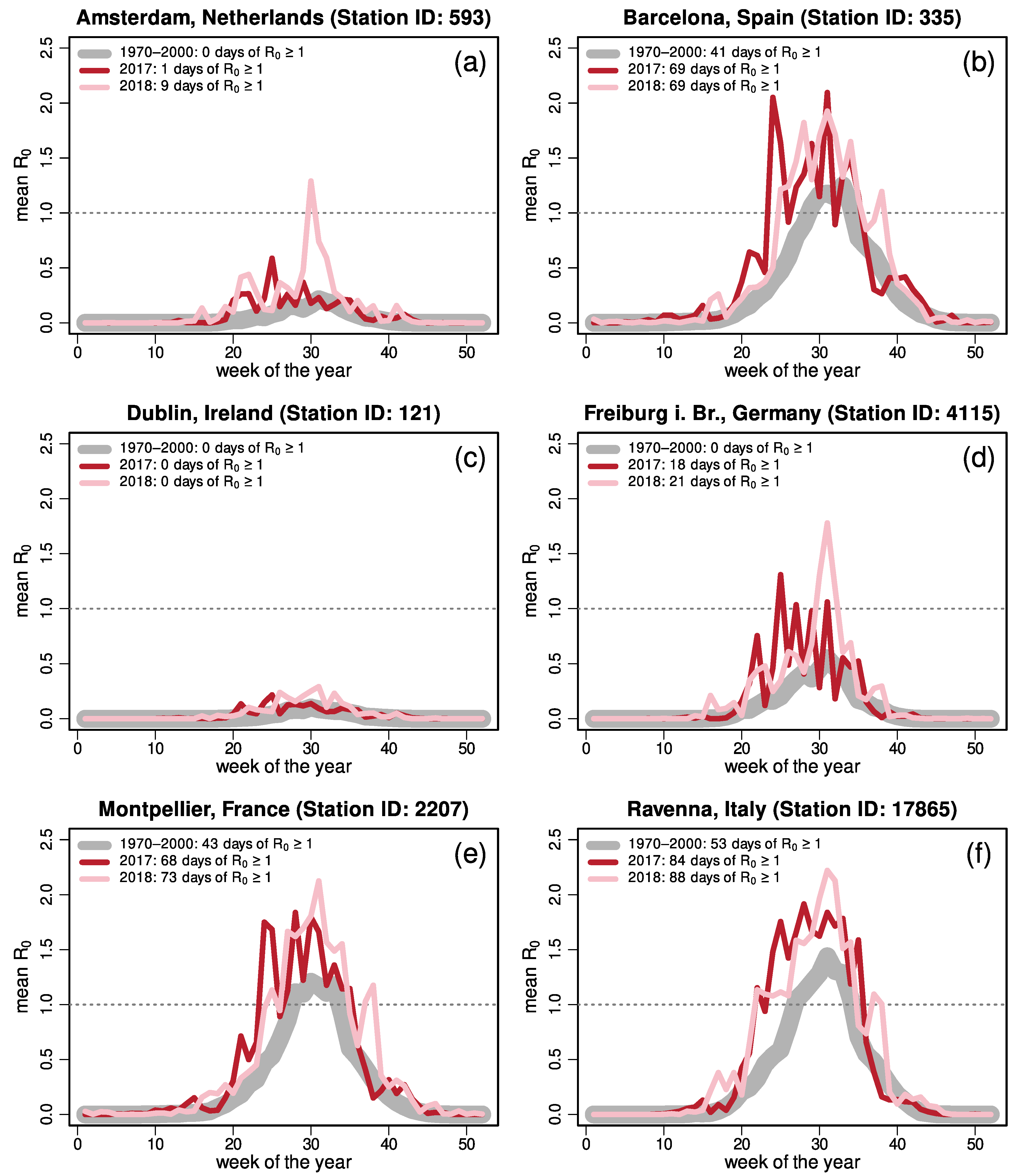{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ecological Niche Models
2.1.1. Occurrence Records
2.1.2. Environmental Data
2.1.3. Calibration Area
2.1.4. Model Selection
2.1.5. Thresholds
2.2. Epidemiological Model
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rezza, G.; Weaver, S.C. Chikungunya as a paradigm for emerging viral diseases: Evaluating disease impact and hurdles to vaccine development. PLoS Negl. Trop. Dis. 2019, 13, e0006919. [Google Scholar] [CrossRef] [PubMed]
- Burt, F.J.; Chen, W.Q.; Miner, J.J.; Lenschow, D.J.; Merits, A.; Schnettler, E.; Kohl, A.; Rudd, P.A.; Taylor, A.; Herrero, L.J.; et al. Chikungunya virus: An update on the biology and pathogenesis of this emerging pathogen. Lancet Infect. Dis. 2017, 17, E107–E117. [Google Scholar] [CrossRef]
- Robinson, M.C. An epidemic of virus disease in southern province, Tanganyika territory, in 1952-53. I. Clinical features. Trans R. Soc. Trop. Med. Hyg. 1955, 49, 28–32. [Google Scholar] [CrossRef]
- Valentine, M.J.; Murdock, C.C.; Kelly, P.J. Sylvatic cycles of arboviruses in non-human primates. Parasites Vectors 2019, 12, 463. [Google Scholar] [CrossRef]
- Gérardin, P.; Sampériz, S.; Ramful, D.; Boumahni, B.; Bintner, M.; Alessandri, J.L.; Carbonnier, M.; Tiran-Rajaoefera, I.; Beullier, G.; Boya, I.; et al. Neurocognitive outcome of children exposed to perinatal mother-to-child chikungunya virus infection: The CHIMERE cohort study on Reunion Island. PLoS Negl. Trop. Dis. 2014, 8, e2996. [Google Scholar] [CrossRef]
- Torres, J.R.; Falleiros-Arlant, L.H.; Duenas, L.; Pleitez-Navarrete, J.; Salgado, D.M.; Brea-Del Castillo, J. Congenital and perinatal complications of chikungunya fever: A Latin American experience. Int. J. Infect. Dis. 2016, 51, 85–88. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Chen, R.B.; Diallo, M. Chikungunya virus: Role of vectors in emergence from enzootic cycles. Annu. Rev. Entomol. 2020, 65, 313–332. [Google Scholar] [CrossRef] [Green Version]
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its history in Africa and Asia and its spread to new regions in 2013-2014. J. Infect. Dis. 2016, 214, S436–S440. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control, Geographical Distribution of Chikungunya Virus Disease Cases Reported Worldwide. 2019. Available online: https://www.ecdc.europa.eu/en/publications-data/geographical-distribution-chikungunya-virus-disease-cases-reported-worldwide-2019 (accessed on 28 May 2020).
- Centers for Disease Control and Prevention (CDC), Countries and territories Where Chikungunya Cases Have Been Reported (as of September 17, 2019). Available online: https://www.cdc.gov/chikungunya/geo/index.html (accessed on 28 May 2020).
- Enserink, M. Infectious diseases—Massive outbreak draws fresh attention to little-known virus. Science 2006, 311, 1085. [Google Scholar] [CrossRef] [Green Version]
- Pialoux, G.; Gauzere, B.A.; Jaureguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Mavalankar, D.; Shastri, P.; Raman, P. Chikungunya epidemic in India: A major public-health disaster. Lancet Infect. Dis. 2007, 7, 306–307. [Google Scholar] [CrossRef]
- Charrel, R.N.; de Lamballerie, X.; Raoult, D. Chikungunya outbreaks—The globalization of vectorborne diseases. N. Engl. J. Med. 2007, 356, 769–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parola, P.; de Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel chikungunya virus variant in travelers returning from Indian Ocean islands. Emerging Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC), USA-Basically Yellowbook 2020 Information. Chikungunya, Chapter 4: Travel-Related Infectious Diseases. Available online: https://wwwnc.cdc.gov/travel/yellowbook/2020/travel-related-infectious-diseases/chikungunya#box401 (accessed on 28 May 2020).
- Noël, H.; Rizzo, C. Spread of chikungunya from the Caribbean to mainland Central and South America: A greater risk of spillover in Europe? Euro Surveill. 2014, 19, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, I.M.; Eyase, F.; Pollett, S.; Konongoi, S.L.; Joyce, M.G.; Figueroa, K.; Ofula, V.; Koka, H.; Koskei, E.; Nyunja, A.; et al. Global outbreaks and origins of a chikungunya virus variant carrying mutations which may increase fitness for Aedes aegypti: Revelations from the 2016 Mandera, Kenya outbreak. Am. J. Trop. Med. Hyg. 2019, 100, 1249–1257. [Google Scholar] [CrossRef]
- Kabir, I.; Dhimal, M.; Muller, R.; Banik, S.; Haque, U. The 2017 Dhaka chikungunya outbreak. Lancet Infect. Dis. 2017, 17, 1118. [Google Scholar] [CrossRef] [Green Version]
- Rauf, M.; Fatima tuz, Z.; Manzoor, S.; Mehmood, A.; Bhatti, S. Outbreak of chikungunya in Pakistan. Lancet Infect. Dis. 2017, 17, 258. [Google Scholar] [CrossRef] [Green Version]
- Gould, L.H.; Osman, M.S.; Farnon, E.C.; Griffith, K.S.; Godsey, M.S.; Karch, S.; Mulenda, B.; El Kholy, A.; Grandesso, F.; de Radigues, X.; et al. An outbreak of yellow fever with concurrent chikungunya virus transmission in South Kordofan, Sudan, 2005. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1247–1254. [Google Scholar] [CrossRef]
- Pandey, K.; Pandey, B.D.; Chaurasiya, R.R.; Thakur, M.; Neupane, B.; Shah, Y.; Tun, M.M.N.; Morita, K. Evidence of chikungunya virus circulation in the Terai region of Nepal in 2014 and 2015. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 294–299. [Google Scholar] [CrossRef]
- Neto, A.S.L.; Sousa, G.S.; Nascimento, O.J.; Castro, M.C. Chikungunya-attributable deaths: A neglected outcome of a neglected disease. PLoS Negl. Trop. Dis. 2019, 13, e0007575. [Google Scholar] [CrossRef] [Green Version]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.C.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F.; et al. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souarès, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, Southeastern France. Emerging Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef]
- Delisle, E.; Rousseau, C.; Broche, B.; Leparc-Goffart, I.; L’Ambert, G.; Cochet, A.; Prat, C.; Foulongne, V.; Ferre, J.B.; Catelinois, O.; et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Euro Surveill. 2015, 20, 8–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiz, D.; Boussès, P.; Simard, F.; Paupy, C.; Fontenille, D. Autochthonous chikungunya transmission and extreme climate events in southern France. PLoS Negl. Trop. Dis. 2015, 9, e0003854. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control, Epidemiological update: Chikungunya–Europe-2017. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-chikungunya-europe-2017 (accessed on 28 May 2020).
- European Centre for Disease Prevention and Control, Rapid Risk Assessment: Clusters of Autochthonous Chikungunya Cases in Italy. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-clusters-autochthonous-chikungunya-cases-italy (accessed on 28 May 2020).
- European Centre for Disease Prevention and Control, Rapid Risk Assessment: Clusters of Autochthonous Chikungunya Cases in Italy, First Update. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-clusters-autochthonous-chikungunya-cases-italy-first-update (accessed on 28 May 2020).
- Venturi, G.; Di Luca, M.; Fortuna, C.; Remoli, M.E.; Riccardo, F.; Severini, F.; Toma, L.; Del Manso, M.; Benedetti, E.; Caporali, M.G.; et al. Detection of a chikungunya outbreak in Central Italy, August to September 2017. Euro Surveill. 2017, 22, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Rezza, G. Chikungunya is back in Italy: 2007–2017. J. Travel Med. 2018, 25, tay004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calba, C.; Guerbois-Galla, M.; Franke, F.; Jeannin, C.; Auzet-Caillaud, M.; Grard, G.; Pigaglio, L.; Cadiou, B.; de Lamballerie, X.; Paty, M.-C.; et al. Investigation of an autochthonous chikungunya outbreak, July–September 2017, France. Revue d’Épidémiologie et de Santé Publique 2018, 66, S387–S388. [Google Scholar]
- European Centre for Disease Prevention and Control, Rapid Risk Assessment: Cluster of Autochthonous Chikungunya Cases in France. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-cluster-autochthonous-chikungunya-cases-france (accessed on 28 May 2020).
- Cebrián-Camisón, S.; Martínez-de la Puente, J.; Figuerola, J. A literature review of host feeding patterns of invasive Aedes mosquitoes in Europe. Insects 2020, 11, 848. [Google Scholar] [CrossRef]
- Adhami, J.; Murati, N. Prani e mushkonjës Aedes albopictus në shqipëri. Rev Mjekesore 1987, I, 13–16. [Google Scholar]
- Thomas, S.M.; Tjaden, N.B.; van den Bos, S.; Beierkuhnlein, C. Implementing cargo movement into climate based risk assessment of vector-borne diseases. Int. J. Environ. Res. Public Health 2014, 11, 3360–3374. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Geier, M.; Balczun, C.; Bradersen, U.; Huber, K.; Kiel, E.; Kruger, A.; Luhken, R.; Orendt, C.; Plenge-Bonig, A.; et al. Repeated introduction of Aedes albopictus into Germany, July to October 2012. Parasitol. Res. 2013, 112, 1787–1790. [Google Scholar] [CrossRef]
- Thomas, S.M.; Obermayr, U.; Fischer, D.; Kreyling, J.; Beierkuhnlein, C. Low-temperature threshold for egg survival of a post-diapause and non-diapause European aedine strain, Aedes albopictus (Diptera: Culicidae). Parasites Vectors 2012, 5, 100. [Google Scholar] [CrossRef] [Green Version]
- Tippelt, L.; Werner, D.; Kampen, H. Tolerance of three Aedes albopictus strains (Diptera: Culicidae) from different geographical origins towards winter temperatures under field conditions in northern Germany. PLoS ONE 2019, 14, e0219553. [Google Scholar] [CrossRef] [Green Version]
- Kramer, I.M.; Kreß, A.; Klingelhöfer, D.; Scherer, C.; Phuyal, P.; Kuch, U.; Ahrens, B.; Groneberg, D.A.; Dhimal, M.; Müller, R. Does winter cold really limit the dengue vector Aedes aegypti in Europe? Parasites Vectors 2020, 13, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, A.T. Mapping Disease Transmission Risk: Enriching Models Using Biogeography and Ecology; Johns Hopkins University Press: Baltimore, MD, USA, 2014; p. 210. [Google Scholar]
- Tjaden, N.B.; Caminade, C.; Beierkuhnlein, C.; Thomas, S.M. Mosquito-borne diseases: Advances in modelling climate-change impacts. Trends Parasitol. 2018, 34, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Samy, A.M.; Thomas, S.M.; Abd El Wahed, A.; Cohoon, K.P.; Peterson, A.T. Mapping the global geographic potential of Zika virus spread. Memorias Do Instituto Oswaldo Cruz 2016, 111, 559–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samy, A.M.; Alkishe, A.A.; Thomas, S.M.; Wang, L.Y.; Zhang, W.Y. Mapping the potential distributions of etiological agent, vectors, and reservoirs of Japanese Encephalitis in Asia and Australia. Acta Trop. 2018, 188, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Nsoesie, E.O.; Kraemer, M.U.; Golding, N.; Pigott, D.M.; Brady, O.J.; Moyes, C.L.; Johansson, M.A.; Gething, P.W.; Velayudhan, R.; Khan, K.; et al. Global distribution and environmental suitability for Chikungunya virus, 1952 to 2015. Euro Surveill. 2016, 21, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjaden, N.B.; Suk, J.E.; Fischer, D.; Thomas, S.M.; Beierkuhnlein, C.; Semenza, J.C. Modelling the effects of global climate change on Chikungunya transmission in the 21st century. Sci. Rep. 2017, 7, 3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasello, D.; Schlagenhauf, P. Chikungunya and dengue autochthonous cases in Europe, 2007–2012. Travel Med. Infect. Dis. 2013, 11, 274–284. [Google Scholar] [CrossRef]
- Ng, V.; Fazil, A.; Gachon, P.; Deuymes, G.; Radojević, M.; Mascarenhas, M.; Garasia, S.; Johansson, M.A.; Ogden, N.H. Assessment of the probability of autochthonous transmission of chikungunya virus in Canada under recent and projected climate change. Environ. Health Perspect. 2017, 125, 067001. [Google Scholar] [CrossRef] [Green Version]
- Pecchi, M.; Marchi, M.; Burton, V.; Giannetti, F.; Moriondo, M.; Bernetti, I.; Bindi, M.; Chirici, G. Species distribution modelling to support forest management. A literature review. Ecol. Modell. 2019, 411, 108817. [Google Scholar] [CrossRef]
- Valencia-Rodríguez, D.; Jiménez-Segura, L.; Rogéliz, C.A.; Parra, J.L. Ecological niche modeling as an effective tool to predict the distribution of freshwater organisms: The case of the Sabaleta Brycon henni (Eigenmann, 1913). PLoS ONE 2021, 16, e0247876. [Google Scholar] [CrossRef]
- Vetter, V.M.S.; Tjaden, N.B.; Jaeschke, A.; Buhk, C.; Wahl, V.; Wasowicz, P.; Jentsch, A. Invasion of a legume ecosystem engineer in a cold biome alters plant biodiversity. Front. Plant Sci. 2018, 9, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beukema, W.; Bruni, G. New records and a revision of the actual and potential distribution of Discoglossus montalentii to facilitate future conservation assessments. Amphib. Reptil. 2020, 41, 269–274. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006; 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.C.; Tjaden, N.B.; Jaeschke, A.; Thomas, S.M.; Beierkuhnlein, C. Using centroids of spatial units in ecological niche modelling: Effects on model performance in the context of environmental data grain size. Global Ecol. Biogeogr. 2021, 30, 611–621. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. Available online: http://www.R-project.org/ (accessed on 28 May 2020).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- VanDerWal, J.; Shoo, L.P.; Graham, C.; William, S.E. Selecting pseudo-absence data for presence-only distribution modeling: How far should you stray from what you know? Ecol. Modell. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Tjaden, N.B.; Jaeschke, A.; Lüken, R.; Ziegler, U.; Thomas, S.M.; Beierkuhnlein, C. Evaluating the risk for Usutu virus circulation in Europe: Comparison of environmental niche models and epidemiological models. Int. J. Health Geogr. 2018, 17. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Scholte, E.-J.; Schafner, F. Waiting for the tiger: Establishment and spread of the Aedes albopictus mosquito in Europe. In Emerging Pests and Vector-Borne Diseases in Europe; Takken, W., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 241–260. [Google Scholar]
- Ahmed, Q.A.; Memish, Z.A. The public health planners’ perfect storm: Hurricane Matthew and Zika virus. Travel Med. Infect. Dis. 2017, 15, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Koenraadt, C.J.M.; Harrington, L.C. Flushing effect of rain on container-inhabiting mosquitoes Aedes aegypti and Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Merow, C.; Smith, M.J.; Edwards, T.C.; Guisan, A.; McMahon, S.M.; Normand, S.; Thuiller, W.; Wuest, R.O.; Zimmermann, N.E.; Elith, J. What do we gain from simplicity versus complexity in species distribution models? Ecography 2014, 37, 1267–1281. [Google Scholar] [CrossRef]
- Bell, D.M.; Schlaepfer, D.R. On the dangers of model complexity without ecological justification in species distribution modeling. Ecol. Modell. 2016, 330, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Galante, P.J.; Alade, B.; Muscarella, R.; Jansa, S.A.; Goodman, S.M.; Anderson, R.P. The challenge of modeling niches and distributions for data-poor species: A comprehensive approach to model complexity. Ecography 2018, 41, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Papes, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Modell. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V. ENMGadgets: Pre and Post Processing in ENM Workflow; R package version 0.1.0.1; 2019. Available online: https://rdrr.io/github/narayanibarve/ENMGadgets/ (accessed on 28 May 2020).
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Liu, C.R.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Liu, C.R.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Bean, W.T.; Stafford, R.; Brashares, J.S. The effects of small sample size and sample bias on threshold selection and accuracy assessment of species distribution models. Ecography 2012, 35, 250–258. [Google Scholar] [CrossRef]
- Liu, C.R.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Tank, A.; Wijngaard, J.B.; Konnen, G.P.; Bohm, R.; Demaree, G.; Gocheva, A.; Mileta, M.; Pashiardis, S.; Hejkrlik, L.; Kern-Hansen, C.; et al. Daily dataset of 20th-century surface air temperature and precipitation series for the European Climate Assessment. Int. J. Climatol. 2002, 22, 1441–1453. [Google Scholar] [CrossRef]
- Zeileis, A.; Grothendieck, G. zoo: S3 infrastructure for regular and irregular time series. J. Stat.Softw. 2005, 14. [Google Scholar] [CrossRef] [Green Version]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An ensemble version of the E-OBS temperature and precipitation data sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control, Aedes albopictus—Current Known Distribution: March 2021. Available online: https://www.ecdc.europa.eu/en/publications-data/aedes-albopictus-current-known-distribution-march-2021 (accessed on 13 April 2021).
- Ramasamy, R.; Surendran, S. Global climate change and its potential impact on disease transmission by salinity-tolerant mosquito vectors in coastal zones. Front. Physiol. 2012, 3, 198. [Google Scholar] [CrossRef] [Green Version]
- Service, M.W. Effects of wind on the behaviour and distribution of mosquitoes and blackflies. Int. J. Biometeorol. 1980, 24, 347–353. [Google Scholar] [CrossRef]
- Poletti, P.; Messeri, G.; Ajellii, M.; Vallorani, R.; Rizzo, C.; Merler, S. Transmission potential of chikungunya virus and control measures: The case of Italy. PLoS ONE 2011, 6, e18860. [Google Scholar] [CrossRef] [PubMed]
- Manica, M.; Guzzetta, G.; Poletti, P.; Filipponi, F.; Solimini, A.; Caputo, B.; della Torre, A.; Rosa, R.; Merler, S. Transmission dynamics of the ongoing chikungunya outbreak in Central Italy: From coastal areas to the metropolitan city of Rome, summer 2017. Euro Surveill. 2017, 22, 2–9. [Google Scholar] [CrossRef]
- Rocklöv, J.; Tozan, Y.; Ramadona, A.; Sewe, M.O.; Sudre, B.; Garrido, J.; de Saint Lary, C.B.; Lohr, W.; Semenza, J.C. Using big data to monitor the introduction and spread of chikungunya, Europe, 2017. Emerging Infect. Dis. 2019, 25, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Solimini, A.G.; Manica, M.; Rosa, R.; della Torre, A.; Caputo, B. Estimating the risk of Dengue, Chikungunya and Zika outbreaks in a large European city. Sci. Rep. 2018, 8, 16435. [Google Scholar] [CrossRef]
- Furuya, H. Estimating vector-borne viral infections in the urban setting of the 2020 Tokyo Olympics, Japan, using mathematical modeling. Tokai J. Exp. Clin. Med. 2017, 42, 160–164. [Google Scholar]
- Manore, C.A.; Ostfeld, R.S.; Agusto, F.B.; Gaff, H.; LaDeau, S.L. Defining the risk of Zika and chikungunya virus transmission in human population centers of the eastern United States. PLoS Negl. Trop. Dis. 2017, 11, e0005255. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero, M.A.; Sillero, N. Evaluating how species niche modelling is affected by partial distributions with an empirical case. Acta Oecol. 2016, 77, 207–216. [Google Scholar] [CrossRef]
- Medley, K.A. Niche shifts during the global invasion of the Asian tiger mosquito, Aedes albopictus Skuse (Culicidae), revealed by reciprocal distribution models. Global Ecol. Biogeogr. 2010, 19, 122–133. [Google Scholar] [CrossRef]
- Wearing, H.J.; Robert, M.A.; Christofferson, R.C. Dengue and chikungunya: Modelling the expansion of mosquito-borne viruses into naïve populations. Parasitology 2016, 143, 860–873. [Google Scholar] [CrossRef]
- Chipwaza, B.; Mugasa, J.P.; Selemani, M.; Amuri, M.; Mosha, F.; Ngatunga, S.D.; Gwakisa, P.S. Dengue and Chikungunya fever among viral diseases in outpatient febrile children in Kilosa district hospital, Tanzania. PLoS Negl. Trop Dis. 2014, 8, e3335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalantri, S.P.; Joshi, R.; Riley, L.W. Chikungunya epidemic: An Indian perspective. Natl. Med. J. India 2006, 19, 315–322. [Google Scholar]
- Fischer, D.; Thomas, S.M.; Suk, J.E.; Sudre, B.; Hess, A.; Tjaden, N.B.; Beierkuhnlein, C.; Semenza, J.C. Climate change effects on Chikungunya transmission in Europe: Geospatial analysis of vector’s climatic suitability and virus’ temperature requirements. Int. J. Health Geogr. 2013, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Liu-Helmersson, J.; Rocklöv, J.; Sewe, M.; Brännström, Å. Climate change may enable Aedes aegypti infestation in major European cities by 2100. Environ. Res. 2019, 172, 693–699. [Google Scholar] [CrossRef]
- Vega-Rúa, A.; Marconcini, M.; Madec, Y.; Manni, M.; Carraretto, D.; Gomulski, L.M.; Gasperi, G.; Failloux, A.B.; Malacrida, A.R. Vector competence of Aedes albopictus populations for chikungunya virus is shaped by their demographic history. Commun. Biol. 2020, 3. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control, Autochthonous Transmission of Chikungunya Virus in EU/EEA, 2007–2017. Available online: https://www.ecdc.europa.eu/en/all-topics-z/chikungunya-virus-disease/surveillance-and-disease-data/autochthonous-transmission (accessed on 20 May 2021).
- Ciota, A.T.; Chin, P.A.; Ehrbar, D.J.; Micieli, M.V.; Fonseca, D.M.; Kramer, L.D. Differential effects of temperature and mosquito genetics determine transmissibility of arboviruses by Aedes aegypti in Argentina. Am. J. Trop. Med. Hyg. 2018, 99, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tjaden, N.B.; Cheng, Y.; Beierkuhnlein, C.; Thomas, S.M. Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe? Viruses 2021, 13, 1024. https://doi.org/10.3390/v13061024
Tjaden NB, Cheng Y, Beierkuhnlein C, Thomas SM. Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe? Viruses. 2021; 13(6):1024. https://doi.org/10.3390/v13061024
Chicago/Turabian StyleTjaden, Nils Benjamin, Yanchao Cheng, Carl Beierkuhnlein, and Stephanie Margarete Thomas. 2021. "Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe?" Viruses 13, no. 6: 1024. https://doi.org/10.3390/v13061024
APA StyleTjaden, N. B., Cheng, Y., Beierkuhnlein, C., & Thomas, S. M. (2021). Chikungunya Beyond the Tropics: Where and When Do We Expect Disease Transmission in Europe? Viruses, 13(6), 1024. https://doi.org/10.3390/v13061024