
Identification of a Putative Novel Genotype of Avian Hepatitis E Virus from Apparently Healthy Chickens in Southwestern Nigeria

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Viral RNA Extraction
2.3. Molecular Detection and Characterization of aHEV RNA in Fecal and Serum Samples
2.4. Sequencing and Phylogenetic Analyses
2.5. Statistical analysis
3. Results
3.1. Molecular Detection of aHEV
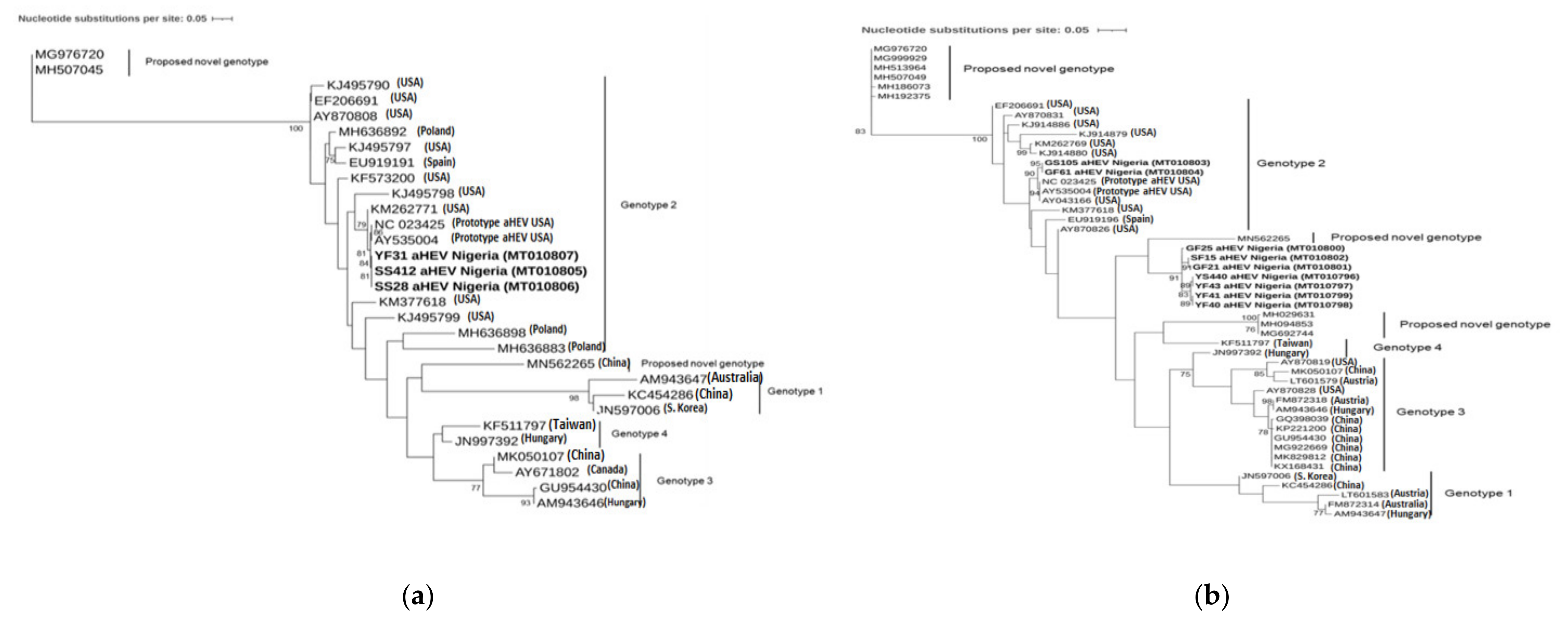
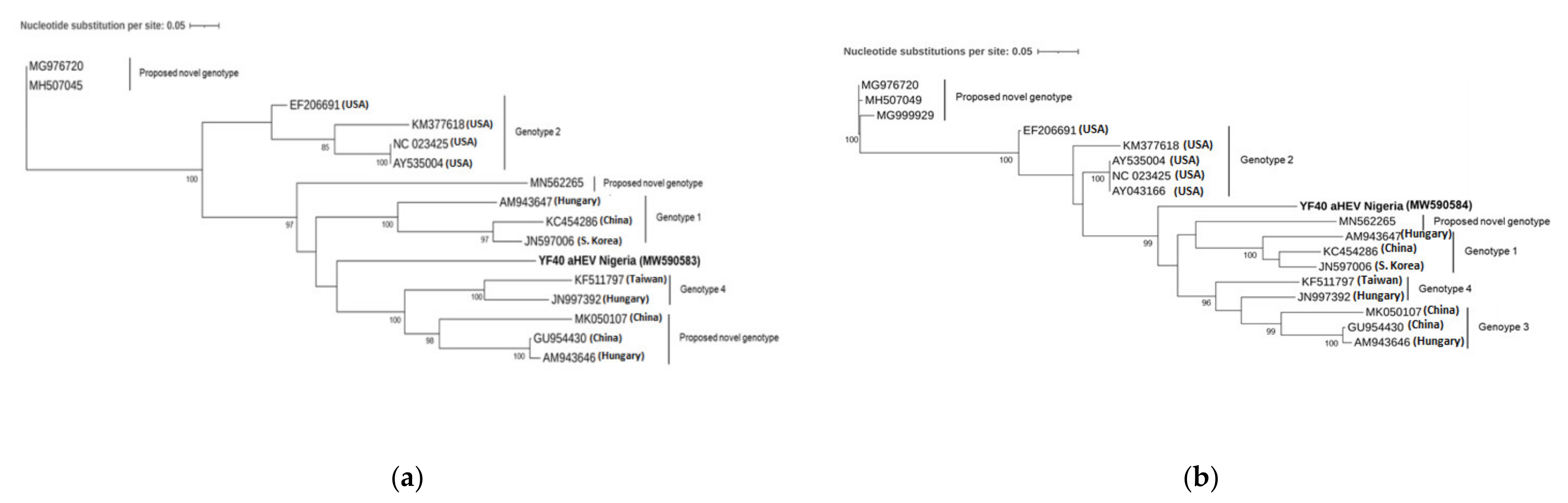
3.2. Phylogenetic Analysis and Genotyping
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, F.F.; Sun, Z.F.; Emerson, S.U.; Purcell, R.H.; Shivaprasad, H.L.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Determination and analysis of the complete genomic sequence of avian hepatitis E virus (avian HEV) and attempts to infect rhesus monkeys with avian HEV. J. Gen. Virol. 2004, 85, 1609–1618. [Google Scholar] [CrossRef]
- Meng, X.J. Hepatitis E virus: Animal reservoirs and zoonotic risk. Vet. Microbiol. 2010, 140, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic Hepatitis E Virus: Classification, Animal Reservoirs and Transmission routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Meng, X.J. Expanding Host Range and Cross species infections of Hepatitis E virus. PLoS Pathog. 2016, 12, e1005695. [Google Scholar] [CrossRef]
- Su, Q.; Li, Y.; Meng, F.; Cui, Z.; Chang, S.; Zhao, P. Hepatic rupture hemorrhage syndrome in chickens caused by a novel genotype avian hepatitis E virus. Vet. Microbiol. 2018, 222, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Li, Y.; Zhang, Y.; Zhang, Z.; Meng, F.; Cui, Z.; Chang, S.; Zhao, P. Characterization of the novel genotype avian hepatitis E viruses from outbreaks of hepatic rupture haemorrhage syndrome in different geographical regions of China. Transbound. Emerg. Dis. 2018, 65, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Zhang, Z.; Zhang, Y.; Cui, Z.; Chang, S.; Zhao, P. Complete genome analysis of avian hepatitis E virus from chicken with hepatic rupture hemorrhage syndrome. Vet. Microbiol. 2020, 242, 108577. [Google Scholar] [CrossRef]
- Handlinger, J.H.; Williams, W. An egg drop associated with splenomegaly in broiler breeders. Avian Dis. 1988, 32, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.J.; Riddel, C. British Columbia.“Hepatitis-splenomegaly” syndrome in commercial egg laying hens. Can. Vet. J. 1991, 32, 500–501. [Google Scholar] [PubMed]
- Payne, C.J.; Ellis, T.M.; Plant, S.L.; Gregory, A.R.; Wilcox, G.E. Sequence data suggests big liver and spleen disease virus (BLSV) is genetically related to hepatitis E virus. Vet. Microbiol. 1999, 68, 119–125. [Google Scholar] [CrossRef]
- Riddell, C. Hepatitis-splenomegaly syndrome. In Diseases of Poultry; Calnek, B.W., Barnes, H.J., Beard, C.W., McDougald, L.R., Saif, Y.M., Eds.; Iowa State University Press: Iowa City, IA, USA, 1997; p. 1041. [Google Scholar]
- Haqshenas, G.; Shivaprasad, H.L.; Woolcock, P.R.; Read, D.H.; Meng, X.J. Genetic identification and characterization of a novel virus related to human hepatitis E virus from chickens with hepatitis-splenomegaly syndrome in the United States. J. Gen. Virol. 2001, 82, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Matczuk, A.K.; Ćwiek, K.; Wieliczko, A. Avian hepatitis E virus is widespread among chickens in Poland and belongs to genotype 2. Arch. Virol. 2019, 164, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.F.; Larsen, C.T.; Dunlop, A.; Huang, F.F.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Genetic identification of avian hepatitis E virus (HEV) from healthy chicken flocks and characterization of the capsid gene of 14 avian HEV isolates from chickens with hepatitis–splenomegaly syndrome in different geographical regions of the United States. J. Gen. Virol. 2004, 85, 693–700. [Google Scholar] [CrossRef]
- Zhao, Q.; Xie, S.; Sun, Y.; Chen, Y.; Gao, J.; Li, H.; Wang, X.; Syed, S.F.; Liu, B.; Wang, L.; et al. Development and evaluation of a SYBR Green real-time RT-PCR assay for detection of avian hepatitis E virus. BMC Vet. Res. 2015, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Purdy, M.A. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Lin, S.; He, S.; Zhou, E.M.; Zhao, Q. Avian Hepatitis E Virus: With the Trend of Genotypes and Host Expansion. Front. Microbiol. 2019, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.M.; Sung, H.W.; Meng, X.J. Serological prevalence, genetic identification, and characterization of the first strains of avian hepatitis E virus from chickens in Korea. Virus Genes 2012, 45, 237–245. [Google Scholar] [CrossRef]
- Peralta, B.; Biarnés, M.; Ordóñez, G.; Porta, R.; Martín, M.; Mateu, E.; Pina, S.; Meng, X.J. Evidence of widespread infection of avian hepatitis E virus (avian HEV) in chickens from Spain. Vet. Microbiol. 2009, 137, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.W.; Lee, B.W.; Sung, H.W.; Yoon, B.I.; Kwon, H.M. Identification and characterization of avian hepatitis E virus genotype 2 from chickens with hepatitis-splenomegaly syndrome in Korea. Virus Genes 2016, 52, 738–742. [Google Scholar] [CrossRef]
- Bilic, I.; Jaskulska, B.; Basic, A.; Morrow, C.J.; Hess, M. Sequence analysis and comparison of avian hepatitis E viruses from Australia and Europe indicate the existence of different genotypes. J. Gen. Virol. 2009, 90, 863–873. [Google Scholar] [CrossRef]
- Sprygin, A.V.; Nikonova, Z.B.; Zinyakov, N.G. Avian hepatitis E virus identified in Russian chicken flocks exhibits high genetic divergence based on the ORF2 capsid gene. Avian Pathol. 2012, 41, 459–463. [Google Scholar] [CrossRef]
- Morrow, C.J.; Samu, G.; Mátrai, E.; Klausz, A.; Wood, A.M.; Richter, S.; Jaskulska, B.; Hess, M. Avian hepatitis E virus infection and possible associated clinical disease in broiler breeder flocks in Hungary. Avian Pathol. 2008, 37, 527–535. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhou, E.M.; Dong, S.W.; Qiu, H.K.; Zhang, L.; Hu, S.B.; Zhao, F.F.; Jiang, S.J.; Sun, Y.N. Analysis of avian hepatitis E virus from chickens, China. Emerg. Infect. Dis. 2010, 16, 1469–1472. [Google Scholar] [CrossRef]
- Bányai, K.; Tóth, Á.G.; Ivanics, É.; Glávits, R.; Szentpáli-Gavallér, K.; Dán, Á. Putative novel genotype of avian hepatitis E virus, Hungary, 2010. Emerg. Infect. Dis. 2012, 18, 1365–1368. [Google Scholar] [CrossRef]
- Hsu, I.W.; Tsai, H.J. Avian hepatitis E virus in chickens, Taiwan, 2013. Emerg. Infect. Dis. 2014, 20, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Kenney, S.P. The current host range of hepatitis E viruses. Viruses 2019, 11, 452. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Chen, Y.; Zhao, L.; Zhang, M.; Ren, X.; Zhang, Y.; Zhang, B.; Fan, M.; Zhao, Q.; Zhou, E.M. Identification and pathogenicity of a novel genotype avian hepatitis E virus from silkie fowl (gallus gallus). Vet. Microbiol. 2020, 245, 108688. [Google Scholar] [CrossRef]
- Kim, J.H.; Nelson, K.E.; Panzner, U.; Kasture, Y.; Labrique, A.B.; Wierzba, T.F. A systematic review of the epidemiology of hepatitis E virus in Africa. BMC Infect. Dis. 2014, 14, 308. [Google Scholar] [CrossRef] [PubMed]
- Buisson, Y.; Grandadam, M.; Nicand, E.; Cheval, P.; van Cuyck-Gandre, H.; Innis, B.; Tsarev, S. Identification of a novel hepatitis E virus in Nigeria. J. Gen. Virol. 2000, 81, 903–909. [Google Scholar] [CrossRef]
- Wang, B.; Akanbi, O.; Harms, D.; Adesina, O.; Osundare, F.; Naidoo, D. A New hepatitis E virus genotype 2 strain identified from an outbreak in Nigeria, 2017. Virol. J. 2018, 15, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owolodun, O.A.; Gerber, P.F.; Giménez-Lirola, L.G.; Kwaga, J.K.; Opriessnig, T. First report of hepatitis E virus circulation in domestic pigs in Nigeria. Am. J. Trop. Med. Hyg. 2014, 91, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Meseko, C.; Odurinde, O.; Odaibo, G.; Olaleye, D. Serological evidence and risk factors associated with hepatitis E virus infection in pigs and human at an intensive piggery complex, Lagos, Nigeria. J. Healthc. Commun. 2016, 1, 1. [Google Scholar]
- Oluwayelu, D.; Adebiyi, A.; Abiola, J.; Akingbola, T.; Cadmus, S. Serologic evidence of hepatitis E virus activity among slaughtered pigs and in selected pig farms in Ibadan, Nigeria: Implications for zoonotic transmission? Trop. Vet. 2017, 35, 91–100. [Google Scholar]
- Ouoba, J.B.; Traore, K.A.; Rouamba, H.; Setondji, K.V.M.; Minoungou, G.L.; Ouoba, B.L.; Doumbia, M. Prevalence of anti-hepatitis E virus antibodies in domestic animal from three representative provinces of Burkina Faso. Vet. Anim. Sci. 2019, 7, 100059. [Google Scholar] [CrossRef]
- Osamudiamen, F.T.; Akanbi, O.A.; Oluwayelu, D.O.; Bock, C.T.; Klink, P. Serological evidence of avian HEV antibodies in apparently healthy chickens in southwest Nigeria. PLoS ONE 2021, 16, e0247889. [Google Scholar] [CrossRef] [PubMed]
- Ogbolu, E.E.; Obi, R.K. Detection of Hepatitis E Virus RNA in Chicken Droppings and Pig Feces in Ogun and Lagos States, South Western, Nigeria. J. Adv. Med. 2016, 12, 1–8. [Google Scholar] [CrossRef]
- Fatunmbi, O.O.; Adene, D.F. A ten year prevalence study of Marek’s disease and avian leukoses at Ibadan, Nigeria. Acta Vet. Brno 1986, 55, 49–53. [Google Scholar] [CrossRef]
- Wakawa, A.M.; Muhammad, Z.K.; Aliyu, H.B.; Mohammed, B. A retrospective analysis of Marek’s disease diagnosed at Poultry Clinic of Ahmadu Bello University, Zaria, Nigeria. J. Vet. Adv. 2012, 2, 424–429. [Google Scholar]
- Abalaka, S.E.; Sani, N.A.; Idoko, I.S.; Tenuche, O.Z.; Oyelowo, F.O.; Ejeh, S.A.; Enem, S.I. Pathological changes associated with an outbreak of colibacillosis in a commercial broiler flock. Sokoto J. Vet. Sci. 2017, 15, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Sani, N.A.; Aliyu, H.B.; Musa, I.W.; Wakawa, A.M.; Abalaka, S.E.; Oladele, S.B.; Sa’idu, L.; Abdu, P.A. A nine-year retrospective study of avian neoplastic diseases in Zaria, Kaduna state, Nigeria. Sokoto J. Vet. Sci. 2017, 15, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. Model Finder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Billam, P.; LeRoith, T.; Pudupakam, R.S.; Pierson, F.W.; Duncan, R.B.; Meng, X.J. Comparative pathogenesis in specific-pathogen-free chickens of two strains of avian hepatitis E virus recovered from a chicken with Hepatitis–Splenomegaly syndrome and from a clinically healthy chicken. Vet. Microbiol. 2009, 139, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.F.; Trampel, D.W.; Willinghan, E.M.; Billam, P.; Meng, X.J.; Opriessnig, T. Subclinical avian hepatitis E virus infection in layer flocks in the United States. Vet. J. 2015, 206, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.F.; Larsen, C.T.; Huang, F.F.; Billam, P.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Generation and infectivity titration of an infectious stock of avian hepatitis E virus (HEV) in chickens and cross-species infection of turkeys with avian HEV. J. Clin. Microbiol. 2004, 42, 2658–2662. [Google Scholar] [CrossRef] [Green Version]
- Billam, P.; Sun, Z.F.; Meng, X.J. Analysis of the complete genomic sequence of an apparently avirulent strain of avian hepatitis E virus (avian HEV) identified major genetic differences compared with the prototype pathogenic strain of avian HEV. J. Gen. Virol. 2007, 88, 1538–1544. [Google Scholar] [CrossRef]
- Tuaillon, E.; Mondain, A.M.; Meroueh, F.; Ottomani, L.; Picot, M.C.; Nagot, N.; Ducos, J. Dried blood spot for hepatitis C virus serology and molecular testing. Hepatology 2010, 51, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.; Meixenberger, K.; Machnowska, P.; Fiedler, S.; Hanke, K.; Hofmann, A.; Kuecherer, C. Robust and sensitive subtype-generic HIV-1 pol genotyping for use with dried serum spots in epidemiological studies. J. Virol. Methods 2018, 259, 32–38. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Primer | Location * | Size (bp) | Sequence (5′-3′) |
|---|---|---|---|
| ORF1 | |||
| aHEV-8f | 1302–1320 | 585 | TCGTCAGCTCGCCACATGG |
| aHEV-13r | 1869–1886 | AAAACAGCAAGGACCTCCa | |
| aHEV-9fn | 1303–1320 | 583 | CGTCAGCTCGCCACATGG |
| aHEV15rn | 1867–1885 | AAACAGCAAGGACCTCCTC | |
| ORF2/3 | |||
| aHEV-3f | 4807–4825 | 739 | CGTGACAACTCAGCCCAGTGb |
| aHEV-25r | 5523–5545 | GGTCAGCTTTGATGGTGTGTGG | |
| aHEV-7fn | 4809–4829 | 732 | GACAACTCAGCACAGTGGAGCc |
| aHEV-26rn | 5520–5540 | AGCTTTGATGGTGTGTGGTGC | |
| ORF2 (Part 1) | |||
| aHEV-27f | 5429–5448 | 370 | ACTCGGCATGGTTGATCTGG |
| aHEV-28r | 5777–5798 | AGCATCCTCAACCGACATGTAC | |
| aHEV-29fn | 5473–5492 | 289 | TATCACCCGGCAACACGAAC |
| a-HEV30rn | 5742–5761 | TCACCGTTCGCATTCCCAAC | |
| ORF2 (Part 2) | |||
| aHEV-24f | 5684–5702 | 796 | TGGCYTGCCCTCGACATTG |
| aHEV-2r0 | 6459–6479 | AGCTTTGATGGTGTGTGGTGC | |
| aHEV-23fn | 5689–5707 | 778 | TGCCCTCGACATTGTTGCG |
| aHEV-21rn | 6448–6466 | TGGTCTTCAGGTGCCTGGC |
| States | Fecal Samples | Serum Samples | ||||
|---|---|---|---|---|---|---|
| Age (w) (Median ± IQR) | No. Tested | No. Positive (%) | Age (w) (Median ± IQR) | No. Tested | No. Positive (%) | |
| Ogun | 30–52 (45 ± 12) | 32 | 3 (9.4) | 24–52 (42 ± 26) | 20 | 2(10.0) |
| Osun | 32–52 (42 ± 11) | 31 | 2(6.5) | 25–52 (42 ± 17) | 29 | 5(17.2) |
| Oyo | 32–48 (45 ± 8) | 47 | 5(10.6) | 24–52 (38 ± 22) | 39 | 4(10.3) |
| Total | 110 | 10(9.1) | 88 | 11(12.5) | ||
| ID | GenBank Number | State | Age (Weeks) | Gene | Type | Genotype |
|---|---|---|---|---|---|---|
| GF25 | MT010800 | Ogun | 45 | Capsid | Fecal | Putative novel genotype |
| GF21 | MT010801 | Ogun | 45 | Capsid | Fecal | Putative novel genotype |
| GF 61 | MT010804 | Ogun | 50 | Capsid | Fecal | Genotype 2 |
| GS105 | MT010803 | Ogun | 24 | Capsid | Serum | Genotype 2 |
| SF15 | MT010802 | Osun | 32 | Capsid | Fecal | Putative novel genotype |
| SS28 | MT010806 | Osun | 25 | Helicase | Serum | Genotype 2 |
| SS412 | MT010805 | Osun | 46 | Helicase | Serum | Genotype 2 |
| YF31 | MT010807 | Oyo | 32 | Helicase | Fecal | Genotype 2 |
| YF40 | MT010798 | Oyo | 38 | Capsid | Fecal | Putative novel genotype |
| MW590583 | PCP/HVR | |||||
| MW590584 | Capsid | |||||
| YF 41 | MT010799 | Oyo | 38 | Capsid | Fecal | Putative novel genotype |
| YF43 | MT010797 | Oyo | 40 | Capsid | Fecal | Putative novel genotype |
| YS440 | MT010796 | Oyo | 50 | Capsid | Serum | Putative novel genotype |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osamudiamen, F.T.; Akanbi, O.A.; Zander, S.; Oluwayelu, D.O.; Bock, C.-T.; Klink, P. Identification of a Putative Novel Genotype of Avian Hepatitis E Virus from Apparently Healthy Chickens in Southwestern Nigeria. Viruses 2021, 13, 954. https://doi.org/10.3390/v13060954
Osamudiamen FT, Akanbi OA, Zander S, Oluwayelu DO, Bock C-T, Klink P. Identification of a Putative Novel Genotype of Avian Hepatitis E Virus from Apparently Healthy Chickens in Southwestern Nigeria. Viruses. 2021; 13(6):954. https://doi.org/10.3390/v13060954
Chicago/Turabian StyleOsamudiamen, Fisayo Temilade, Olusola Aanuoluwapo Akanbi, Steffen Zander, Daniel Oladimeji Oluwayelu, Claus-Thomas Bock, and Patrycja Klink. 2021. "Identification of a Putative Novel Genotype of Avian Hepatitis E Virus from Apparently Healthy Chickens in Southwestern Nigeria" Viruses 13, no. 6: 954. https://doi.org/10.3390/v13060954
APA StyleOsamudiamen, F. T., Akanbi, O. A., Zander, S., Oluwayelu, D. O., Bock, C. -T., & Klink, P. (2021). Identification of a Putative Novel Genotype of Avian Hepatitis E Virus from Apparently Healthy Chickens in Southwestern Nigeria. Viruses, 13(6), 954. https://doi.org/10.3390/v13060954