
Influence of Different Glycoproteins and of the Virion Core on SERINC5 Antiviral Activity

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid DNA
2.2. Cell Culture
2.3. Virus Production, and Transductions
2.4. Virion Purification and Western Blotting
2.5. Statistics
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deacon, N.J.; Tsykin, A.; Solomon, A.; Smith, K.; Ludford-Menting, M.; Hooker, D.J.; McPhee, D.A.; Greenway, A.L.; Ellett, A.; Chatfield, C.; et al. Genomic Structure of an Attenuated Quasi Species of HIV-1 from a Blood Transfusion Donor and Recipients. Science 1995, 270, 988–991. [Google Scholar] [CrossRef]
- Kestler, H.W., 3rd; Ringler, D.J.; Mori, K.; Panicali, D.L.; Sehgal, P.K.; Daniel, M.D.; Desrosiers, R.C. Importance of the Nef Gene for Maintenance of High Virus Loads and for Development of AIDS. Cell 1991, 65, 651–662. [Google Scholar] [CrossRef]
- Kirchhoff, F.; Greenough, T.C.; Brettler, D.B.; Sullivan, J.L.; Desrosiers, R.C. Absence of Intact Nef Sequences in a Long-Term Survivor with Nonprogressive HIV-1 Infection. N. Engl. J. Med. 1995, 332, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.; Shugars, D.C.; Swanstrom, R.; Garcia, J.V. Nef from Primary Isolates of Human Immunodeficiency Virus Type 1 Suppresses Surface CD4 Expression in Human and Mouse T Cells. J. Virol. 1993, 67, 4923–4931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, R.; Skowronski, J. CD4 down-Regulation by Nef Alleles Isolated from Human Immunodeficiency Virus Type 1-Infected Individuals. Proc. Natl. Acad. Sci. USA 1993, 90, 5549–5553. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.V.; Miller, A.D. Serine Phosphorylation-Independent Downregulation of Cell-Surface CD4 by Nef. Nature 1991, 350, 508–511. [Google Scholar] [CrossRef]
- Stove, V.; Van de Walle, I.; Naessens, E.; Coene, E.; Stove, C.; Plum, J.; Verhasselt, B. Human Immunodeficiency Virus Nef Induces Rapid Internalization of the T-Cell Coreceptor CD8alphabeta. J. Virol. 2005, 79, 11422–11433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, O.; Maréchal, V.; Le Gall, S.; Lemonnier, F.; Heard, J.M. Endocytosis of Major Histocompatibility Complex Class I Molecules Is Induced by the HIV-1 Nef Protein. Nat. Med. 1996, 2, 338–342. [Google Scholar] [CrossRef]
- Schindler, M.; Schmökel, J.; Specht, A.; Li, H.; Münch, J.; Khalid, M.; Sodora, D.L.; Hahn, B.H.; Silvestri, G.; Kirchhoff, F. Inefficient Nef-Mediated Downmodulation of CD3 and MHC-I Correlates with Loss of CD4+T Cells in Natural SIV Infection. PLoS Pathog. 2008, 4, e1000107. [Google Scholar] [CrossRef]
- Chowers, M.Y.; Spina, C.A.; Kwoh, T.J.; Fitch, N.J.; Richman, D.D.; Guatelli, J.C. Optimal Infectivity in Vitro of Human Immunodeficiency Virus Type 1 Requires an Intact Nef Gene. J. Virol. 1994, 68, 2906–2914. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, E.; Geleziunas, R.; Greene, W.C. Human Immunodeficiency Virus Type 1 Nef Functions at the Level of Virus Entry by Enhancing Cytoplasmic Delivery of Virions. J. Virol. 2001, 75, 2993–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobiume, M.; Lineberger, J.E.; Lundquist, C.A.; Miller, M.D.; Aiken, C. Nef Does Not Affect the Efficiency of Human Immunodeficiency Virus Type 1 Fusion with Target Cells. J. Virol. 2003, 77, 10645–10650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, E.M.; Nunez, R.; Hope, T.J. Disruption of the Actin Cytoskeleton Can Complement the Ability of Nef to Enhance Human Immunodeficiency Virus Type 1 Infectivity. J. Virol. 2004, 78, 5745–5755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavrois, M.; Neidleman, J.; Yonemoto, W.; Fenard, D.; Greene, W.C. HIV-1 Virion Fusion Assay: Uncoating Not Required and No Effect of Nef on Fusion. Virology 2004, 328, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, J.R.; Münk, C.; Guatelli, J.C. The Membrane-Proximal Tyrosine-Based Sorting Signal of Human Immunodeficiency Virus Type 1 gp41 Is Required for Optimal Viral Infectivity. J. Virol. 2004, 78, 1069–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, O.; Maréchal, V.; Danos, O.; Heard, J.M. Human Immunodeficiency Virus Type 1 Nef Increases the Efficiency of Reverse Transcription in the Infected Cell. J. Virol. 1995, 69, 4053–4059. [Google Scholar] [CrossRef] [Green Version]
- Aiken, C.; Trono, D. Nef Stimulates Human Immunodeficiency Virus Type 1 Proviral DNA Synthesis. J. Virol. 1995, 69, 5048–5056. [Google Scholar] [CrossRef] [Green Version]
- Rosa, A.; Chande, A.; Ziglio, S.; De Sanctis, V.; Bertorelli, R.; Goh, S.L.; McCauley, S.M.; Nowosielska, A.; Antonarakis, S.E.; Luban, J.; et al. HIV-1 Nef Promotes Infection by Excluding SERINC5 from Virion Incorporation. Nature 2015, 526, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Usami, Y.; Wu, Y.; Göttlinger, H.G. SERINC3 and SERINC5 Restrict HIV-1 Infectivity and Are Counteracted by Nef. Nature 2015, 526, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, C.; Marin, M.; Chande, A.; Pizzato, M.; Melikyan, G.B. SERINC5 Protein Inhibits HIV-1 Fusion Pore Formation by Promoting Functional Inactivation of Envelope Glycoproteins. J. Biol. Chem. 2017, 292, 6014–6026. [Google Scholar] [CrossRef] [Green Version]
- Trautz, B.; Pierini, V.; Wombacher, R.; Stolp, B.; Chase, A.J.; Pizzato, M.; Fackler, O.T. The Antagonism of HIV-1 Nef to SERINC5 Particle Infectivity Restriction Involves the Counteraction of Virion-Associated Pools of the Restriction Factor. J. Virol. 2016, 90, 10915–10927. [Google Scholar] [CrossRef] [Green Version]
- Heigele, A.; Kmiec, D.; Regensburger, K.; Langer, S.; Peiffer, L.; Stürzel, C.M.; Sauter, D.; Peeters, M.; Pizzato, M.; Learn, G.H.; et al. The Potency of Nef-Mediated SERINC5 Antagonism Correlates with the Prevalence of Primate Lentiviruses in the Wild. Cell Host Microbe 2016, 20, 381–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chande, A.; Cuccurullo, E.C.; Rosa, A.; Ziglio, S.; Carpenter, S.; Pizzato, M. S2 from Equine Infectious Anemia Virus Is an Infectivity Factor Which Counteracts the Retroviral Inhibitors SERINC5 and SERINC3. Proc. Natl. Acad. Sci. USA 2016, 113, 13197–13202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzato, M. MLV Glycosylated-Gag Is an Infectivity Factor That Rescues Nef-Deficient HIV-1. Proc. Natl. Acad. Sci. USA 2010, 107, 9364–9369. [Google Scholar] [CrossRef] [Green Version]
- Ahi, Y.S.; Zhang, S.; Thappeta, Y.; Denman, A.; Feizpour, A.; Gummuluru, S.; Reinhard, B.; Muriaux, D.; Fivash, M.J.; Rein, A. Functional Interplay Between Murine Leukemia Virus Glycogag, Serinc5, and Surface Glycoprotein Governs Virus Entry, with Opposite Effects on Gammaretroviral and Ebolavirus Glycoproteins. MBio 2016, 7, e01985-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, A.E.; Kiessling, V.; Pornillos, O.; White, J.M.; Ganser-Pornillos, B.K.; Tamm, L.K. HIV-Cell Membrane Fusion Intermediates Are Restricted by Serincs as Revealed by Cryo-Electron and TIRF Microscopy. J. Biol. Chem. 2020, 295, 15183–15195. [Google Scholar] [CrossRef]
- Lai, R.P.J.; Yan, J.; Heeney, J.; McClure, M.O.; Göttlinger, H.; Luban, J.; Pizzato, M. Nef Decreases HIV-1 Sensitivity to Neutralizing Antibodies That Target the Membrane-Proximal External Region of TMgp41. PLoS Pathog. 2011, 7, e1002442. [Google Scholar] [CrossRef] [PubMed]
- Usami, Y.; Göttlinger, H. HIV-1 Nef Responsiveness Is Determined by Env Variable Regions Involved in Trimer Association and Correlates with Neutralization Sensitivity. Cell Rep. 2013, 5, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Timilsina, U.; Umthong, S.; Lynch, B.; Stablewski, A.; Stavrou, S. SERINC5 Potently Restricts Retrovirus Infection In Vivo. MBio 2020, 11, e00588-20. [Google Scholar] [CrossRef] [PubMed]
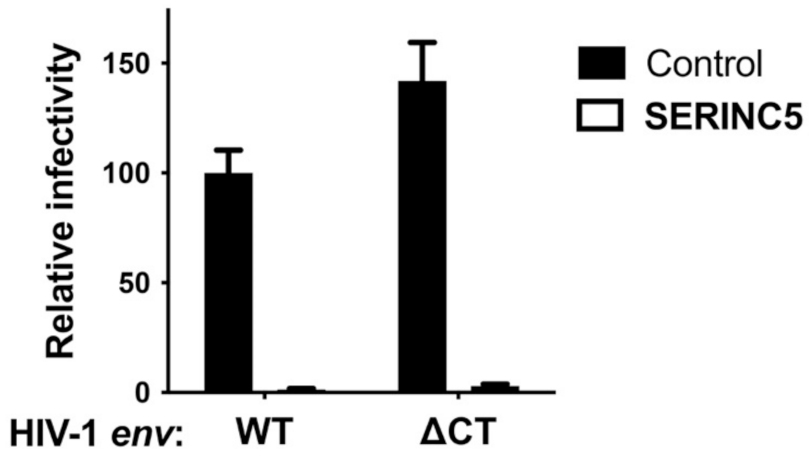
- Wilk, T.; Pfeiffer, T.; Bosch, V. Retained in Vitro Infectivity and Cytopathogenicity of HIV-1 despite Truncation of the C-Terminal Tail of the Env Gene Product. Virology 1992, 189, 167–177. [Google Scholar] [CrossRef]
- Wei, X.; Decker, J.M.; Liu, H.; Zhang, Z.; Arani, R.B.; Kilby, J.M.; Saag, M.S.; Wu, X.; Shaw, G.M.; Kappes, J.C. Emergence of Resistant Human Immunodeficiency Virus Type 1 in Patients Receiving Fusion Inhibitor (T-20) Monotherapy. Antimicrob. Agents Chemother. 2002, 46, 1896–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, W.E.; Stansell, E.; Kaiser, S.M.; Emerman, M.; Hunter, E. Identification of Postentry Restrictions to Mason-Pfizer Monkey Virus Infection in New World Monkey Cells. J. Virol. 2008, 82, 11140–11151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-Y.; La Russa, V.F.; Bao, L.; Kolls, J.; Schwarzenberger, P.; Reiser, J. Lentiviral Vectors for Sustained Transgene Expression in Human Bone Marrow-Derived Stromal Cells. Mol. Ther. 2002, 5, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Newman, R.M.; Hall, L.; Connole, M.; Chen, G.-L.; Sato, S.; Yuste, E.; Diehl, W.; Hunter, E.; Kaur, A.; Miller, G.M.; et al. Balancing Selection and the Evolution of Functional Polymorphism in Old World Monkey TRIM5alpha. Proc. Natl. Acad. Sci. USA 2006, 103, 19134–19139. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, M.; Emerman, M. Capsid Is a Dominant Determinant of Retrovirus Infectivity in Nondividing Cells. J. Virol. 2004, 78, 5670–5678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorny, M.K.; Gianakakos, V.; Sharpe, S.; Zolla-Pazner, S. Generation of Human Monoclonal Antibodies to Human Immunodeficiency Virus. Proc. Natl. Acad. Sci. USA 1989, 86, 1624–1628. [Google Scholar] [CrossRef] [Green Version]
- Chesebro, B.; Britt, W.; Evans, L.; Wehrly, K.; Nishio, J.; Cloyd, M. Characterization of Monoclonal Antibodies Reactive with Murine Leukemia Viruses: Use in Analysis of Strains of Friend MCF and Friend Ecotropic Murine Leukemia Virus. Virology 1983, 127, 134–148. [Google Scholar] [CrossRef]
- Liermann, M.; Steel, A.; Rosing, M.; Guttorp, P. Random Denominators and the Analysis of Ratio Data. Environ. Ecol. Stat. 2004, 11, 55–71. [Google Scholar] [CrossRef]
- Tedbury, P.R.; Freed, E.O. The role of matrix in HIV-1 envelope glycoprotein incorporation. Trends Microbiol. 2014, 22, 372–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
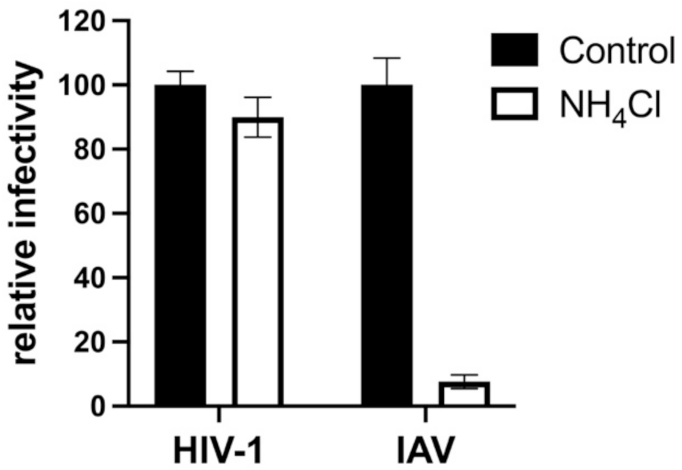
- Doms, R.W.; Gething, M.J.; Henneberry, J.; White, J.; Helenius, A. Variant Influenza Virus Hemagglutinin That Induces Fusion at Elevated pH. J. Virol. 1986, 57, 603–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccinotti, S.; Kirchhausen, T.; Whelan, S.P.J. Uptake of Rabies Virus into Epithelial Cells by Clathrin-Mediated Endocytosis Depends upon Actin. J. Virol. 2013, 87, 11637–11647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Simone, C.; Zandonatti, M.A.; Buchmeier, M.J. Acidic pH Triggers LCMV Membrane Fusion Activity and Conformational Change in the Glycoprotein Spike. Virology 1994, 198, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Stein, B.S.; Gowda, S.D.; Lifson, J.D.; Penhallow, R.C.; Bensch, K.G.; Engleman, E.G. pH-Independent HIV Entry into CD4-Positive T Cells via Virus Envelope Fusion to the Plasma Membrane. Cell 1987, 49, 659–668. [Google Scholar] [CrossRef]
- McClure, M.O.; Marsh, M.; Weiss, R.A. Human Immunodeficiency Virus Infection of CD4-Bearing Cells Occurs by a pH-Independent Mechanism. EMBO J. 1988, 7, 513–518. [Google Scholar] [CrossRef] [PubMed]
- McClure, M.O.; Sommerfelt, M.A.; Marsh, M.; Weiss, R.A. The pH Independence of Mammalian Retrovirus Infection. J. Gen. Virol. 1990, 71, 767–773. [Google Scholar] [CrossRef]
- Bissonnette, M.L.Z.; Connolly, S.A.; Young, D.F.; Randall, R.E.; Paterson, R.G.; Lamb, R.A. Analysis of the pH Requirement for Membrane Fusion of Different Isolates of the Paramyxovirus Parainfluenza Virus 5. J. Virol. 2006, 80, 3071–3077. [Google Scholar] [CrossRef] [Green Version]
- Chandran, K.; Sullivan, N.J.; Felbor, U.; Whelan, S.P.; Cunningham, J.M. Endosomal Proteolysis of the Ebola Virus Glycoprotein Is Necessary for Infection. Science 2005, 308, 1643–1645. [Google Scholar] [CrossRef] [Green Version]
- Gnirss, K.; Kühl, A.; Karsten, C.; Glowacka, I.; Bertram, S.; Kaup, F.; Hofmann, H.; Pöhlmann, S. Cathepsins B and L Activate Ebola but Not Marburg Virus Glycoproteins for Efficient Entry into Cell Lines and Macrophages Independent of TMPRSS2 Expression. Virology 2012, 424, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Carette, J.E.; Raaben, M.; Wong, A.C.; Herbert, A.S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A.I.; Kranzusch, P.J.; Griffin, A.M.; Ruthel, G.; et al. Ebola Virus Entry Requires the Cholesterol Transporter Niemann-Pick C1. Nature 2011, 477, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Côté, M.; Misasi, J.; Ren, T.; Bruchez, A.; Lee, K.; Filone, C.M.; Hensley, L.; Li, Q.; Ory, D.; Chandran, K.; et al. Small Molecule Inhibitors Reveal Niemann-Pick C1 Is Essential for Ebola Virus Infection. Nature 2011, 477, 344–348. [Google Scholar] [CrossRef]
- Diehl, W.E.; Lin, A.E.; Grubaugh, N.D.; Carvalho, L.M.; Kim, K.; Kyawe, P.P.; McCauley, S.M.; Donnard, E.; Kucukural, A.; McDonel, P.; et al. Ebola Virus Glycoprotein with Increased Infectivity Dominated the 2013-2016 Epidemic. Cell 2016, 167, 1088–1098.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanowicz, R.A.; McClure, C.P.; Sakuntabhai, A.; Sall, A.A.; Kobinger, G.; Müller, M.A.; Holmes, E.C.; Rey, F.A.; Simon-Loriere, E.; Ball, J.K. Human Adaptation of Ebola Virus during the West African Outbreak. Cell 2016, 167, 1079–1087.e5. [Google Scholar] [CrossRef] [Green Version]
- Le Tortorec, A.; Neil, S.J.D. Antagonism to and Intracellular Sequestration of Human Tetherin by the Human Immunodeficiency Virus Type 2 Envelope Glycoprotein. J. Virol. 2009, 83, 11966–11978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgenson, R.L.; Vogt, V.M.; Johnson, M.C. Foreign Glycoproteins Can Be Actively Recruited to Virus Assembly Sites during Pseudotyping. J. Virol. 2009, 83, 4060–4067. [Google Scholar] [CrossRef] [Green Version]
- Freed, E.O.; Martin, M.A. Domains of the Human Immunodeficiency Virus Type 1 Matrix and gp41 Cytoplasmic Tail Required for Envelope Incorporation into Virions. J. Virol. 1996, 70, 341–351. [Google Scholar] [CrossRef] [Green Version]
- West, J.T.; Weldon, S.K.; Wyss, S.; Lin, X.; Yu, Q.; Thali, M.; Hunter, E. Mutation of the Dominant Endocytosis Motif in Human Immunodeficiency Virus Type 1 gp41 Can Complement Matrix Mutations without Increasing Env Incorporation. J. Virol. 2002, 76, 3338–3349. [Google Scholar] [CrossRef] [Green Version]
- Pandori, M.W.; Fitch, N.J.; Craig, H.M.; Richman, D.D.; Spina, C.A.; Guatelli, J.C. Producer-Cell Modification of Human Immunodeficiency Virus Type 1: Nef Is a Virion Protein. J. Virol. 1996, 70, 4283–4290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiken, C. Pseudotyping Human Immunodeficiency Virus Type 1 (HIV-1) by the Glycoprotein of Vesicular Stomatitis Virus Targets HIV-1 Entry to an Endocytic Pathway and Suppresses Both the Requirement for Nef and the Sensitivity to Cyclosporin A. J. Virol. 1997, 71, 5871–5877. [Google Scholar] [CrossRef] [Green Version]
- Chazal, N.; Singer, G.; Aiken, C.; Hammarskjöld, M.L.; Rekosh, D. Human Immunodeficiency Virus Type 1 Particles Pseudotyped with Envelope Proteins That Fuse at Low pH No Longer Require Nef for Optimal Infectivity. J. Virol. 2001, 75, 4014–4018. [Google Scholar] [CrossRef] [Green Version]
- Chertova, E.; Bess, J.W., Jr.; Crise, B.J.; Sowder, R.C., II; Schaden, T.M.; Hilburn, J.M.; Hoxie, J.A.; Benveniste, R.E.; Lifson, J.D.; Henderson, L.E.; et al. Envelope Glycoprotein Incorporation, Not Shedding of Surface Envelope Glycoprotein (gp120/SU), Is the Primary Determinant of SU Content of Purified Human Immunodeficiency Virus Type 1 and Simian Immunodeficiency Virus. J. Virol. 2002, 76, 5315–5325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandenberg, O.F.; Magnus, C.; Rusert, P.; Regoes, R.R.; Trkola, A. Different Infectivity of HIV-1 Strains Is Linked to Number of Envelope Trimers Required for Entry. PLoS Pathog. 2015, 11, e1004595. [Google Scholar] [CrossRef] [PubMed]
- Trautz, B.; Wiedemann, H.; Uumlchtenborg, C.L.; Pierini, V.; Kranich, J.; Glass, B.A.; Kr Aumlusslich, H.-G.; Brocker, T.; Pizzato, M.; Ruggieri, A.; et al. The Host-Cell Restriction Factor SERINC5 Restricts HIV-1 Infectivity without Altering the Lipid Composition and Organization of Viral Particles. J. Biol. Chem. 2017, 292, 13702–13713. [Google Scholar] [CrossRef] [Green Version]
- Chan, R.; Uchil, P.D.; Jin, J.; Shui, G.; Ott, D.E.; Mothes, W.; Wenk, M.R. Retroviruses Human Immunodeficiency Virus and Murine Leukemia Virus Are Enriched in Phosphoinositides. J. Virol. 2008, 82, 11228–11238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, H.C.; Mason, M.M. A New Virus in a Spontaneous Mammary Tumor of a Rhesus Monkey. Cancer Res. 1970, 30, 2081–2086. [Google Scholar] [PubMed]
- Sfakianos, J.N.; LaCasse, R.A.; Hunter, E. The M-PMV Cytoplasmic Targeting-Retention Signal Directs Nascent Gag Polypeptides to a Pericentriolar Region of the Cell. Traffic 2003, 4, 660–670. [Google Scholar] [CrossRef]
- Ivanova, P.T.; Myers, D.S.; Milne, S.B.; McClaren, J.L.; Thomas, P.G.; Brown, H.A. Lipid composition of viral envelope of three strains of influenza virus—Not all viruses are created equal. ACS Infect. Dis. 2015, 1, 399–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
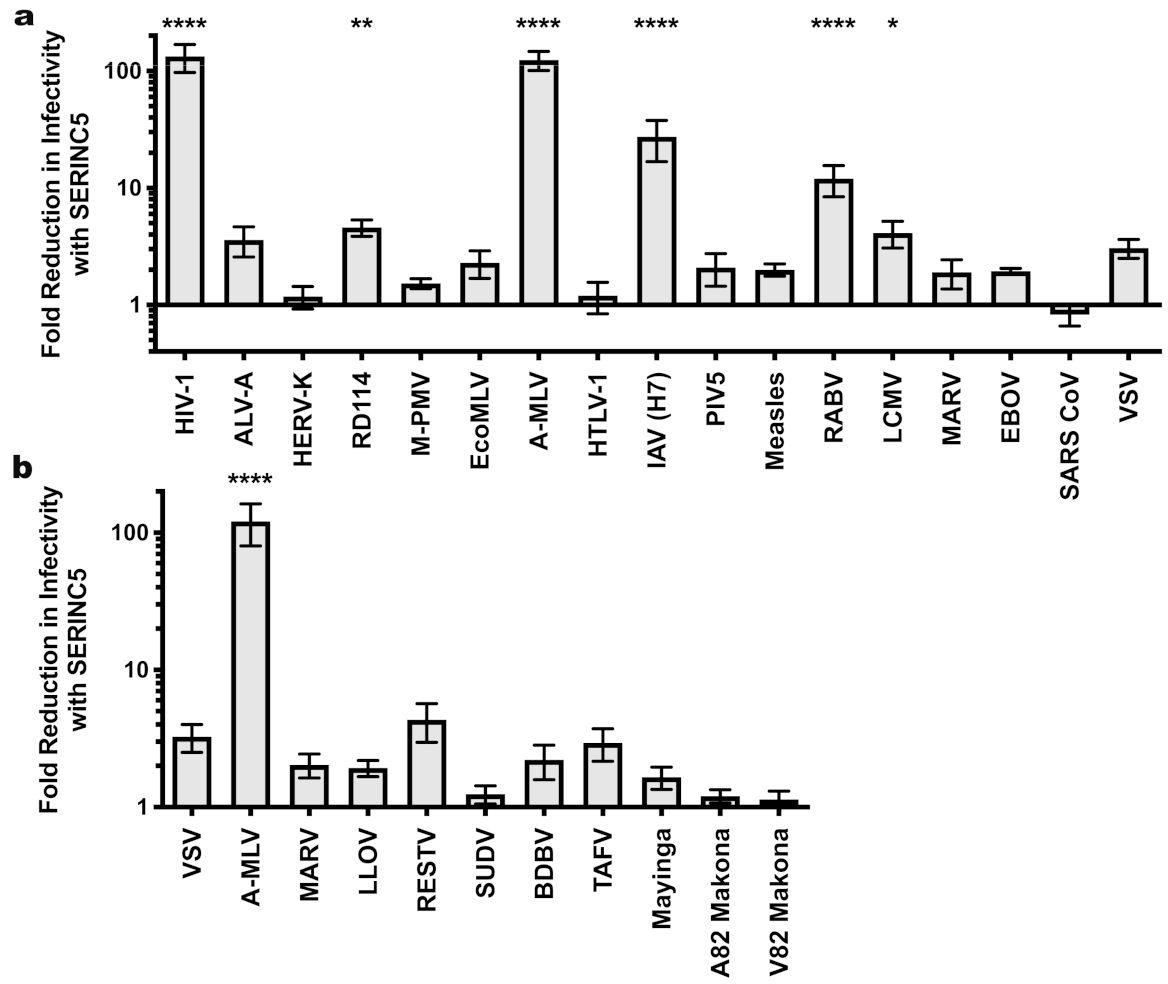
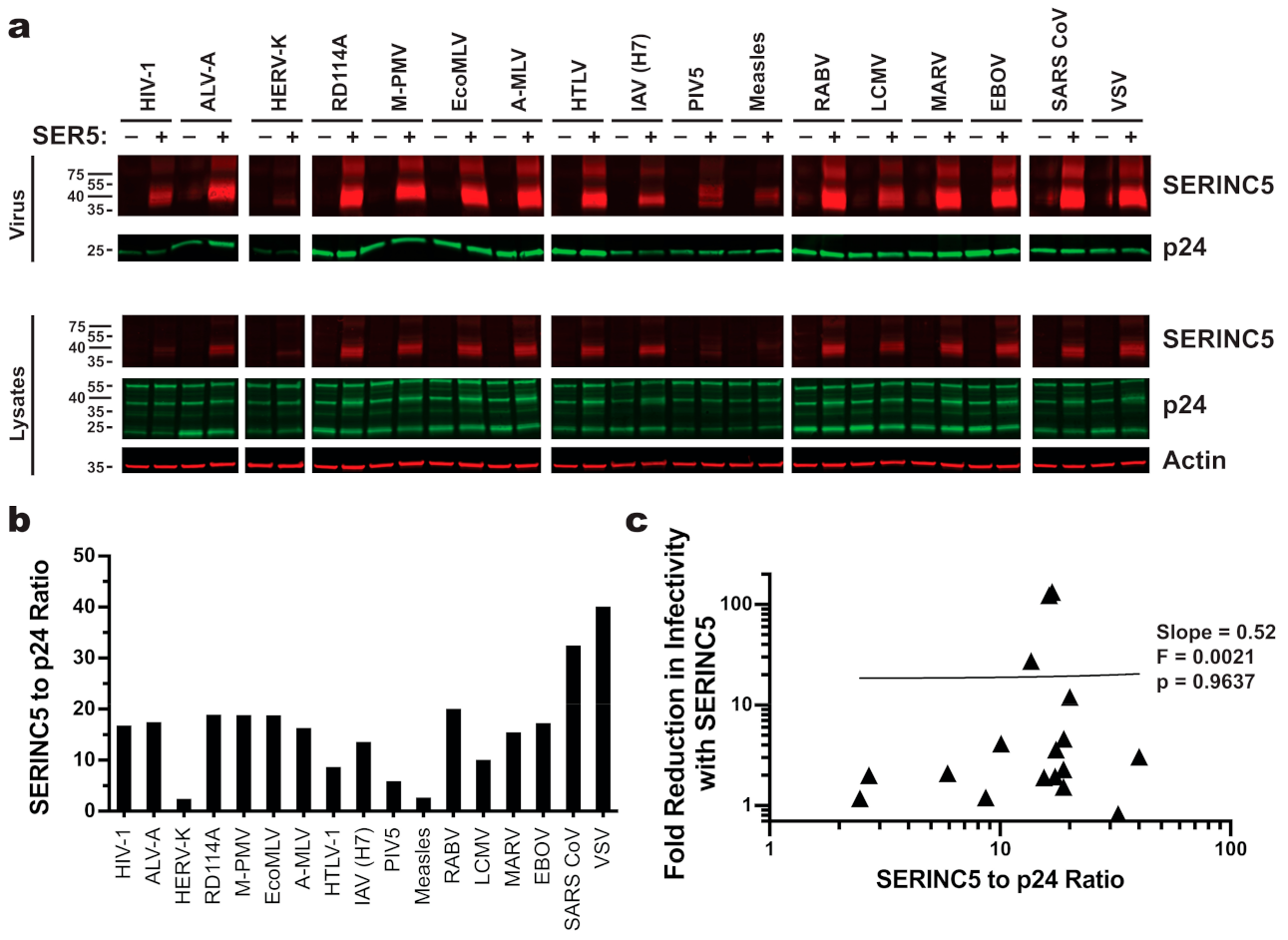
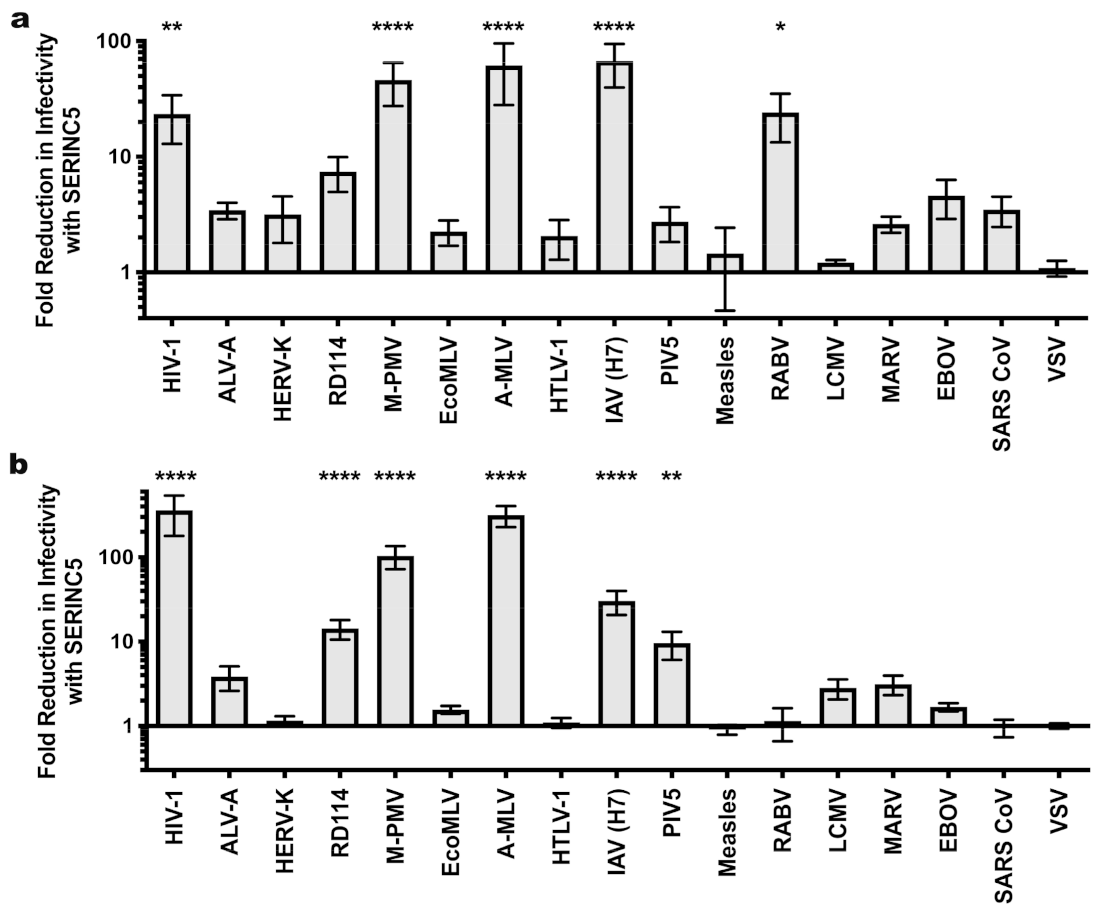
| HIV-1 Core | MLV Core | M-PMV Core | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glycoprotein | Fold Restriction a | SEM | n b | Avg. Inf. (TU/mL) c | Fold Restriction a | SEM | n b | Avg. Inf. (TU/mL) c | Fold Restriction a | SEM | n b | Avg. Inf. (TU/mL) c |
| HIV-1 | 132.9 **** | 35.6 | 3 | 1.50 × 105 | 23.5 ** | 10.6 | 3 | 7.50 × 103 | 360.1 **** | 181.1 | 3 | 2.63 × 104 |
| ALV-A | 3.6 | 1.0 | 4 | 9.30 × 106 | 3.4 | 0.6 | 4 | 1.61 × 105 | 3.9 | 1.2 | 4 | 4.32 × 105 |
| HERV-K | 1.2 | 0.3 | 3 | 4.96 × 102 | 3.2 | 1.4 | 5 | 2.43 × 102 | 1.2 | 0.2 | 4 | 2.10 × 103 |
| RD114 | 4.6 ** | 0.7 | 4 | 1.19 × 107 | 7.4 | 2.5 | 5 | 1.04 × 106 | 24.8 **** | 10.9 | 6 | 9.40 × 105 |
| M-PMV | 1.5 | 0.2 | 3 | 6.50 × 104 | 46.3 **** | 18.7 | 6 | 1.76 × 105 | 104.2 **** | 31.9 | 8 | 3.72 × 105 |
| EcoMLV | 1.7 | 0.6 | 3 | 1.30 × 107 | 2.3 | 0.6 | 4 | 6.19 × 105 | 3.2 | 1.6 | 5 | 1.17 × 106 |
| A-MLV | 123.8 **** | 22.7 | 12 | 1.68 × 106 | 61.7 **** | 33.7 | 7 | 4.68 × 105 | 315.8 **** | 87.9 | 12 | 9.80 × 105 |
| HTLV-1 | 1.2 | 0.4 | 3 | 1.66 × 103 | 2.1 | 0.8 | 5 | 8.06 × 102 | 1.1 | 0.2 | 4 | 2.60 × 103 |
| Flu (H7) | 27.3 **** | 10.6 | 5 | 6.61 × 106 | 67.1 **** | 27.2 | 4 | 5.90 × 104 | 31.2 **** | 9.6 | 8 | 1.01 × 106 |
| PIV5 | 2.1 | 0.6 | 6 | 4.06 × 105 | 2.7 | 0.9 | 3 | 2.04 × 104 | 9.6 ** | 3.5 | 7 | 2.21 × 105 |
| Measles | 2.0 | 0.2 | 4 | 5.49 × 104 | 1.5 | 1.0 | 3 | 9.98 × 101 | 0.9 | 0.1 | 4 | 9.01 × 103 |
| RABV | 12.0 **** | 3.6 | 3 | 3.28 × 106 | 24.2 * | 10.8 | 6 | 2.46 × 104 | 1.1 | 0.5 | 4 | 3.91 × 103 |
| LCMV | 4.1 * | 1.1 | 3 | 9.76 × 106 | 1.2 | 0.1 | 3 | 1.19 × 105 | 5.1 | 2.4 | 7 | 4.65 × 105 |
| MARV | 1.9 | 0.5 | 3 | 9.08 × 106 | 2.6 | 0.4 | 3 | 7.19 × 104 | 3.1 | 0.8 | 4 | 2.67 × 105 |
| EBOV | 1.9 | 0.1 | 3 | 3.64 × 105 | 4.6 | 1.7 | 3 | 2.41 × 103 | 1.7 | 0.2 | 4 | 2.50 × 104 |
| SARS CoV | 0.9 | 0.2 | 5 | 1.80 × 104 | 3.5 | 1.0 | 4 | 1.42 × 103 | 1.0 | 0.2 | 4 | 5.11 × 103 |
| VSV | 3.1 | 0.6 | 11 | 7.86 × 107 | 1.1 | 0.2 | 6 | 2.38 × 106 | 1.0 | 0.1 | 11 | 3.68 × 106 |
| HIV-1 Core vs. | MLV Core vs. | ||
|---|---|---|---|
| Glycoprotein | MLV Core | M-PMV Core | M-PMV Core |
| M-PMV | p = 0.0012 a | p < 0.0001 | N.S. b |
| PIV5 | N.S. | p = 0.0355 | N.S. |
| Rabies | N.S. | p = 0.0118 | N.S. |
| RD114 | N.S. | N.S. | N.S. |
| LCMV | N.S. | N.S. | N.S. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diehl, W.E.; Guney, M.H.; Vanzo, T.; Kyawe, P.P.; White, J.M.; Pizzato, M.; Luban, J. Influence of Different Glycoproteins and of the Virion Core on SERINC5 Antiviral Activity. Viruses 2021, 13, 1279. https://doi.org/10.3390/v13071279
Diehl WE, Guney MH, Vanzo T, Kyawe PP, White JM, Pizzato M, Luban J. Influence of Different Glycoproteins and of the Virion Core on SERINC5 Antiviral Activity. Viruses. 2021; 13(7):1279. https://doi.org/10.3390/v13071279
Chicago/Turabian StyleDiehl, William E., Mehmet H. Guney, Teresa Vanzo, Pyae P. Kyawe, Judith M. White, Massimo Pizzato, and Jeremy Luban. 2021. "Influence of Different Glycoproteins and of the Virion Core on SERINC5 Antiviral Activity" Viruses 13, no. 7: 1279. https://doi.org/10.3390/v13071279
APA StyleDiehl, W. E., Guney, M. H., Vanzo, T., Kyawe, P. P., White, J. M., Pizzato, M., & Luban, J. (2021). Influence of Different Glycoproteins and of the Virion Core on SERINC5 Antiviral Activity. Viruses, 13(7), 1279. https://doi.org/10.3390/v13071279