
Antigenic Characterization of ORF2 and ORF3 Proteins of Hepatitis E Virus (HEV)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction and Amplification of HEV Viral RNA
2.2. Production of Recombinant Antigens
2.3. Production and Characterization of Monoclonal Antibodies
2.4. Swine Sera
2.5. Western-Blotting
2.6. Indirect ELISA
2.7. MAb Competitive Binding ELISA
2.8. Sandwich ELISA
2.9. Trapping ELISA
2.10. Sera Competitive ELISA
3. Results
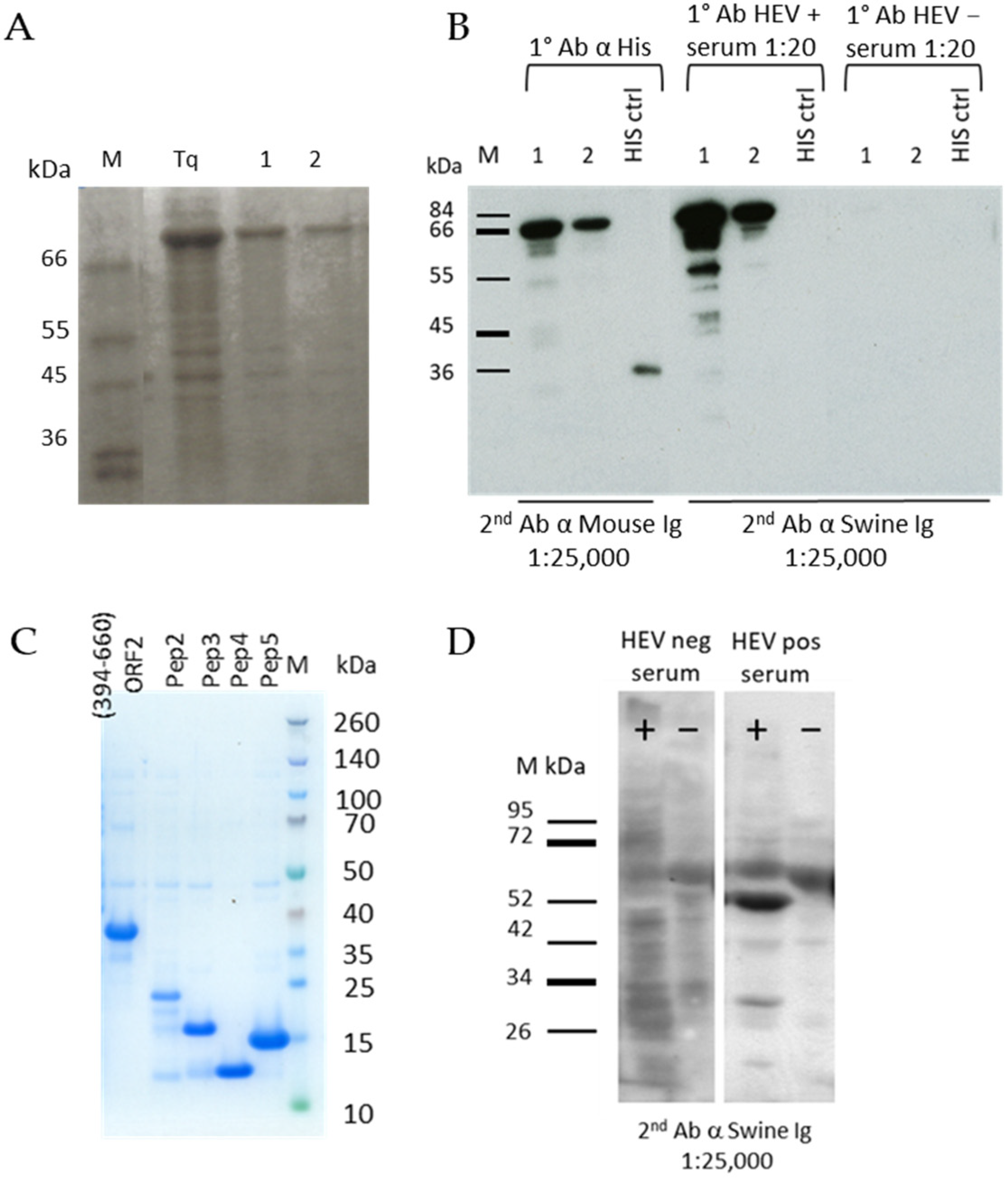
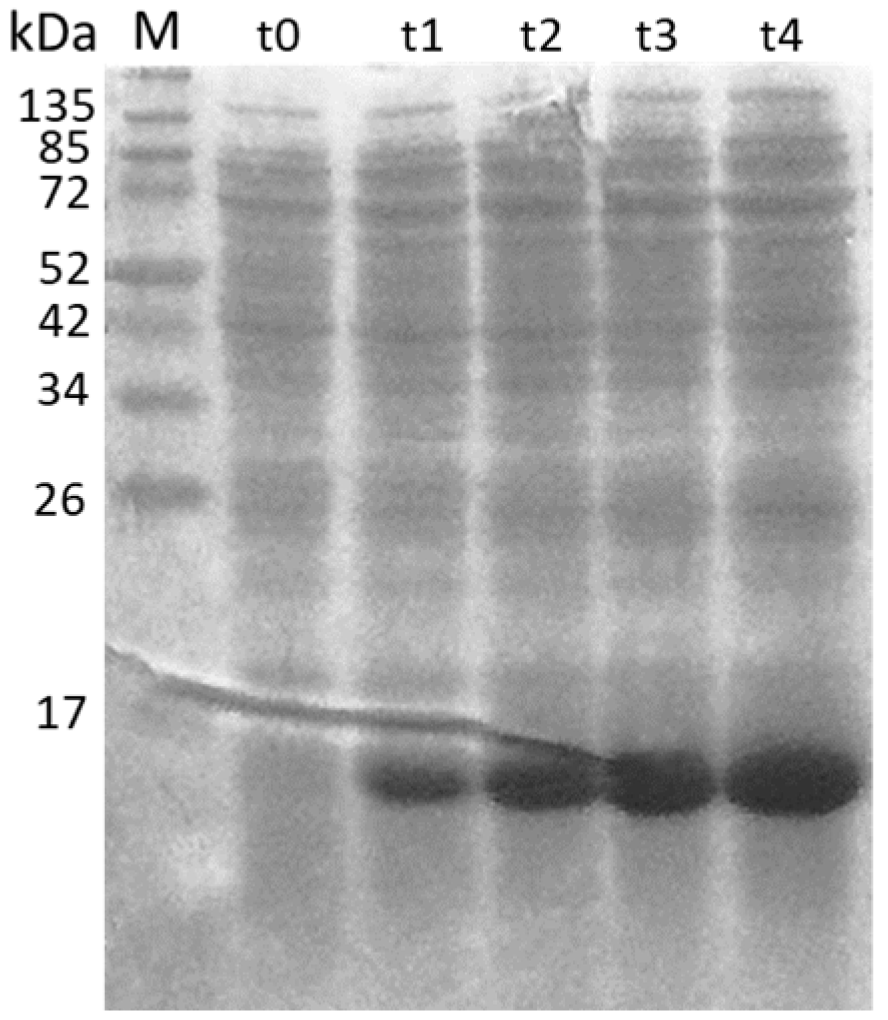
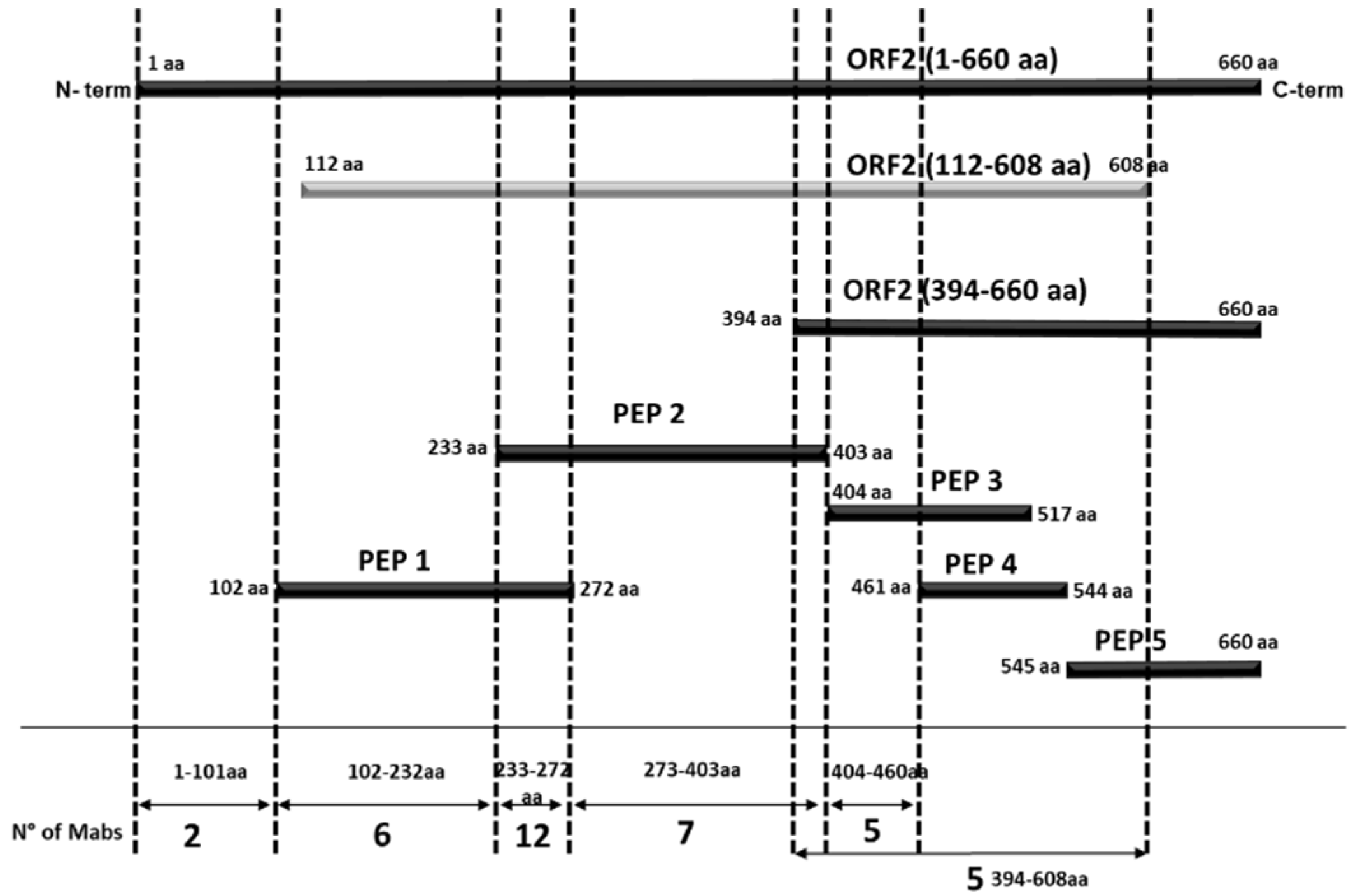
3.1. Production of Recombinant Proteins and Peptides
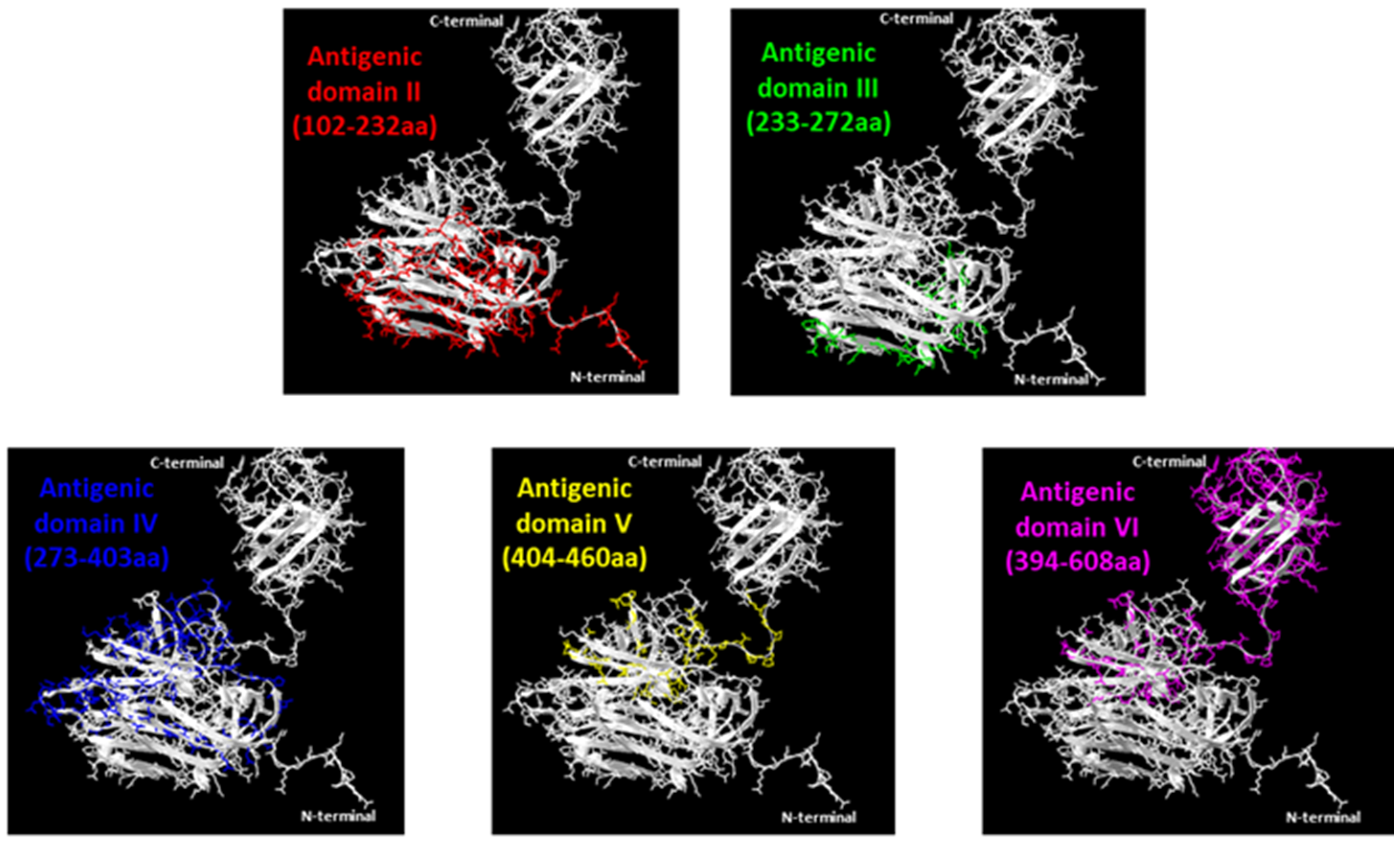
3.2. Evaluation of ORF2 Antigenicity with MAbs
3.3. Epitopes Differentiation by MAbs Reciprocal Competition
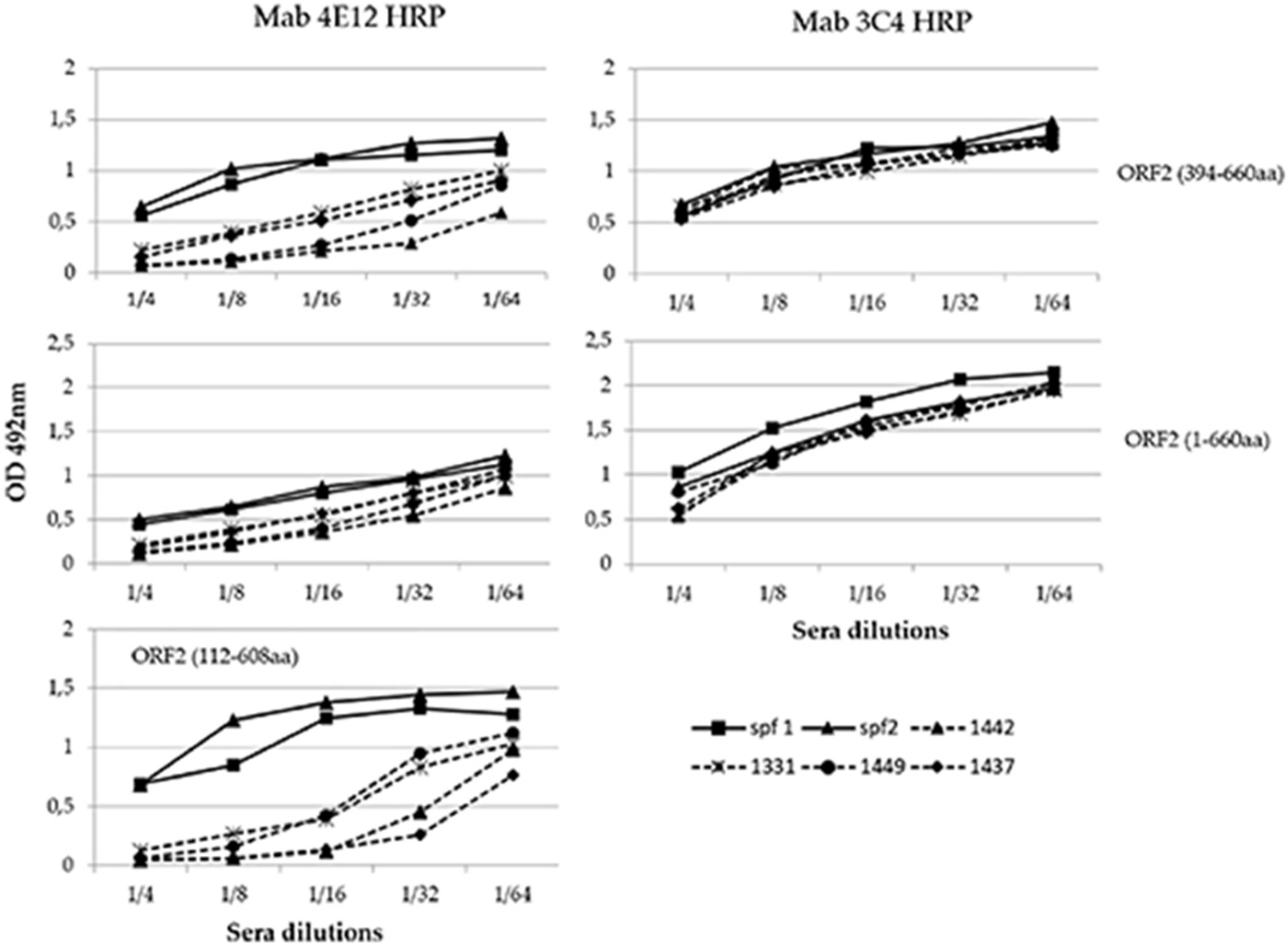
3.4. Evaluation of the Conformational Properties of the Different ORF2 Recombinant Antigens Using Sandwich ELISA with MAbs
3.5. Analysis of Immunogenicity of the Different ORF2 Recombinant Antigens Using Trapping ELISA
3.6. Evaluation of MAbs Target Epitopes Immunogenicity Using Competitive ELISA
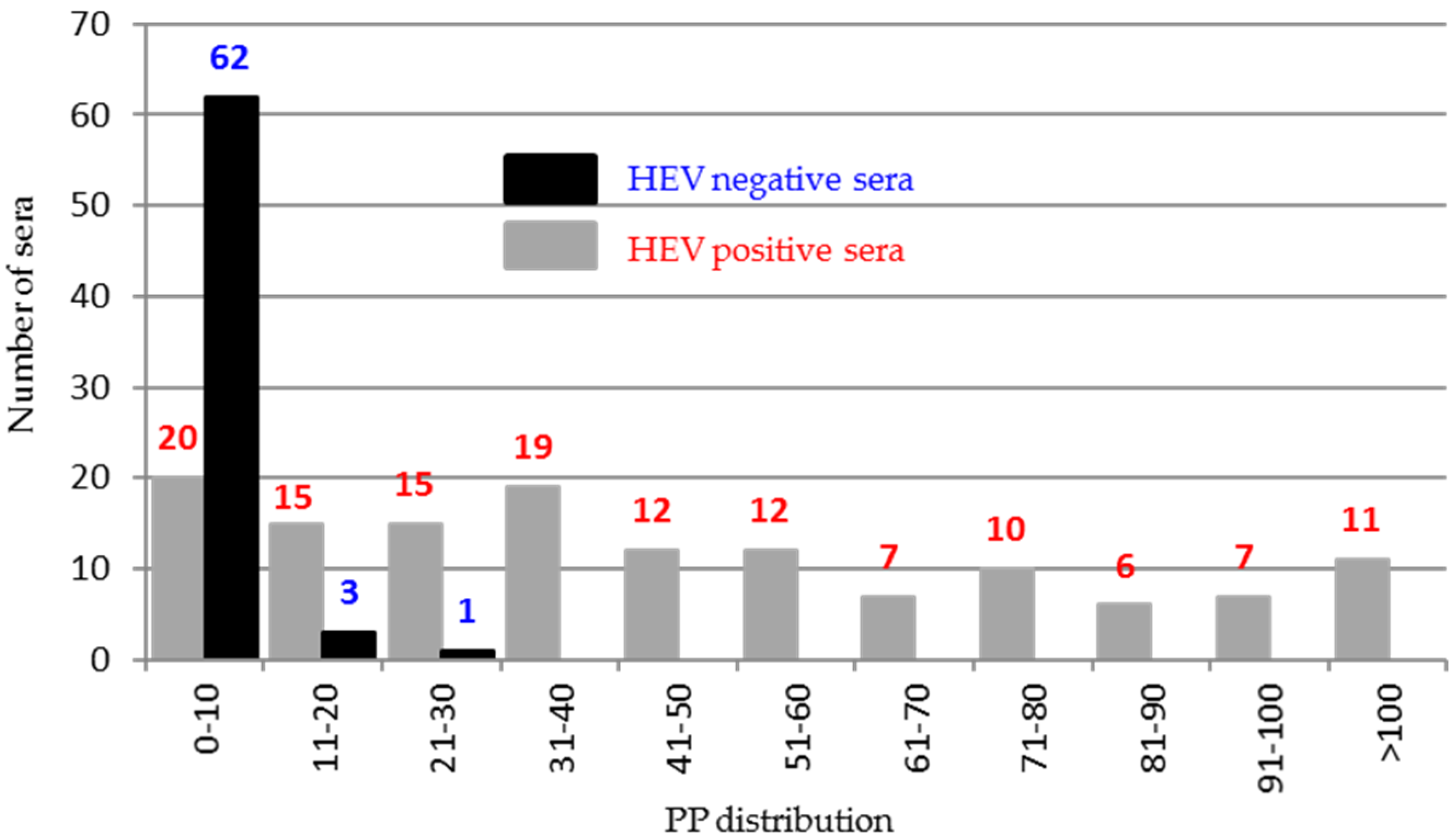
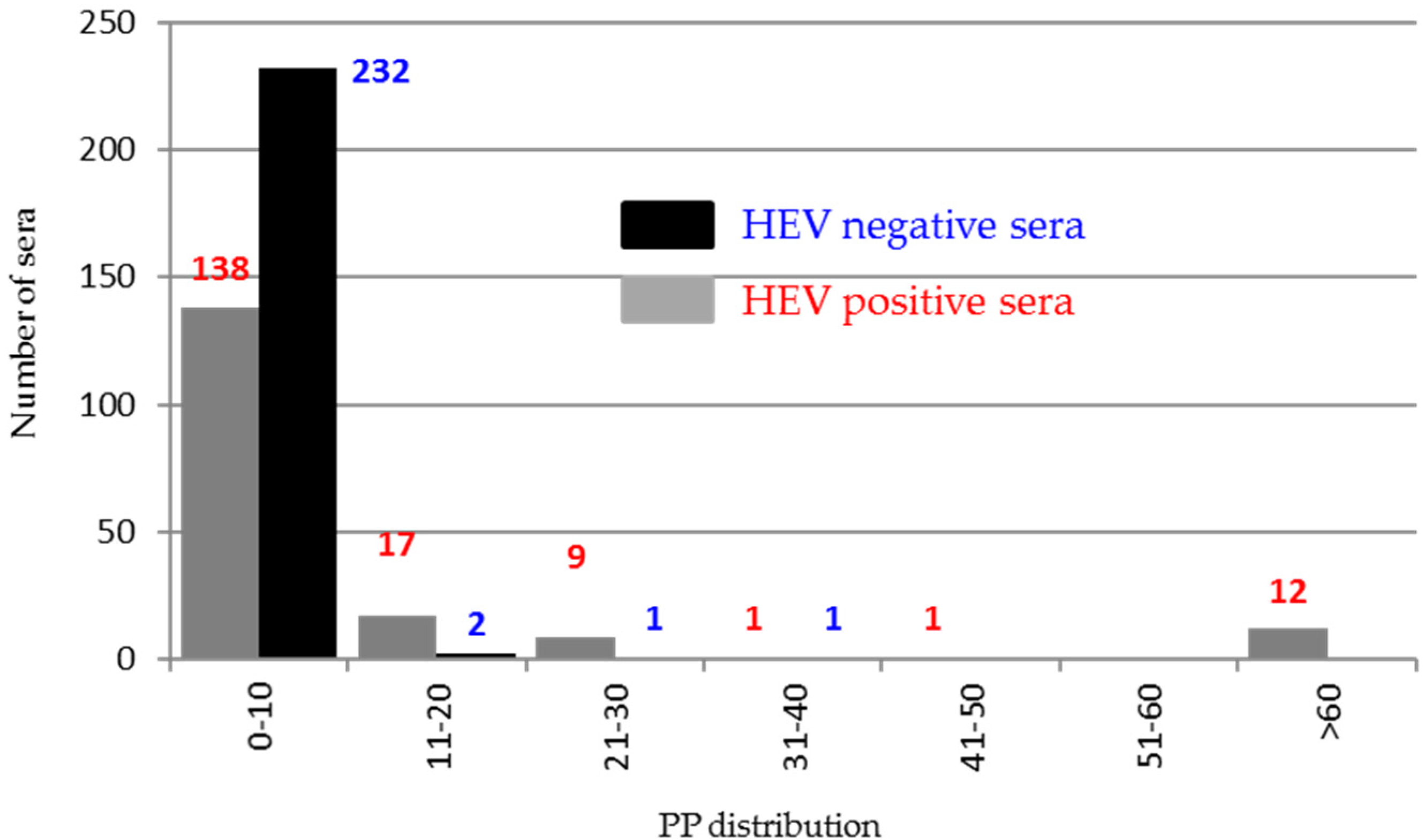
3.7. Evaluation of Immunogenicity of ORF2 Peptides and ORF3 Using Field Sera
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scobie, L.; Dalton, H.R. Hepatitis E: Source and route of infection, clinical manifestations and new developments. J. Viral Hepat. 2013, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Teli, M.R.; Skidmore, S.; Sofi, M.A.; Khuroo, M.I. Incidence and severity of viral hepatitis in pregnancy. Am. J. Med. 1981, 70, 252–255. [Google Scholar] [CrossRef]
- Donnelly, M.C.; Scobie, L.; Crossan, C.L.; Dalton, H.; Hayes, P.C.; Simpson, K.J. Review article: Hepatitis E-a concise review of virology, epidemiology, clinical presentation and therapy. Aliment. Pharmacol. Ther. 2017, 46, 126–141. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.J. Hepatitis E virus: Animal reservoirs and zoonotic risk. Veter Microbiol. 2010, 140, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic Infection With Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on hepatitis E virology: Implications for clinical practice. J. Hepatol. 2016, 65, 200–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khudyakov, Y.E.; Khudyakova, N.S.; Fields, H.A.; Jue, D.; Starling, C.; Favorov, M.O.; Krawczynski, K.; Polish, L.; Mast, E.; Margolis, H. Epitope mapping in proteins of hepatitis E virus. Virology 1993, 194, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Khudyakov Yu, E.; Favorov, M.O.; Jue, D.L.; Hine, T.K.; Fields, H.A. Immunodominant antigenic regions in a structural protein of the hepatitis E virus. Virology 1994, 198, 390–393. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Purcell, R.H.; Emerson, S.U. A truncated ORF2 protein contains the most immunogenic site on ORF2: Antibody responses to non-vaccine sequences following challenge of vaccinated and non-vaccinated macaques with hepatitis E virus. Vaccine 2005, 23, 3157–3165. [Google Scholar] [CrossRef]
- Li, S.W.; Zhao, Q.; Wu, T.; Chen, S.; Zhang, J.; Xia, N.S. The development of a recombinant hepatitis E vaccine HEV 239. Hum. Vaccines Immunother. 2015, 11, 908–914. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Torresi, J.; Locarnini, S.A.; Zhuang, H.; Zhu, W.; Guo, X.; Anderson, D.A. Amino-terminal epitopes are exposed when full-length open reading frame 2 of hepatitis E virus is expressed in Escherichia coli, but carboxy-terminal epitopes are masked. J. Med. Virol. 1997, 52, 289–300. [Google Scholar] [CrossRef]
- Riddell, M.A.; Li, F.; Anderson, D.A. Identification of immunodominant and conformational epitopes in the capsid protein of hepatitis E virus by using monoclonal antibodies. J. Virol. 2000, 74, 8011–8017. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Wang, J.C.; Li, T.C.; Yasutomi, Y.; Lara, J.; Khudyakov, Y.; Schofield, D.; Emerson, S.U.; Purcell, R.H.; Takeda, N.; et al. Spatial configuration of hepatitis E virus antigenic domain. J. Virol. 2011, 85, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Riddell, M.A.; Seow, H.F.; Takeda, N.; Miyamura, T.; Anderson, D.A. Recombinant subunit ORF2.1 antigen and induction of antibody against immunodominant epitopes in the hepatitis E virus capsid protein. J. Med. Virol. 2000, 60, 379–386. [Google Scholar] [CrossRef]
- de Oya, N.J.; Escribano-Romero, E.; Blázquez, A.B.; Lorenzo, M.; Martín-Acebes, M.A.; Blasco, R.; Saiz, J.C. Characterization of hepatitis E virus recombinant ORF2 proteins expressed by vaccinia viruses. J. Virol. 2012, 86, 7880–7886. [Google Scholar] [CrossRef] [Green Version]
- Osterman, A.; Vizoso Pinto, M.G.; Haase, R.; Nitschko, H.; Jäger, S.; Sander, M.; Motz, M.; Mohn, U.; Baiker, A. Systematic screening for novel, serologically reactive Hepatitis E Virus epitopes. Virol. J. 2012, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Yang, C.; Gu, Y.; Song, C.; Zhang, X.; Wang, Y.; Zhang, J.; Hew, C.L.; Li, S.; Xia, N.; et al. Structural basis for the neutralization and genotype specificity of hepatitis E virus. Proc. Natl. Acad. Sci. USA 2011, 108, 10266–10271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Dai, X.; Chang, J.C.; Lopareva, E.; Pillot, J.; Fields, H.A.; Khudyakov, Y.E. Identification and characterization of the neutralization epitope(s) of the hepatitis E virus. Virology 2001, 288, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, D.J.; Glamann, J.; Emerson, S.U.; Purcell, R.H. Identification by phage display and characterization of two neutralizing chimpanzee monoclonal antibodies to the hepatitis E virus capsid protein. J. Virol. 2000, 74, 5548–5555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Song, X.; Harrison, T.J.; Li, R.; Huang, G.; Zhang, H.; Kong, W.; Wang, Y. Immunogenicity and efficacy of a bacterially expressed HEV ORF3 peptide, assessed by experimental infection of primates. Arch. Virol. 2009, 154, 1641–1648. [Google Scholar] [CrossRef] [PubMed]
- Pezzoni, G.; Caminiti, A.; Stercoli, L.; Grazioli, S.; Galletti, G.; Santi, A.; Tamba, M.; Brocchi, E. Comparison of three in-house ELISAs for the detection of hepatitis E virus infection in pigs under field conditions. J. Virol. Methods 2014, 207, 95–103. [Google Scholar] [CrossRef]
- Tijssen, P. Preparation of enzyme-antibody or other enzyme-macromolecule conjugates. In Laboratory Techniques in Biochemistry and Molecular Biology; Burdon, R.H., van Knipperberg, P.H., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 221–277. [Google Scholar]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- He, J.; Kuschner, R.A.; Dewar, V.; Voet, P.; Asher, L.V.; Vaughn, D.W. Characterization of monoclonal antibodies to hepatitis E virus (HEV) capsid protein and identification of binding activity. J. Biomed. Sci. 2007, 14, 555–563. [Google Scholar] [CrossRef]
- Zhao, M.; Li, X.J.; Tang, Z.M.; Yang, F.; Wang, S.L.; Cai, W.; Zhang, K.; Xia, N.S.; Zheng, Z.Z. A Comprehensive Study of Neutralizing Antigenic Sites on the Hepatitis E Virus (HEV) Capsid by Constructing, Clustering, and Characterizing a Tool Box. J. Biol. Chem. 2015, 290, 19910–19922. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Bing, Z.; Guan, S.; Zhang, Z.; Wang, X. Development of new hepatitis E vaccines. Hum. Vaccines Immunother. 2018, 14, 2254–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Liu, Q.; Yu, R.; Li, Z.; Li, J.; Zhu, H.; Wu, X.; Tan, F.; Wang, J.; Tang, X. Screening of specific diagnostic peptides of swine hepatitis E virus. Virol. J. 2009, 6, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Liu, B.; Sun, Y.; Li, H.; Du, T.; Nan, Y.; Hiscox, J.A.; Zhou, E.M.; Zhao, Q. Characterization of Three Novel Linear Neutralizing B-Cell Epitopes in the Capsid Protein of Swine Hepatitis E Virus. J. Virol. 2018, 92, e00251-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.C.; Takeda, N.; Miyamura, T.; Matsuura, Y.; Wang, J.C.; Engvall, H.; Hammar, L.; Xing, L.; Cheng, R.H. Essential elements of the capsid protein for self-assembly into empty virus-like particles of hepatitis E virus. J. Virol. 2005, 79, 12999–13006. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tang, X.; Seetharaman, J.; Yang, C.; Gu, Y.; Zhang, J.; Du, H.; Shih, J.W.; Hew, C.L.; Sivaraman, J.; et al. Dimerization of hepatitis E virus capsid protein E2s domain is essential for virus-host interaction. PLoS Pathog. 2009, 5, e1000537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gu, Y.; Ge, S.X.; Li, S.W.; He, Z.Q.; Huang, G.Y.; Zhuang, H.; Ng, M.H.; Xia, N.S. Analysis of hepatitis E virus neutralization sites using monoclonal antibodies directed against a virus capsid protein. Vaccine 2005, 23, 2881–2892. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Mori, Y.; Miyazaki, N.; Cheng, R.H.; Yoshimura, M.; Unno, H.; Shima, R.; Moriishi, K.; Tsukihara, T.; Li, T.C.; et al. Biological and immunological characteristics of hepatitis E virus-like particles based on the crystal structure. Proc. Natl. Acad. Sci. USA 2009, 106, 12986–12991. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MAbs | Isotype | WB ORF2 | ELISA ORF2 (1–660 aa) | IF ORF2 (112–608 aa) | ELISA | Domain | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF2 (394–660 aa) | Peptide 1 (102–272 aa) | Peptide 2 (233–403 aa) | Peptide 3 (404–517 aa) | Peptide 4 (461–544 aa) | Peptide 5 (545–660 aa) | ||||||
| 1A2 | IgG1 | n.d. | + | - | - | - | - | - | - | - | I (1–101 aa) |
| 3E9 | IgG1 | + | + | - | - | - | - | - | - | - | |
| 5G1 | IgG1/IgG3/IgG2 | + | + | - | - | + | - | - | - | - | II (102–232 aa) |
| 5G5 | IgG1/IgG3/IgG2 | + | + | + | - | + | - | - | - | - | |
| 5H9 | IgG1 | + | + | + | - | + | - | - | - | - | |
| 1G3 | IgG1 | + | + | + | - | + | - | - | - | - | |
| 2B3 | IgG3 | + | + | + | - | + | - | - | - | - | |
| 4G12 | IgG1 | + | + | + | - | + | - | - | - | - | |
| 2C7 | IgG2a | + | + | + | - | + | + | - | - | - | III (233–272 aa) |
| 1E2 | IgG1 | + | + | + | - | + | + | - | - | - | |
| 5F8 | IgG3/IgM | + | + | + | - | + | + | - | - | - | |
| 4E6 | IgG2a | + | + | + | - | + | + | - | - | - | |
| 2G10 | IgG3 | + | + | + | - | + | + | - | - | - | |
| 2F6 | IgG3 | + | + | + | - | + | + | - | - | - | |
| 2D7 | IgG1 | + | + | + | - | + | + | - | - | - | |
| 5F7 | IgG3 | + | + | weak | - | + | + | - | - | - | |
| 2C6 | IgG2a | + | + | weak | - | + | + | - | - | - | |
| 2D6 | IgG2a/IgM | + | + | weak | - | + | + | - | - | - | |
| 3H7 | IgG2a | + | + | + | - | + | + | - | - | - | |
| 1D4 | n.d. | weak | + | + | - | + | + | - | - | - | |
| 2G8 | IgG1 | n.d. | + | + | - | - | + | - | - | - | IV (273–403 aa) |
| 1D10 | IgG1 | weak | + | + | - | - | + | - | - | - | |
| 1G4 | IgG1 | + | + | + | - | - | + | - | - | - | |
| 5E9 | IgG1 | + | + | + | - | - | + | - | - | - | |
| 5C10 | IgG1 | + | + | + | - | - | + | - | - | - | |
| 1H9 | IgG1 | n.d. | + | + | - | - | + | - | - | - | |
| 3E7 | IgG1 | weak | + | + | - | - | + | - | - | - | |
| 3D9 | IgG1/IgG3/IgG2b | + | + | + | + | - | - | + | - | - | V (404–460 aa) |
| 3C3 | IgG2b | + | + | + | + | - | - | + | - | - | |
| 3C4 | IgG2b | + | + | + | + | - | - | + | - | - | |
| 2C12 | IgG2a | + | + | + | + | - | - | + | - | - | |
| 3A8 | IgG1 | n.d. | + | + | + | - | - | + | - | - | |
| 4E12 | IgG1 | n.d. | + | + | + | - | - | - | - | - | VI (394–608 aa) |
| 5D10 | IgG1 | + | + | + | + | - | - | - | - | - | |
| 4G6 | IgG1 | n.d. | + | + | + | - | - | - | - | - | |
| 5G9 | IgG1 | + | + | + | + | - | - | - | - | - | |
| 5A8 | IgG1 | + | + | + | + | - | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pezzoni, G.; Stercoli, L.; Pegoiani, E.; Brocchi, E. Antigenic Characterization of ORF2 and ORF3 Proteins of Hepatitis E Virus (HEV). Viruses 2021, 13, 1385. https://doi.org/10.3390/v13071385
Pezzoni G, Stercoli L, Pegoiani E, Brocchi E. Antigenic Characterization of ORF2 and ORF3 Proteins of Hepatitis E Virus (HEV). Viruses. 2021; 13(7):1385. https://doi.org/10.3390/v13071385
Chicago/Turabian StylePezzoni, Giulia, Lidia Stercoli, Eleonora Pegoiani, and Emiliana Brocchi. 2021. "Antigenic Characterization of ORF2 and ORF3 Proteins of Hepatitis E Virus (HEV)" Viruses 13, no. 7: 1385. https://doi.org/10.3390/v13071385
APA StylePezzoni, G., Stercoli, L., Pegoiani, E., & Brocchi, E. (2021). Antigenic Characterization of ORF2 and ORF3 Proteins of Hepatitis E Virus (HEV). Viruses, 13(7), 1385. https://doi.org/10.3390/v13071385