
Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and PEDV Infection
2.2. Total RNA Extraction and Quality Inspection of Vero Cell
2.3. cDNA Library Construction and High-Throughput Sequencing
2.4. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
2.5. GO Function Annotation Analysis of Differentially Expressed Genes
2.6. KEGG Enrichment Analysis of Differentially Expressed Genes
2.7. Apoptotic Rate Measurement
2.8. Western Blotting Analysis
2.9. MTT Treatments
2.10. Inhibitor Treatments
3. Results
3.1. PEDV Infection Induced the Pathological Effect of Vero Cells at Different Time Points
3.2. Quality Test of Total RNA
3.3. Analysis of Differentially Expressed Genes in Infected Vero Cells Compared to the Uninfected Cells
3.4. qRT-PCR Verification of 5 Seleted Differentially Expressed Genes
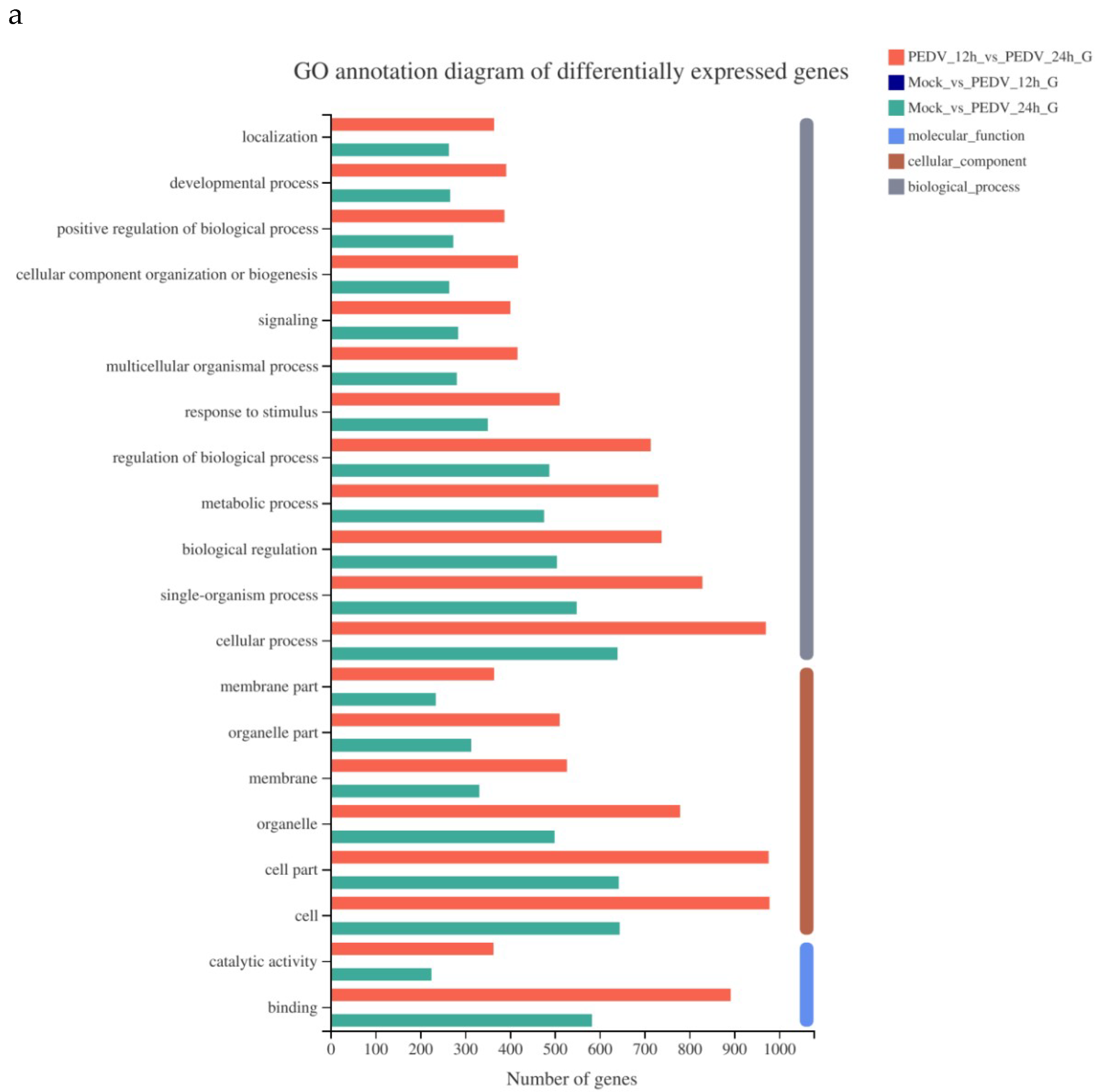
3.5. Analysis of GO Function Annotation of Differentially Expressed Genes
3.6. KEGG Enrichment Analysis of Differentially Expressed Genes
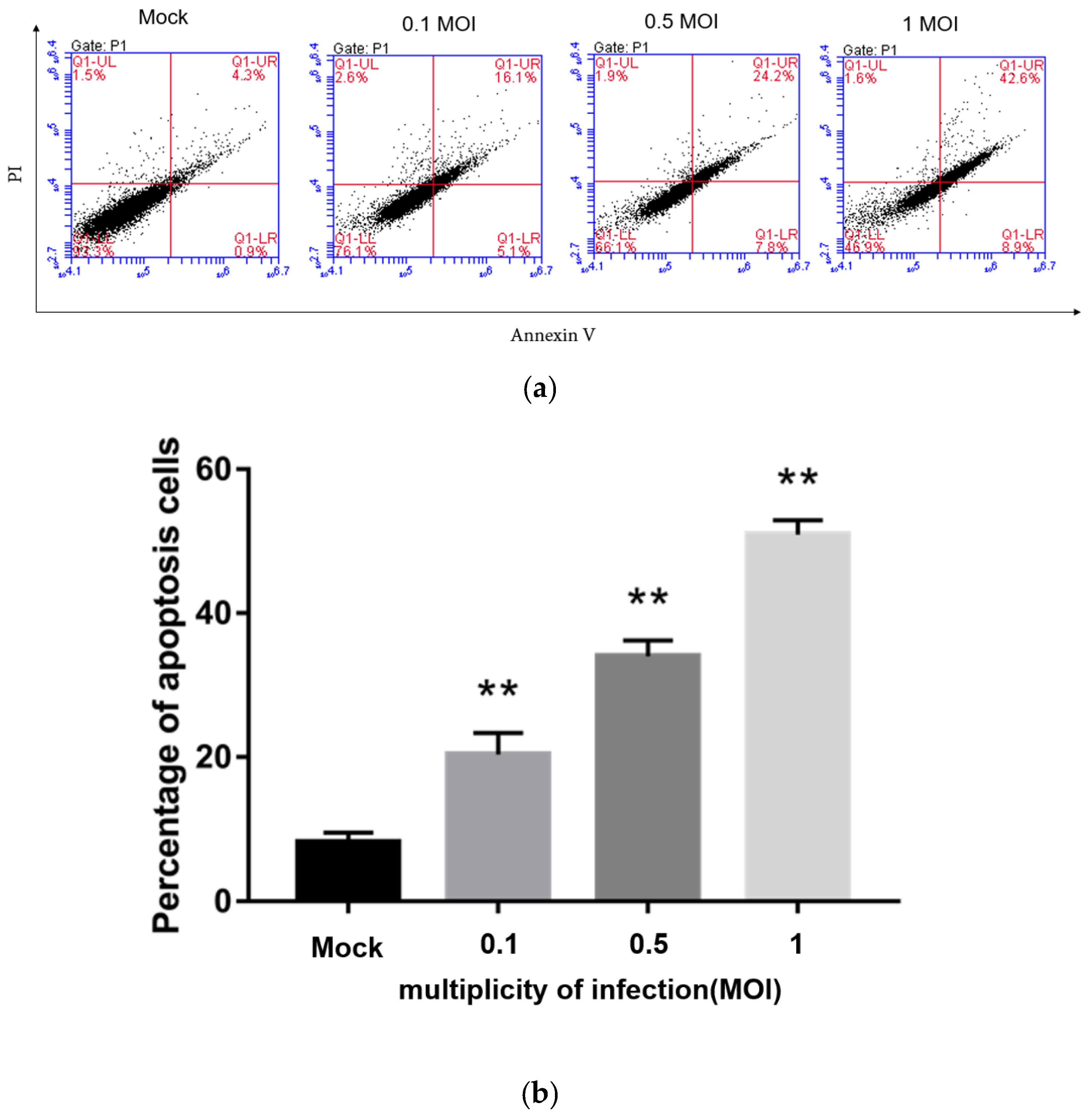
3.7. PEDV Infection Induces Apoptosis in a Dose-Dependent Manner
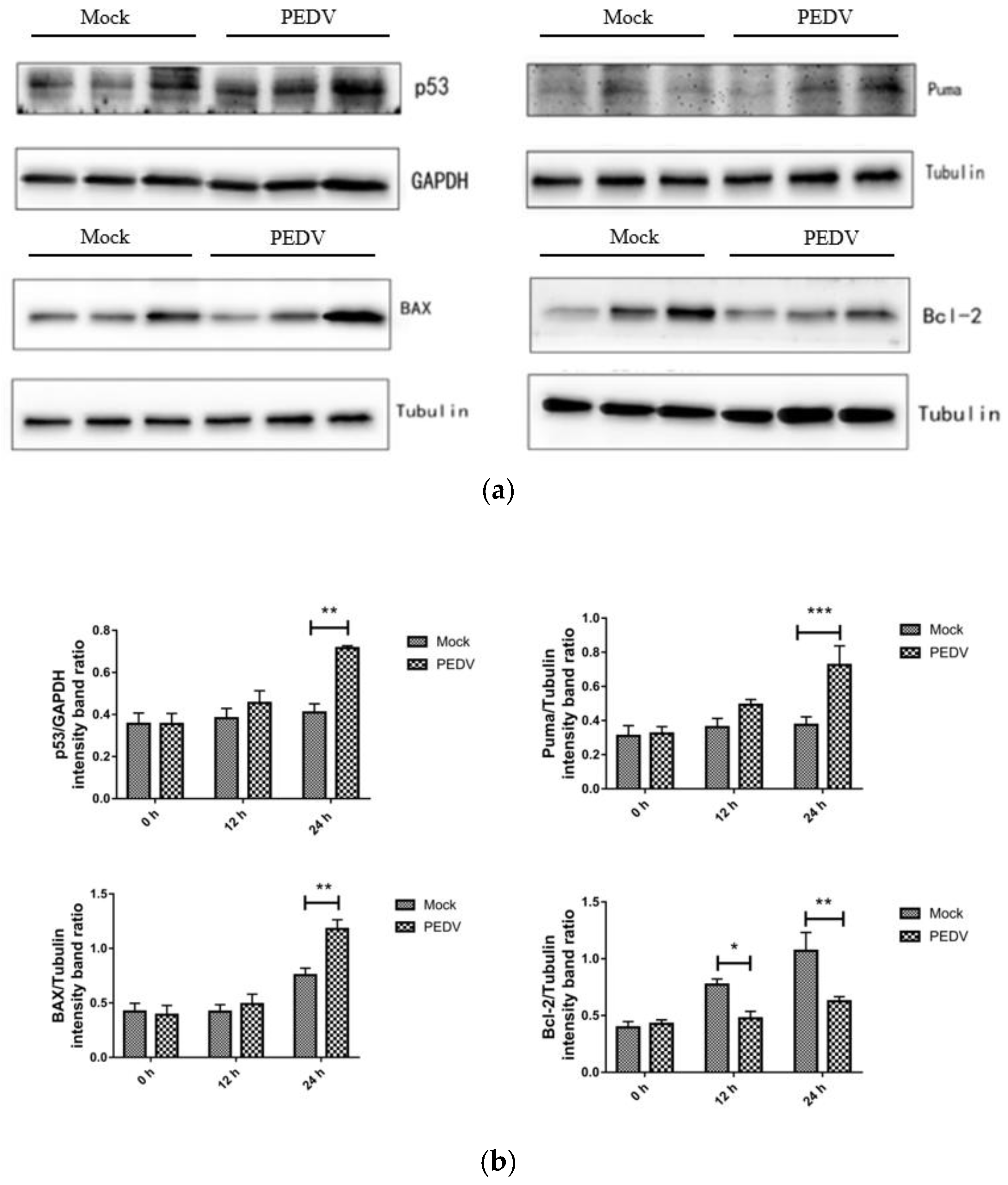
3.8. PEDV Infection Affects the Expression Level of Apoptosis-Related Proteins
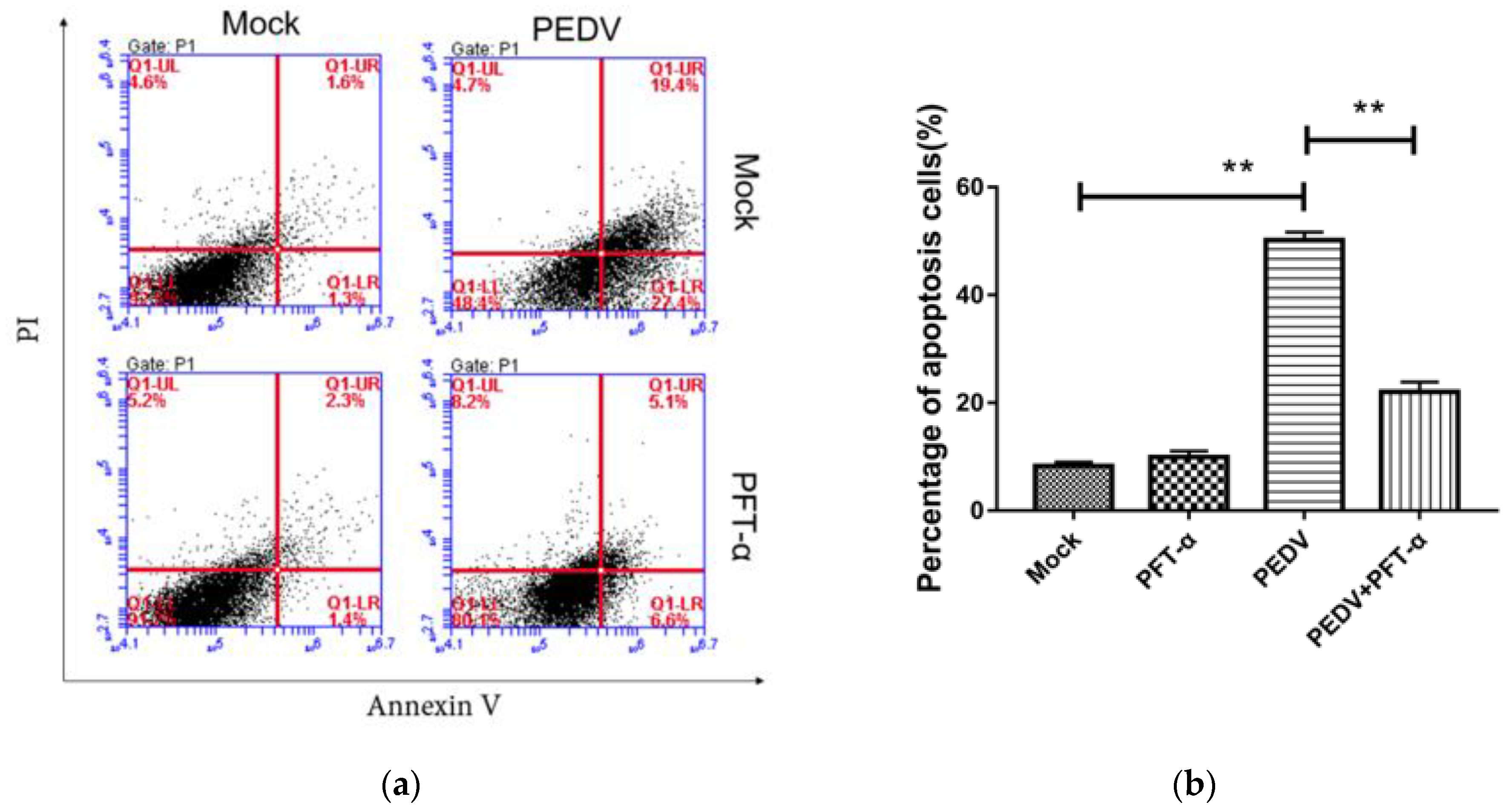
3.9. The Effect of Inhibitor PFT-α on Proteins of p53, Puma, BAX and Bcl-2
3.10. The Inhibitor PFT-α Treatment Inhibits PEDV-Induced Apoptosis with no Effect on Cell Viability
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pensaert, M.B.; De Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Okada, K.; Ohshima, K.-I. An outbreak of swine diarrhea of a new-type associated with coronavirus-like particles in Japan. Jpn. J. Vet. Sci. 1983, 45, 829–832. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Jiao, D.; Zhang, R.; Zhou, J.; Guo, R.; Yu, Z.; Shi, D.; Zhao, Y.; Gu, J.; Niu, B.; et al. Origin and epidemic status of porcine epidemic diarrhea virus variants in China. Transbound. Emerg. Dis. 2020, 67, 1364–1370. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef]
- Brian, D.A.; Baric, R.S. Coronavirus Genome Structure and Replication. Curr. Top. Microbiol. Immunol. 2005, 287, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.-J.; Kim, J.-H.; Lee, C.-H.; Ahn, Y.-J.; Song, J.-H.; Baek, S.-H.; Kwon, D.-H. Antiviral activity of quercetin 7-rhamnoside against porcine epidemic diarrhea virus. Antivir. Res. 2009, 81, 77–81. [Google Scholar] [CrossRef]
- Shi, W.; Fan, W.; Bai, J.; Tang, Y.; Wang, L.; Jiang, Y.; Tang, L.; Liu, M.; Cui, W.; Xu, Y.; et al. TMPRSS2 and MSPL Facilitate Trypsin-Independent Porcine Epidemic Diarrhea Virus Replication in Vero Cells. Viruses 2017, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, P.; Zheng, Q.; Hou, J. Lactobacillus acidophilus S-layer protein-mediated inhibition of PEDV-induced apoptosis of Vero cells. Vet. Microbiol. 2019, 229, 159–167. [Google Scholar] [CrossRef]
- Cai, Y.; Wang, D.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Application of RNAscope technology to studying the infection dynamics of a Chinese porcine epidemic diarrhea virus variant strain BJ2011C in neonatal piglets. Vet. Microbiol. 2019, 235, 220–228. [Google Scholar] [CrossRef]
- Sun, D.; Shi, H.; Guo, D.; Chen, J.; Shi, D.; Zhu, Q.; Zhang, X.; Feng, L. Analysis of protein expression changes of the Vero E6 cells infected with classic PEDV strain CV777 by using quantitative proteomic technique. J. Virol. Methods 2015, 218, 27–39. [Google Scholar] [CrossRef]
- Sun, P.; Wu, H.; Huang, J.; Xu, Y.; Yang, F.; Zhang, Q.; Xu, X. Porcine epidemic diarrhea virus through p53-dependent pathway causes cell cycle arrest in the G0/G1 phase. Virus Res. 2018, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Du, Y.; Wang, G.; Deng, Y.; Li, R.; Li, K. The Novel H7N9 Influenza A Virus NS1 Induces p53-Mediated Apoptosis of A549 Cells. Cell. Physiol. Biochem. 2016, 38, 1447–1458. [Google Scholar] [CrossRef]
- Zhou, P.; Tu, L.; Lin, X.; Hao, X.; Zheng, Q.; Zeng, W.; Zhang, X.; Zheng, Y.; Wang, L.; Li, S. cfa-miR-143 Promotes Apoptosis via the p53 Pathway in Canine Influenza Virus H3N2-Infected Cells. Viruses 2017, 9, 360. [Google Scholar] [CrossRef] [Green Version]
- Inamura, N.; Araki, T.; Enokido, Y.; Nishio, C.; Aizawa, S.; Hatanaka, H. Role of p53 in DNA strand break-induced apoptosis in organotypic slice culture from the mouse cerebellum. J. Neurosci. Res. 2000, 60, 450–457. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, L.; Hwang, P.M.; Kinzler, K.W.; Vogelstein, B. PUMA Induces the Rapid Apoptosis of Colorectal Cancer Cells. Mol. Cell 2001, 7, 673–682. [Google Scholar] [CrossRef]
- Nakano, K.; Vousden, K.H. PUMA, a Novel Proapoptotic Gene, Is Induced by p53. Mol. Cell 2001, 7, 683–694. [Google Scholar] [CrossRef]
- Spender, L.C.; Carter, M.J.; O’Brien, D.I.; Clark, L.J.; Yu, J.; Michalak, E.M.; Happo, L.; Cragg, M.S.; Inman, G.J. Transforming growth factor-beta directly induces p53-up-regulated modulator of apoptosis (PUMA) during the rapid induction of apoptosis in myc-driven B-cell lymphomas. J. Biol. Chem. 2013, 288, 5198–5209. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-W.; Flemington, C.; Houghton, A.B.; Gu, Z.; Zambetti, G.P.; Lutz, R.J.; Zhu, L.; Chittenden, T. Expression of bbc3, a pro-apoptotic BH3-only gene, is regulated by diverse cell death and survival signals. Proc. Natl. Acad. Sci. USA 2001, 98, 11318–11323. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Xu, Y.; Zhang, Q.; Yang, F.; Yin, Z.; Wang, L.; Li, Q. Porcine epidemic diarrhea virus infections induce apoptosis in Vero cells via a reactive oxygen species (ROS)/p53, but not p38 MAPK and SAPK/JNK signalling pathways. Vet. Microbiol. 2019, 232, 1–12. [Google Scholar] [CrossRef]
- Oh, J.; Lee, K.-W.; Choi, H.-W.; Lee, C. Immunogenicity and protective efficacy of recombinant S1 domain of the porcine epidemic diarrhea virus spike protein. Arch. Virol. 2014, 159, 2977–2987. [Google Scholar] [CrossRef] [PubMed]
- Kreis, N.N.; Louwen, F.; Yuan, J. The Multifaceted p21 (Cip1/Waf1/CDKN1A) in Cell Differentiation, Migration and Cancer Therapy. Cancers 2019, 11, 1220. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Li, J.; Wu, G.; Zhou, L.; Wang, X.; Yan, Y.; Xu, Z. The novel roles of virus infection-associated gene CDKN1A in chemoresistance and immune infiltration of glioblastoma. Aging 2021, 13, 6662–6680. [Google Scholar] [CrossRef]
- Guimaraes, D.P.; Oliveira, I.M.; De Moraes, E.; Paiva, G.R.; Souza, D.M.; Barnas, C.; Olmedo, D.B.; Pinto, C.E.; Faria, P.A.; Gallo, C.V.D.M.; et al. Interferon-inducible guanylate binding protein (GBP)-2: A novel p53-regulated tumor marker in esophageal squamous cell carcinomas. Int. J. Cancer 2009, 124, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-H.; Kuhar, D.; Lunney, J.K.; Dawson, H.; Guidry, C.; Uthe, J.J.; Bearson, S.M.; Recknor, J.; Nettleton, D.; Tuggle, C.K. Gene expression profiling in Salmonella Choleraesuis-infected porcine lung using a long oligonucleotide microarray. Mamm. Genome 2006, 17, 777–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.L.; Carton, J.M.; Lou, J.; Xing, L.; Rubin, B.Y. Interferon-Induced Guanylate Binding Protein-1 (GBP-1) Mediates an Antiviral Effect against Vesicular Stomatitis Virus and Encephalomyocarditis Virus. Virology 1999, 256, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, C.C.; Gorbacheva, V.Y.; Vestal, D.J. Inhibition of VSV and EMCV replication by the interferon-induced GTPase, mGBP-2: Differential requirement for wild-type GTP binding domain. Arch. Virol. 2005, 150, 1213–1220. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′ to 3′) |
|---|---|
| β-actin F | CTTCCTGGGTGAGTGGAGAC |
| β-actin R | GAAGGTAGTTTCATGGATGCC |
| CDKN1A F | CAGGAGGCTCGTGAACGATG |
| CDKN1A R | CCTGTGGGAAGGTAGAGCTTG |
| CTPS1 F | CAAGGAACGGAAAGGAGATTACTTGG |
| CTPS1 R | CACGGTTCCACCAAGCTCAATAAC |
| SQSTM1 F | CCTTCGGGCACCTGTCTGAG |
| SQSTM1 R | GATGGACCAGCAGCTGATTC |
| TK1 F | CCTTCGGGCACCTGTCTGAG |
| TK1 R | GATGGACCAGCAGCTGATTC |
| All DEGs | Down-Regulated Genes | Up-Regulated Genes | |
|---|---|---|---|
| Mock vs. PEDV 12 h | 4 | 1 | 3 |
| Mock vs. PEDV 24 h | 1498 | 956 | 542 |
| PEDV 12 h vs. PEDV 24 h | 1643 | 1079 | 564 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wang, C.; Shu, J.; Feng, H.; He, Y.; Chen, J.; Shu, J. Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway. Viruses 2021, 13, 1218. https://doi.org/10.3390/v13071218
Yang L, Wang C, Shu J, Feng H, He Y, Chen J, Shu J. Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway. Viruses. 2021; 13(7):1218. https://doi.org/10.3390/v13071218
Chicago/Turabian StyleYang, Lin, Chenyu Wang, Jinqi Shu, Huapeng Feng, Yulong He, Jian Chen, and Jianhong Shu. 2021. "Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway" Viruses 13, no. 7: 1218. https://doi.org/10.3390/v13071218
APA StyleYang, L., Wang, C., Shu, J., Feng, H., He, Y., Chen, J., & Shu, J. (2021). Porcine Epidemic Diarrhea Virus Induces Vero Cell Apoptosis via the p53-PUMA Signaling Pathway. Viruses, 13(7), 1218. https://doi.org/10.3390/v13071218