
Goose Nephritic Astrovirus Infection of Goslings Induces Lymphocyte Apoptosis, Reticular Fiber Destruction, and CD8 T-Cell Depletion in Spleen Tissue

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virues
2.2. Animal Experiment
2.3. Histopathological Examination
2.4. Silver Staining
2.5. Immunohistochemical (IHC) Examination
2.6. Ultrahistopathological Examination
2.7. Virus Determination
2.8. Terminal Deoxynucleotidyl Tranferase-Mediated Nick-End Labeling (TUNEL) Staining
2.9. Quantitative Real-Time PCR (qPCR) Analysis
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
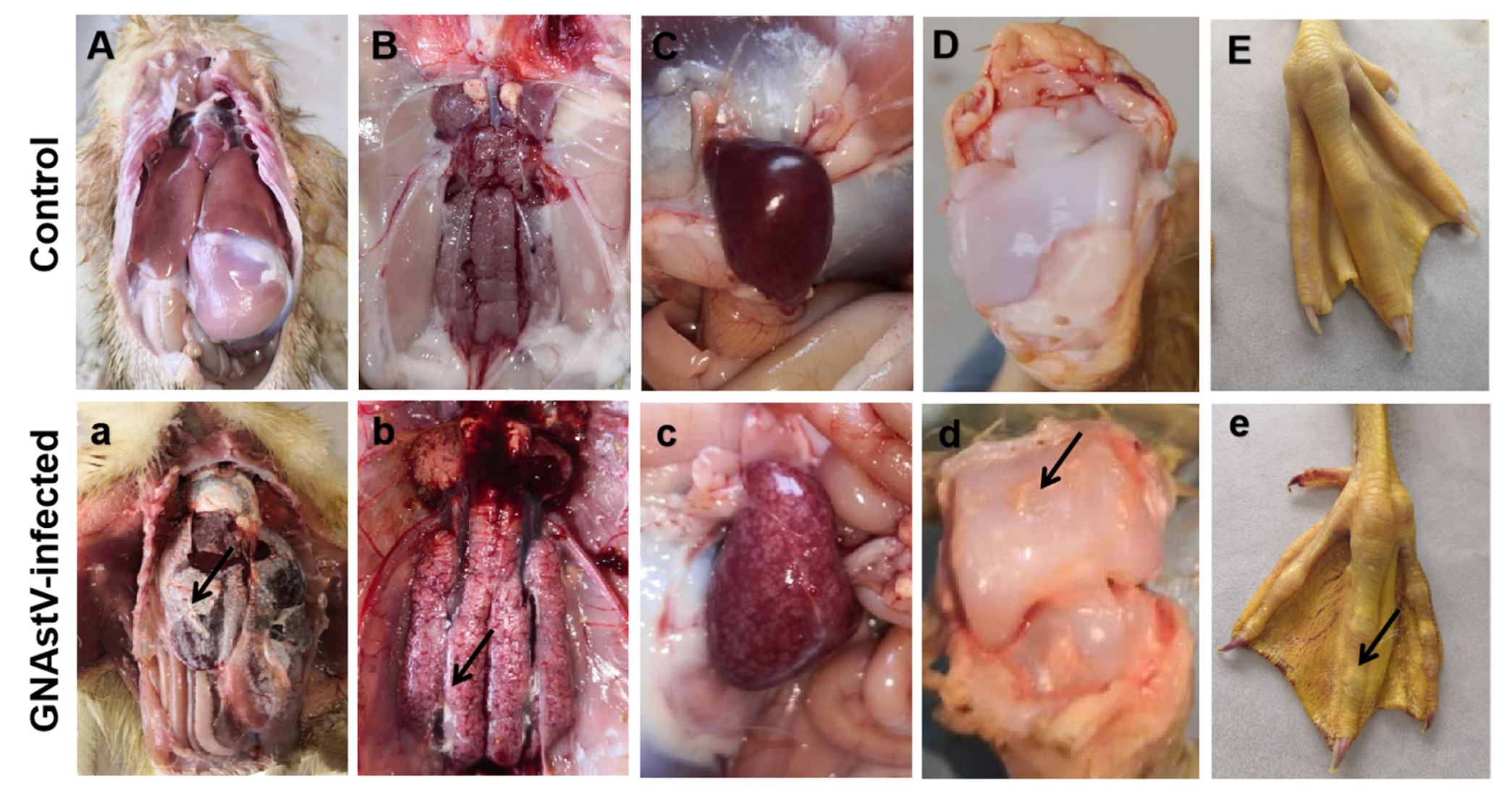
3.1. Clinical Changes
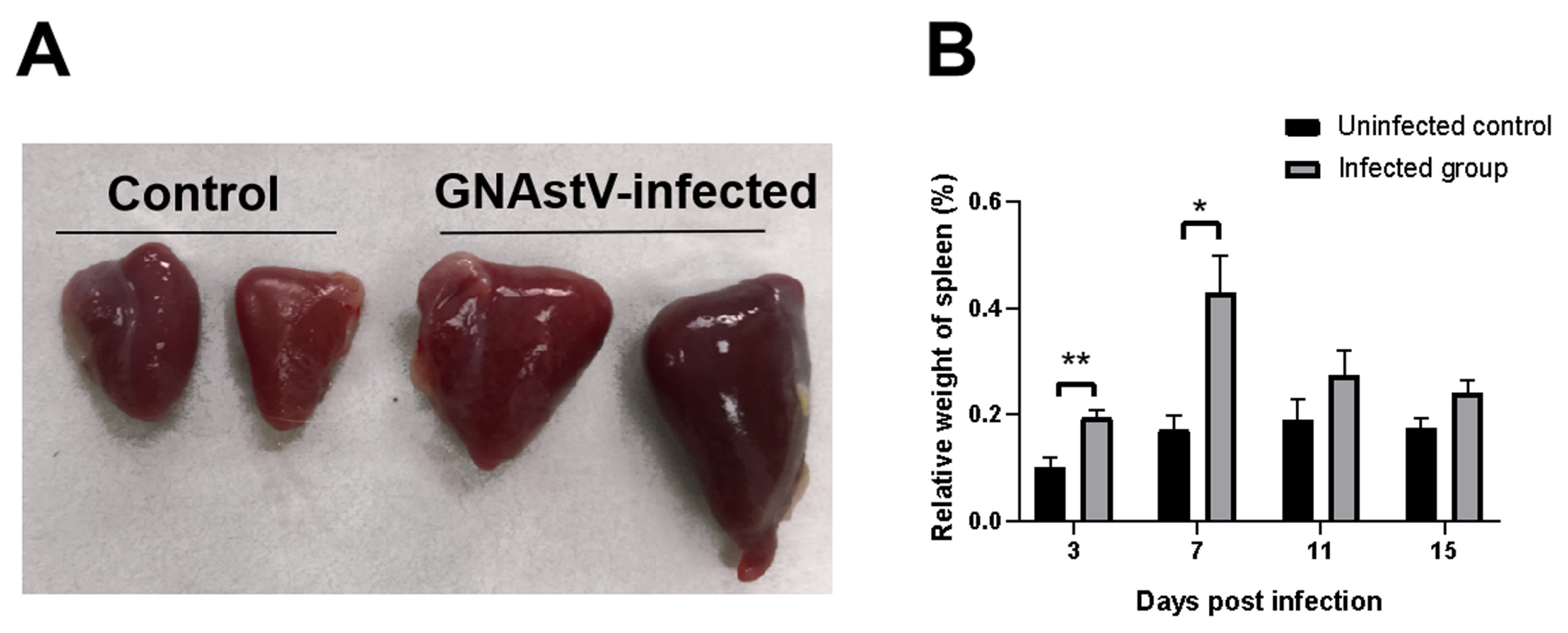
3.2. Gross Morphological Changes in Spleen Tissue
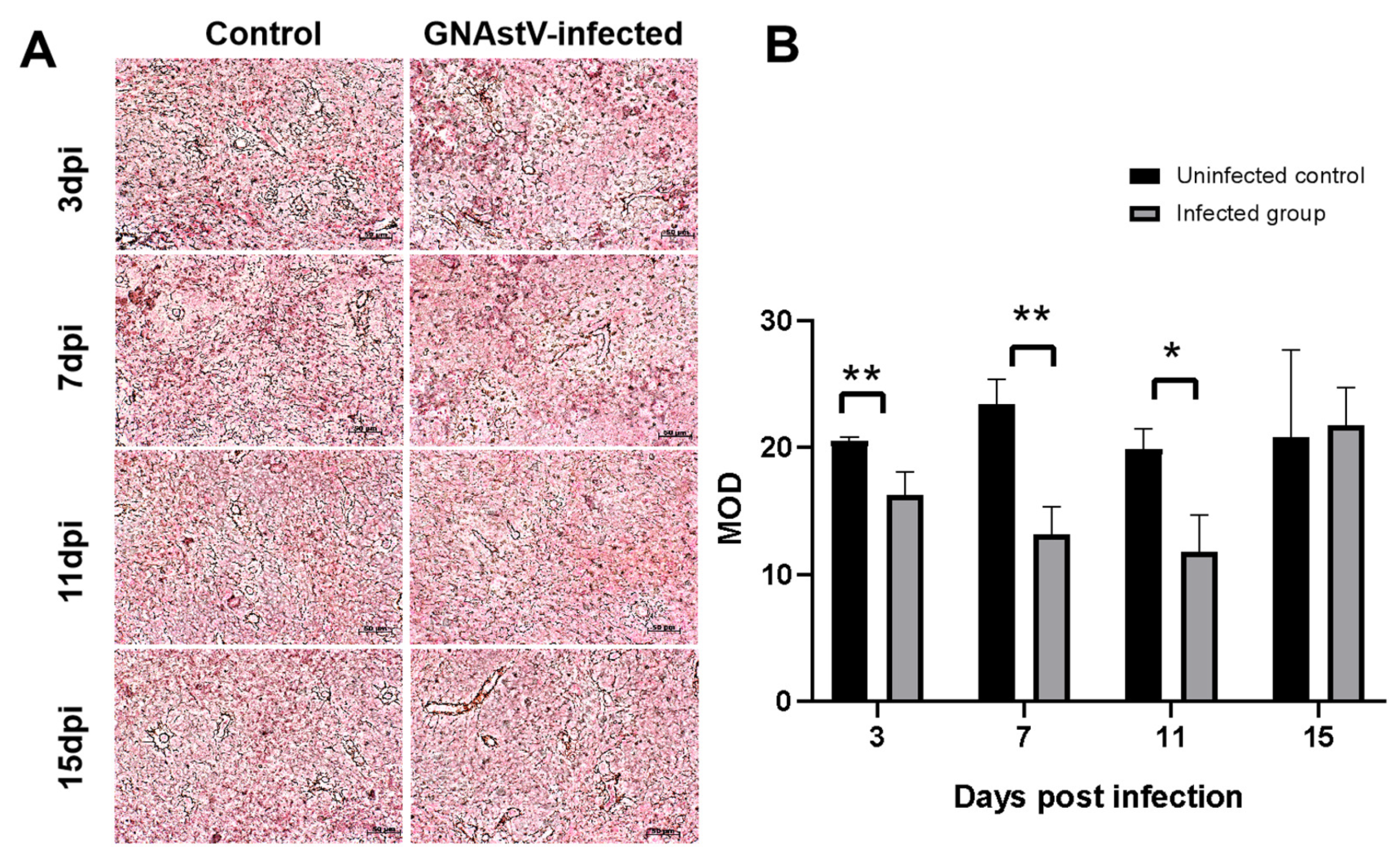
3.3. Histopathological Changes in the Spleen
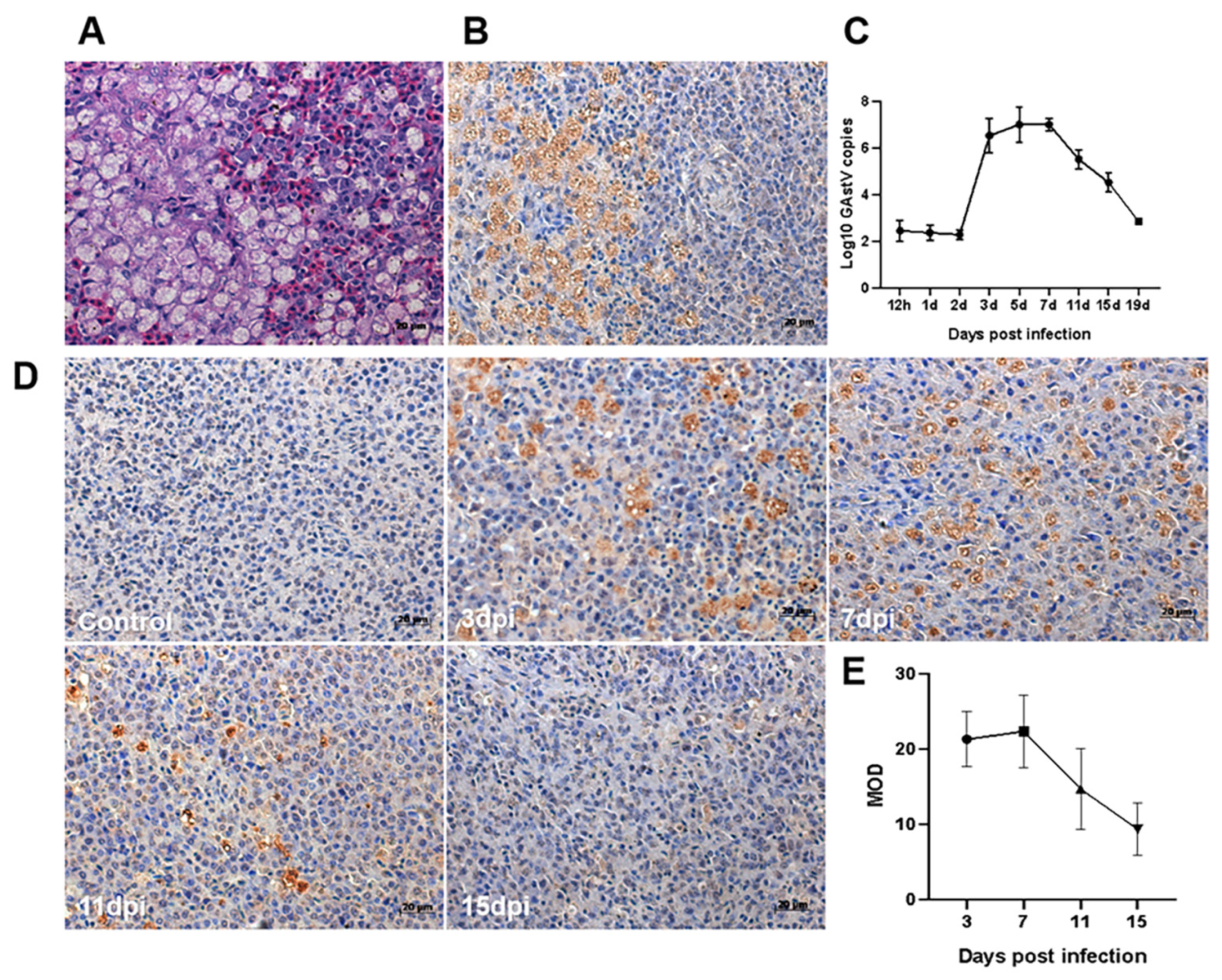
3.4. Virus Location and Viral Load in Spleen Tissue
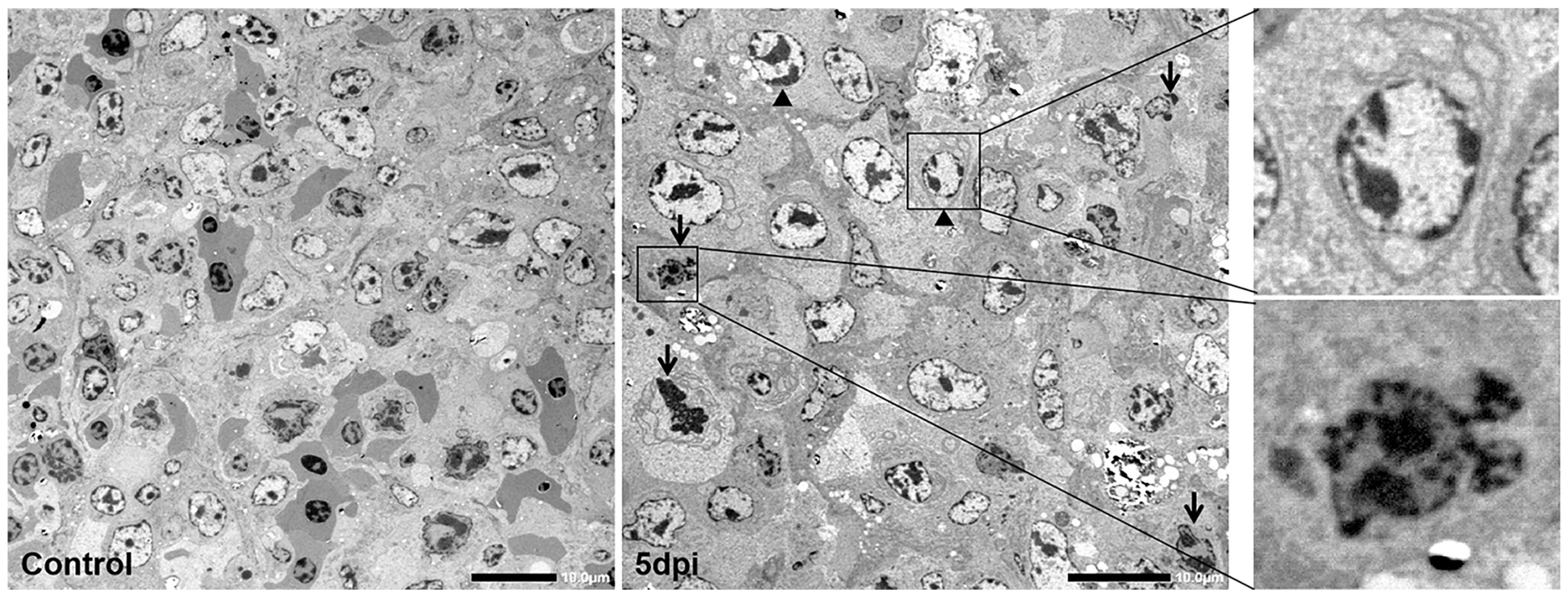
3.5. Ultrahistopathological Changes in Spleen Tissue
3.6. Effects of GNAstV Infection on Apoptosis of Splenic Lymphocytes
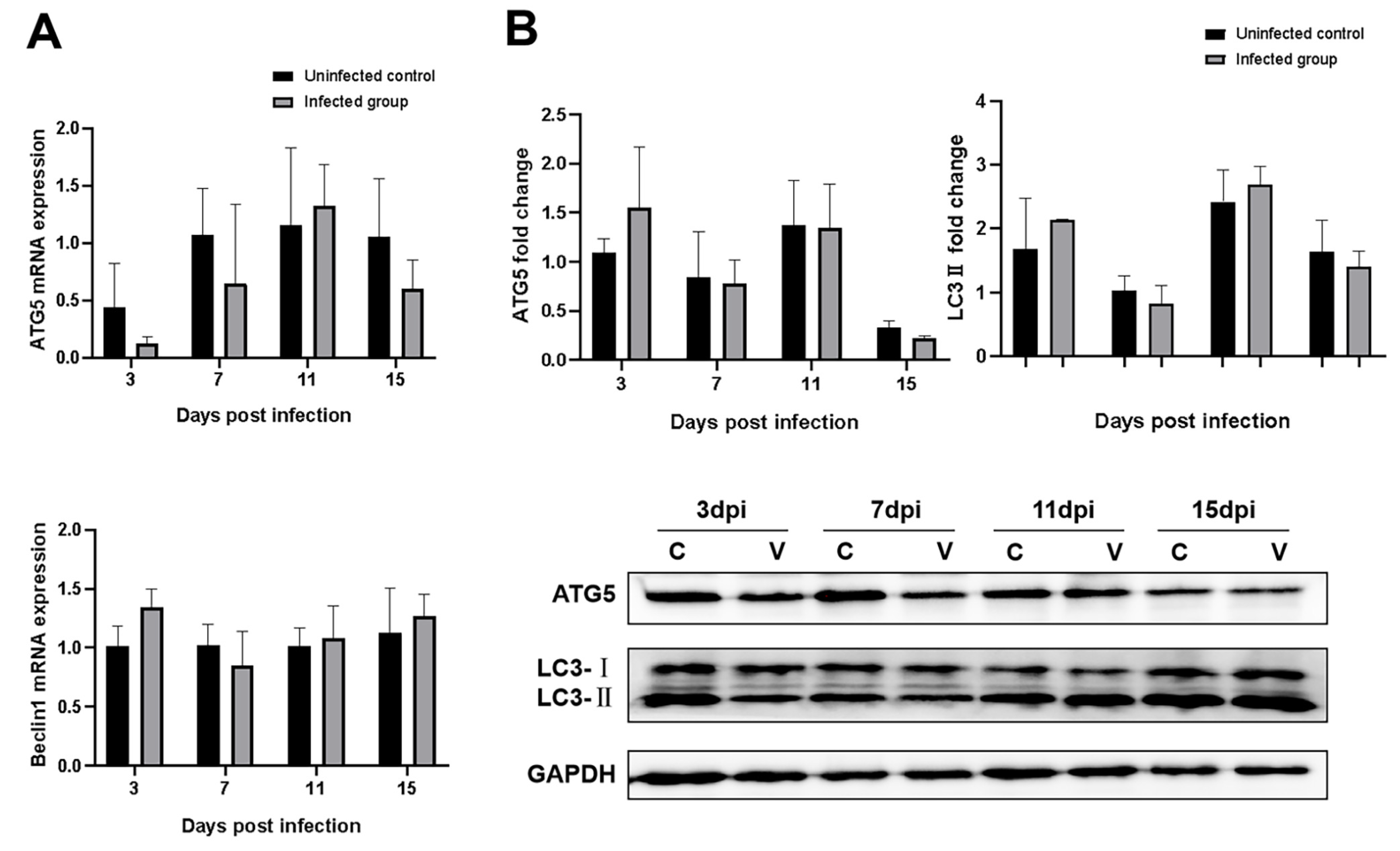
3.7. Effects of GNAstV Infection on Autophagy of Splenic Cells
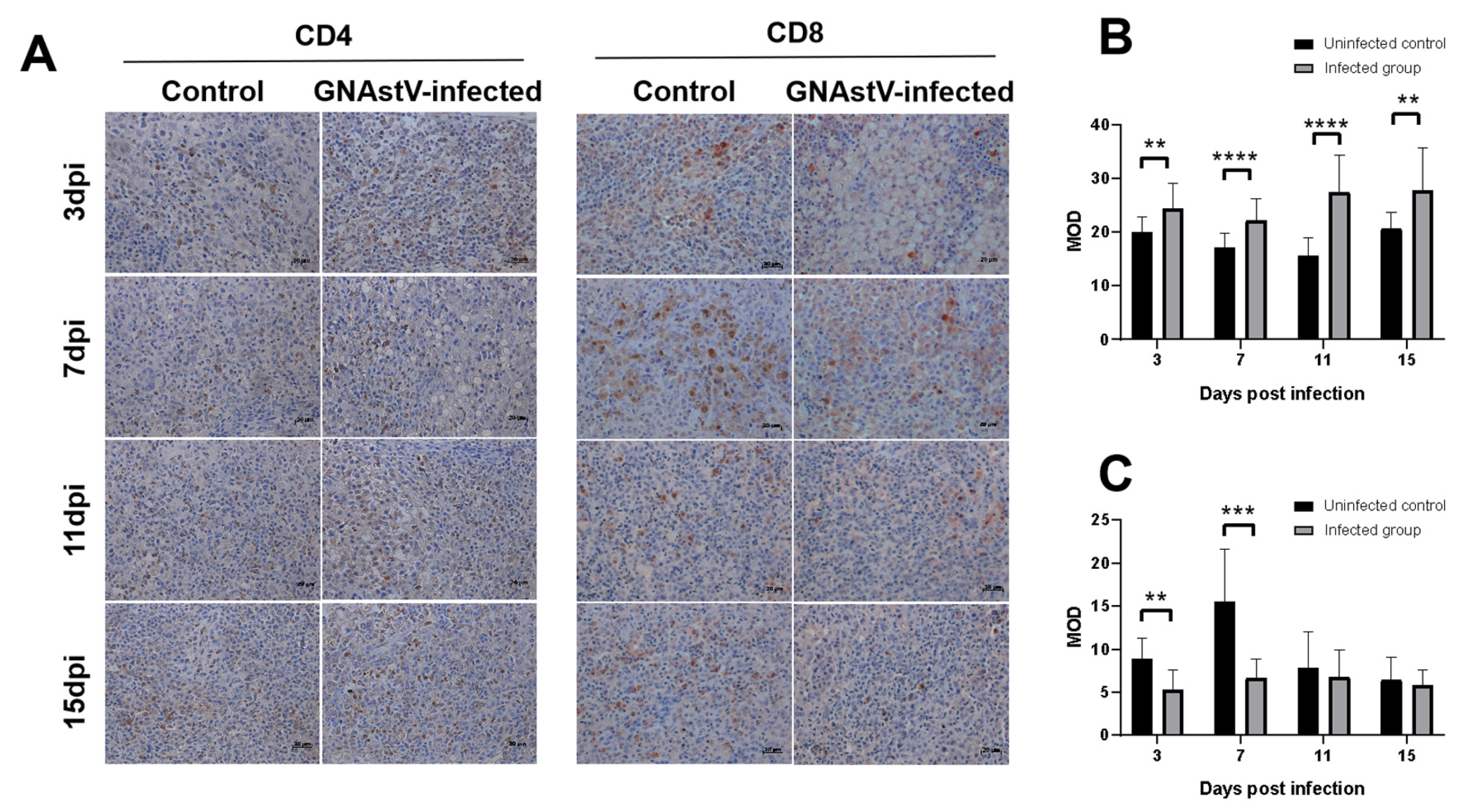
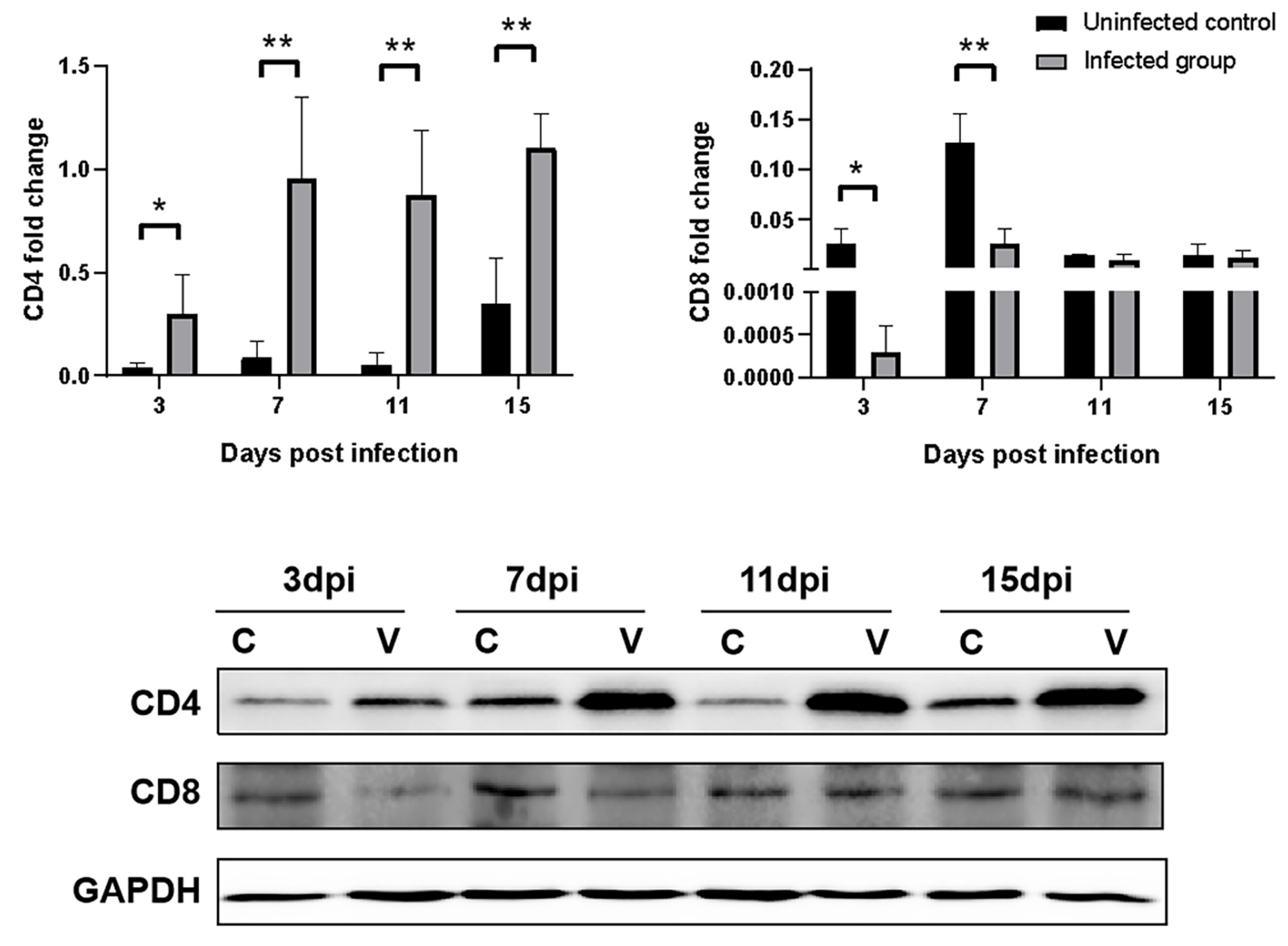
3.8. Effects of GNAstV Infection on CD4 and CD8 Expression in the Spleen
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.; Cao, Y.; Wang, J.; Fu, G.; Sun, M.; Zhang, L.; Meng, L.; Cui, G.; Huang, Y.; Hu, X.; et al. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes. Infect. 2018, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, B.; Yan, M.; Diao, Y.; Tang, Y. First report of a novel goose astrovirus outbreak in Cherry Valley ducklings in China. Transbound. Emerg. Dis. 2020, 67, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Yang, J.; Wang, Y.; Chen, H.; Diao, Y.; Tang, Y. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes. Infec. 2020, 9, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, W.; Liu, T.; Zhang, H.; Opriessnig, T.; Xiao, C. Isolation and evolutionary analyses of gout-associated goose astrovirus causing disease in experimentally infected chickens. Poult. Sci. 2021, 100, 543–552. [Google Scholar] [CrossRef]
- He, D.; Yang, J.; Jiang, X.; Lin, Y.; Chen, H.; Tang, Y.; Diao, Y. A quantitative loop-mediated isothermal amplification assay for detecting a novel goose astrovirus. Poult. Sci. 2020, 99, 6586–6592. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, X.; Yu, Z.; Sui, C.; Zuo, K.; Zhi, G.; Ji, J.; Yao, L.; Kan, Y.; Bi, Y.; et al. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings. Transbound. Emerg. Dis. 2020, 67, 865–876. [Google Scholar] [CrossRef]
- Yin, D.; Yang, J.; Tian, J.; He, D.; Tang, Y.; Diao, Y. Establishment and application of a TaqMan-based one-step real-time RT-PCR for the detection of novel goose-origin astrovirus. J. Virol. Methods 2020, 275, 113757. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, D.; Li, T.; Zhou, H.; Liu, X.; Wang, X.; Lu, H.; Gao, W.; Wang, Y.; Zou, X.; et al. An emerging novel goose astrovirus associated with gosling gout disease. Emerg. Microbes. Infec. 2018, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Tian, J.; Yang, J.; Jiang, X.; Wang, H.; Chen, H.; Yi, T.; Diao, Y. Novel Goose Astrovirus Associated Gout in Gosling. Vet. Microbiol. 2018, 220, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, L.; Liu, P.; Wang, C.; Lu, Q.; Liu, L.; Yang, Y.; Luo, Q.; Shao, H. Host innate immune responses of geese infected with goose origin nephrotic astrovirus. Microb. Pathog. 2021, 152, 104753. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Qiu, S.; Huang, H.; Xu, R.; Bao, E.; Lv, Y. Immune-related gene expression in the kidneys and spleens of goslings infected with goose nephritic astrovirus. Poult. Sci. 2021, 100, 100990. [Google Scholar] [CrossRef]
- Liu, H.; Hu, D.; Zhu, Y.; Xiong, H.; Lv, X.; Wei, C.; Liu, M.; Yin, D.; He, C.; Qi, K.; et al. Coinfection of parvovirus and astrovirus in gout-affected goslings. Transbound. Emerg. Dis. 2020, 67, 2830–2838. [Google Scholar] [CrossRef]
- Wu, W.; Xu, R.; Lv, Y.; Bao, E. Goose astrovirus infection affects uric acid production and excretion in goslings. Poult. Sci. 2020, 99, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and Eosin Staining of Tissue and Cell Sections. Cold Spring Harb. Protoc. 2008, t4986. [Google Scholar] [CrossRef]
- McNeilly, F.; Allan, G.M.; Moffett, D.A.; McNulty, M.S. Detection of chicken anaemia agent in chickens by immunofluorescence and immunoperoxidase staining. Avian Pathol. 1991, 20, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Xu, R.; Guo, Y.; Lv, Y. Establishment of a SYBR Green I-based real-time PCR for goose nephritic astrovirus. Chin. Vet. Sci. 2020, 3, 300–306. (In Chinese) [Google Scholar]
- Sun, J.; Zhang, X.; Cao, Y.; Zhao, Q.; Bao, E.; Lv, Y. Ovarian Toxicity in Female Rats after Oral Administration of Melamine or Melamine and Cyanuric Acid. PLoS ONE 2016, 11, e149063. [Google Scholar] [CrossRef]
- Lou, Y. Study on Follicular Granulisa Cell Autophay Induced by Oxidative Stress in Goose. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2015. (In Chinese). [Google Scholar]
- Huang, B.; Li, J.; Zhang, X.; Zhao, Q.; Lu, M.; Lv, Y. RIG-1 and MDA-5 signaling pathways contribute to IFN-β production and viral replication in porcine circovirus virus type 2-infected PK-15 cells in vitro. Vet. Microbiol. 2017, 211, 36–42. [Google Scholar] [CrossRef]
- An, D.; Zhang, J.; Yang, J.; Tang, Y.; Diao, Y. Novel goose-origin astrovirus infection in geese: The effect of age at infection. Poult. Sci. 2020, 99, 4323–4333. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Li, W.; Yang, S.; Sun, X.; Tarique, I.; Yang, P.; Chen, Q. Morphological characterization of postembryonic development of blood–spleen barrier in duck. Poult. Sci. 2020, 99, 3823–3830. [Google Scholar] [CrossRef]
- Sun, X.; Li, W.; Liu, E.; Huang, H.; Wang, T.; Wang, X.; Shi, Y.; Yang, P.; Chen, Q. In vivo cellular and molecular study on duck spleen infected by duck Tembusu virus. Vet. Microbiol. 2019, 230, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Perez Shibayama, C.; Gil Cruz, C.; Ludewig, B. Fibroblastic reticular cells at the nexus of innate and adaptive immune responses. Immunol. Rev. 2019, 289, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Pellas, T.C.; Weiss, L. Migration pathways of recirculating murine B cells and CD4+ and CD8+ T lymphocytes. Am. J. Anat. 1990, 187, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, K.; Mochizuki, K. Formation of reticular fibers in the developing spleen of the chick embryo. Arch. Histol. Jpn. (Nihon Soshikigaku Kiroku) 1982, 45, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carruthers, V.B.; Cotter, P.A.; Kumamoto, C.A. Microbial Pathogenesis: Mechanisms of Infectious Disease. Cell Host Microbe 2007, 2, 214–219. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Accession Number | Primer Sequences (5′-3′) | Host | Product Length (bp) |
|---|---|---|---|---|
| Beclin1-F | XM_013199763.1 | CGCTGTGCCAGATGTGGAAGG | Anser | 151 bp |
| Beclin1-R | CAGAAGGAATACTGCGAGTTCAAGA [18]. | |||
| ATG5-F | XM_013175657.1 | CCGATTGGTTTGCTCTT | Anser | 226 bp |
| ATG5-R | ATCCCATCCACAGTTGC | |||
| Bax-F | KY788660 | GGACGAGCTGGACAGCAACG | Anas | 159 bp |
| Bax-R | AGGCGGCAGGCGAAGTAGA | |||
| Bcl-2-F | XM_013187395.1 | TGACCGAGTACCTGAACCG | Anser | 154 bp |
| Bcl-2-R | GCTCCCACCAGAACCAAA | |||
| cyt C-F | XM_013171877.1 | AAATGCTCCCAGTGCC | Anser | 159 bp |
| cyt C-R | CATCAGAGTATCCTCACCC | |||
| Fas-F | XM_013171650.1 | CACTCCCACAAGTCAAG | Anser | 163 bp |
| Fas-R | AGTAGGGTTCCATAGGC | |||
| FasL-F | XM_01319016.1 | ATGGAAGATCACGCAAAGC | Anser | 152 bp |
| FasL-R | GGTGGGAAGGGAGCAAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, R.; Huang, H.; Wang, H.; Yi, Z.; Qiu, S.; Lv, Y.; Bao, E. Goose Nephritic Astrovirus Infection of Goslings Induces Lymphocyte Apoptosis, Reticular Fiber Destruction, and CD8 T-Cell Depletion in Spleen Tissue. Viruses 2021, 13, 1108. https://doi.org/10.3390/v13061108
Ding R, Huang H, Wang H, Yi Z, Qiu S, Lv Y, Bao E. Goose Nephritic Astrovirus Infection of Goslings Induces Lymphocyte Apoptosis, Reticular Fiber Destruction, and CD8 T-Cell Depletion in Spleen Tissue. Viruses. 2021; 13(6):1108. https://doi.org/10.3390/v13061108
Chicago/Turabian StyleDing, Rui, Han Huang, Hongyu Wang, Zewen Yi, Siyu Qiu, Yingjun Lv, and Endong Bao. 2021. "Goose Nephritic Astrovirus Infection of Goslings Induces Lymphocyte Apoptosis, Reticular Fiber Destruction, and CD8 T-Cell Depletion in Spleen Tissue" Viruses 13, no. 6: 1108. https://doi.org/10.3390/v13061108
APA StyleDing, R., Huang, H., Wang, H., Yi, Z., Qiu, S., Lv, Y., & Bao, E. (2021). Goose Nephritic Astrovirus Infection of Goslings Induces Lymphocyte Apoptosis, Reticular Fiber Destruction, and CD8 T-Cell Depletion in Spleen Tissue. Viruses, 13(6), 1108. https://doi.org/10.3390/v13061108