
Targeting JAK/STAT Signaling Antagonizes Resistance to Oncolytic Reovirus Therapy Driven by Prior Infection with HTLV-1 in Models of T-Cell Lymphoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Chemicals and Reagents
2.3. Quantification of Drug-Induced Cytotoxicity
2.4. Transmission Electron Microscopy
2.5. Immunoblotting and Antibodies
2.6. RNA Isolation and Expression Arrays
2.7. Quantitative Real-Time Polymerase Chain Reaction
2.8. Statistical Analyses
3. Results
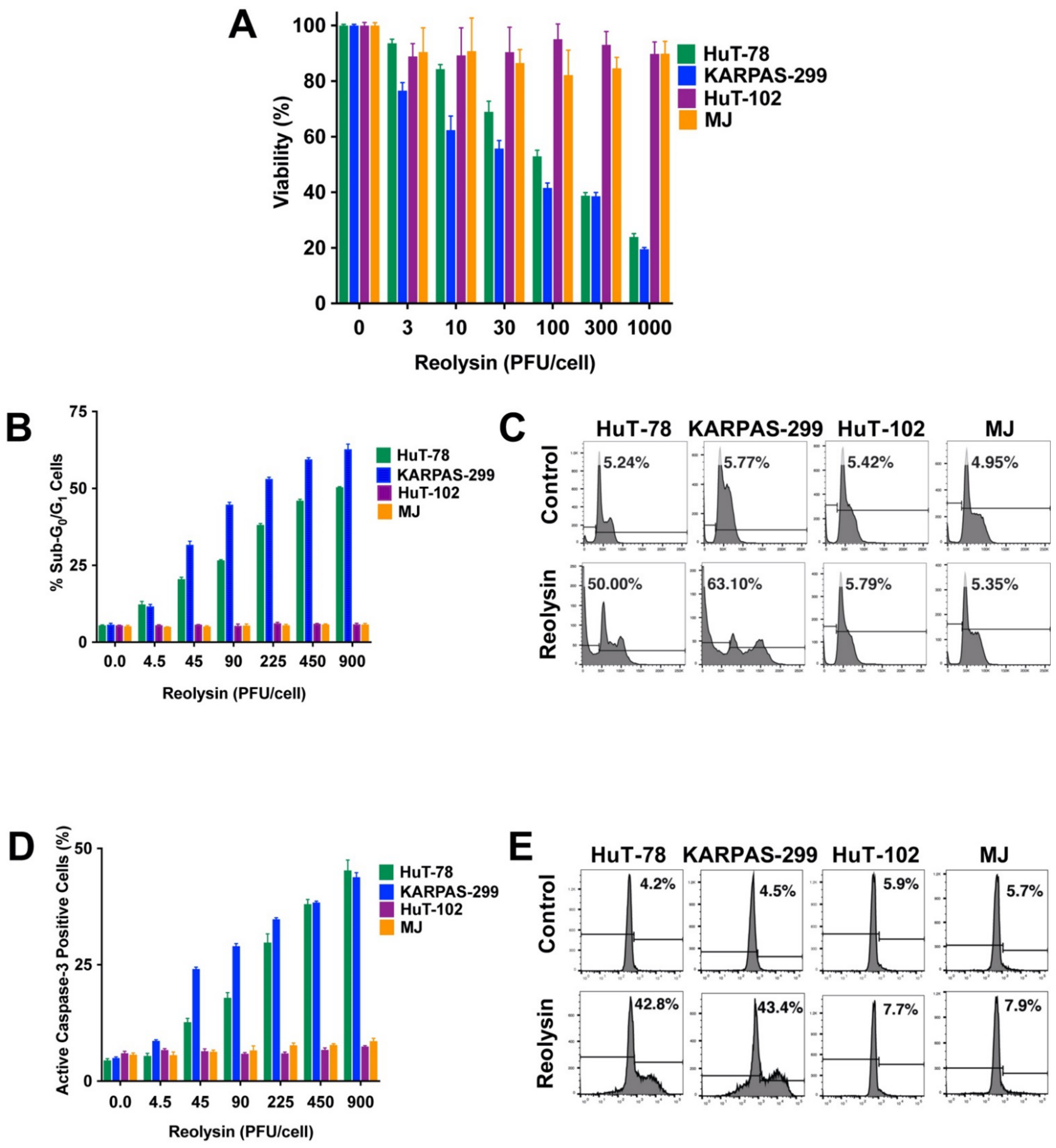
3.1. Reolysin Decreases TCL Cell Viability and Induces Apoptosis in HuT-78 and KARPAS-299 Cells
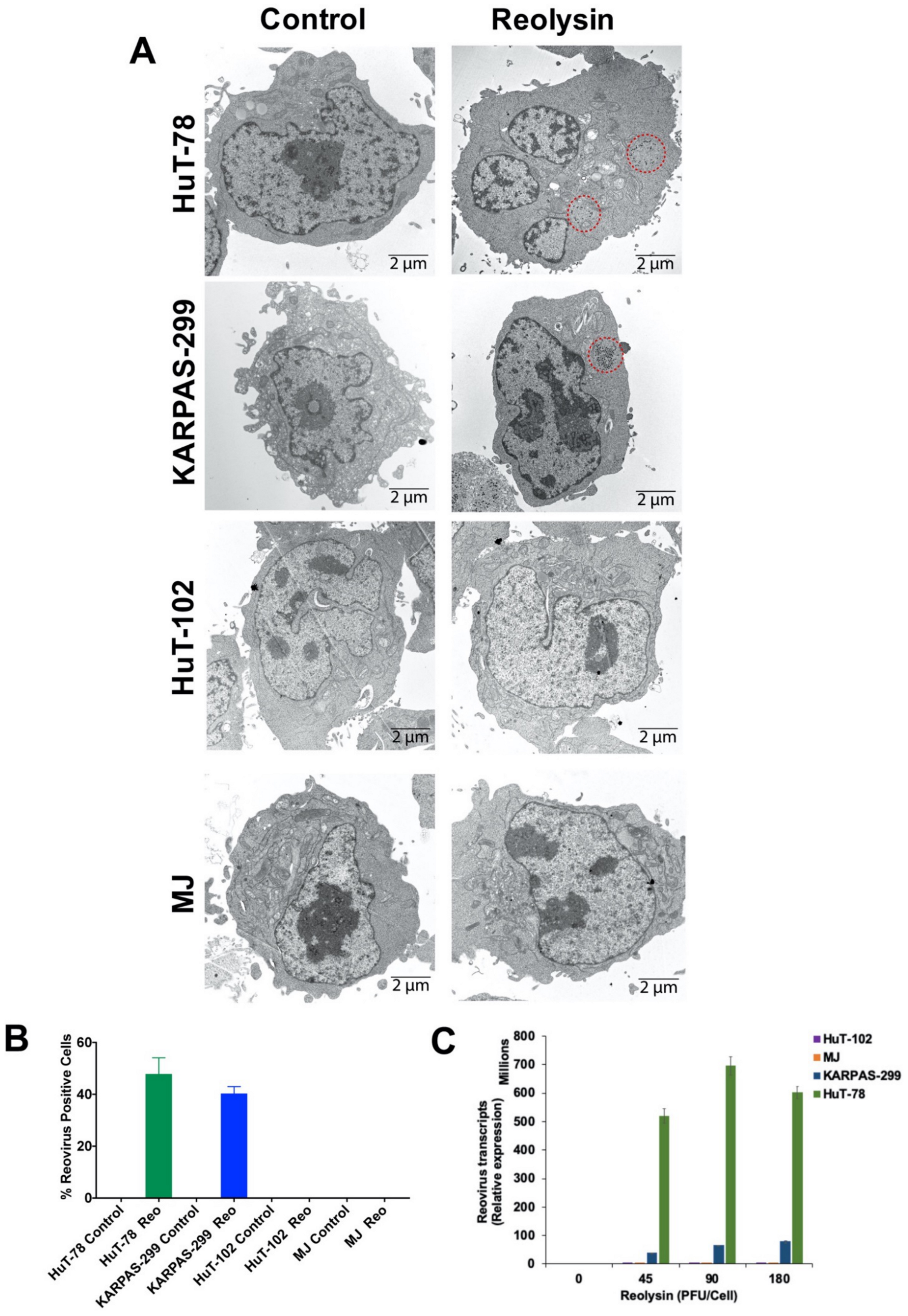
3.2. Oncolytic Reovirus Displays Productive Infection in HuT-78 and KARPAS-299 TCL Cells
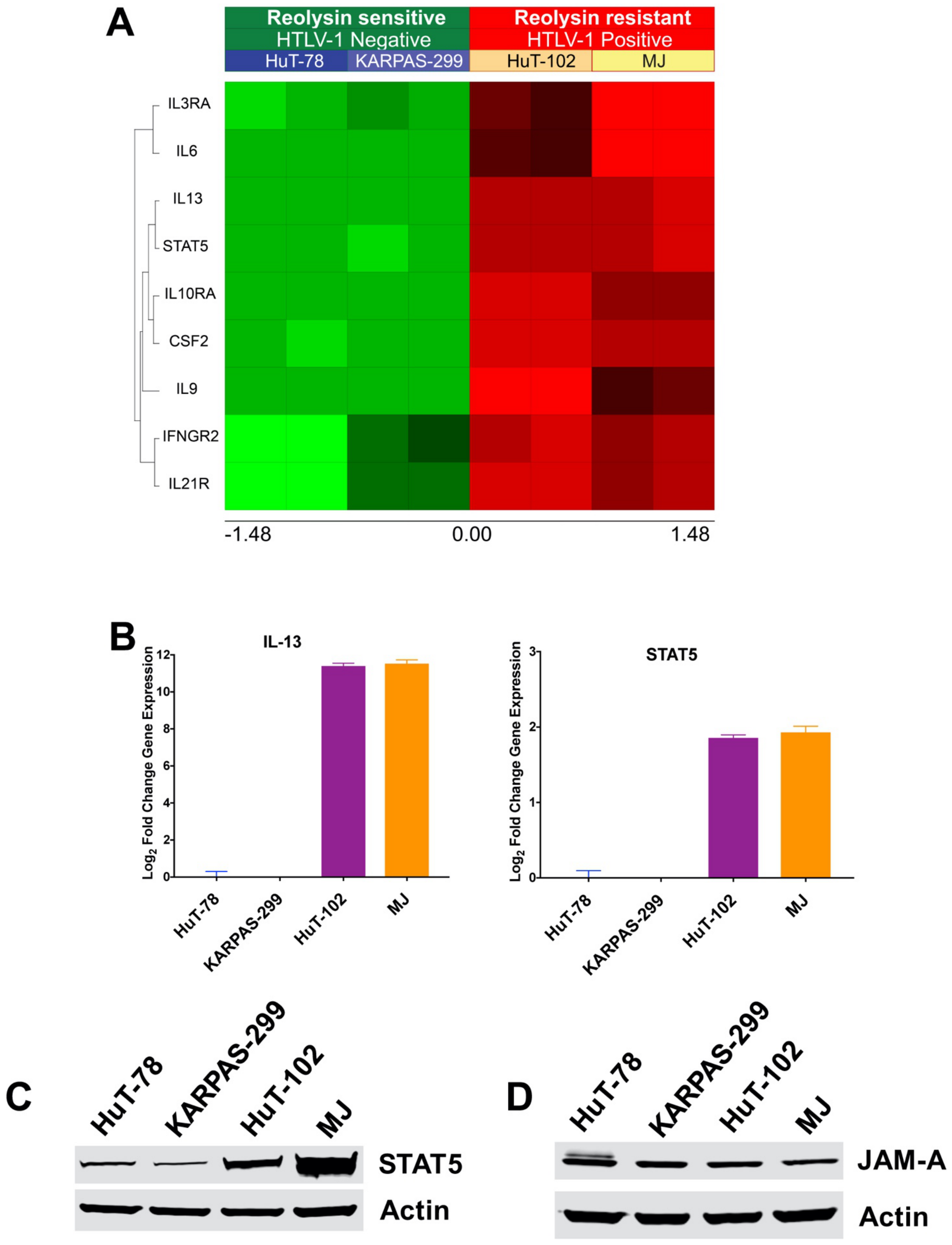
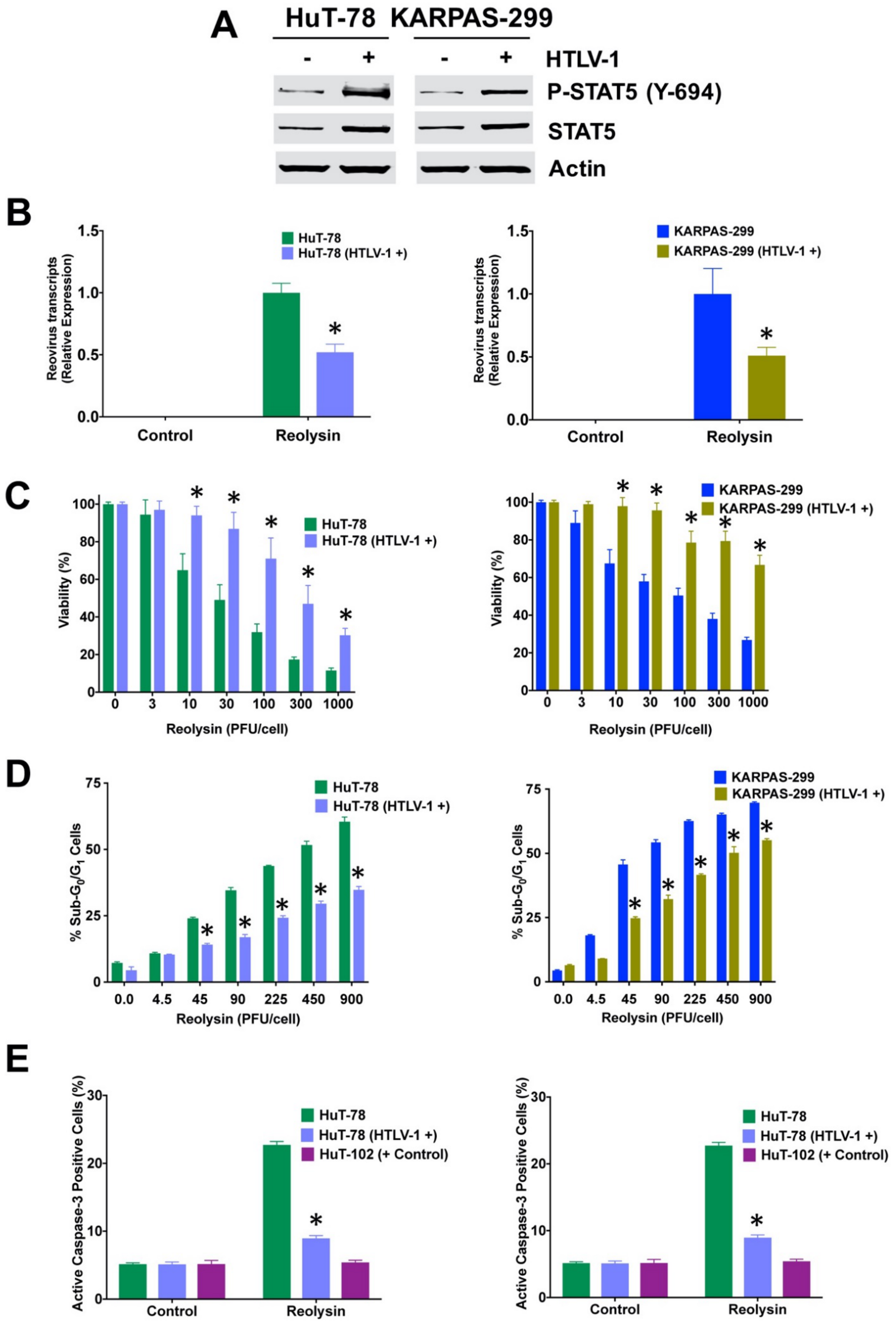
3.3. HTLV-1-Positive TCL Cells Display Significantly Upregulated IL-13 and STAT5 Expression
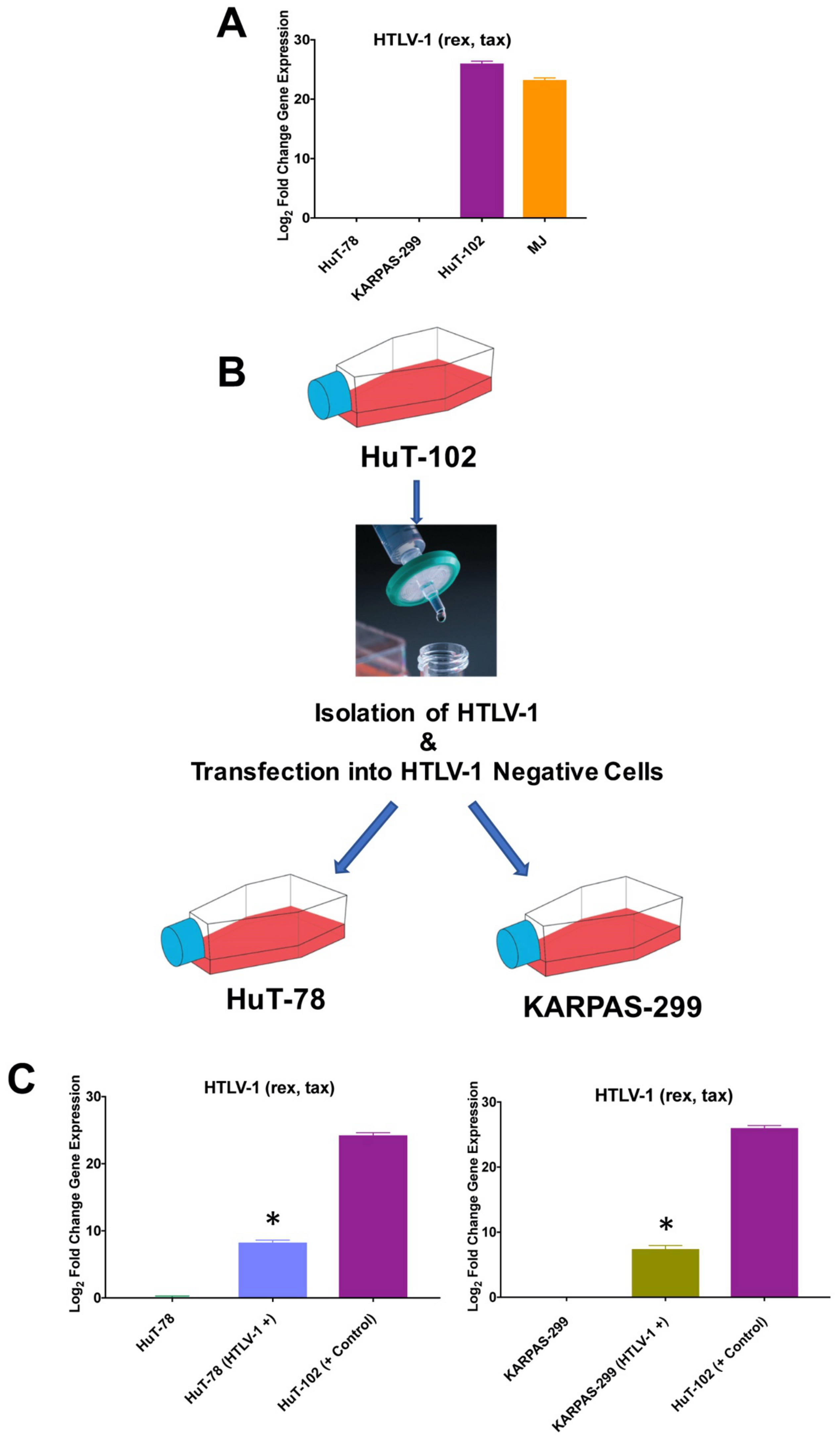
3.4. HuT-78 and KARPAS-299 TCL Cells Are Susceptbile to HTLV-1 Infection
3.5. Infection with HTLV-1 Promotes Resistance to Oncolytic Reovirus Therapy
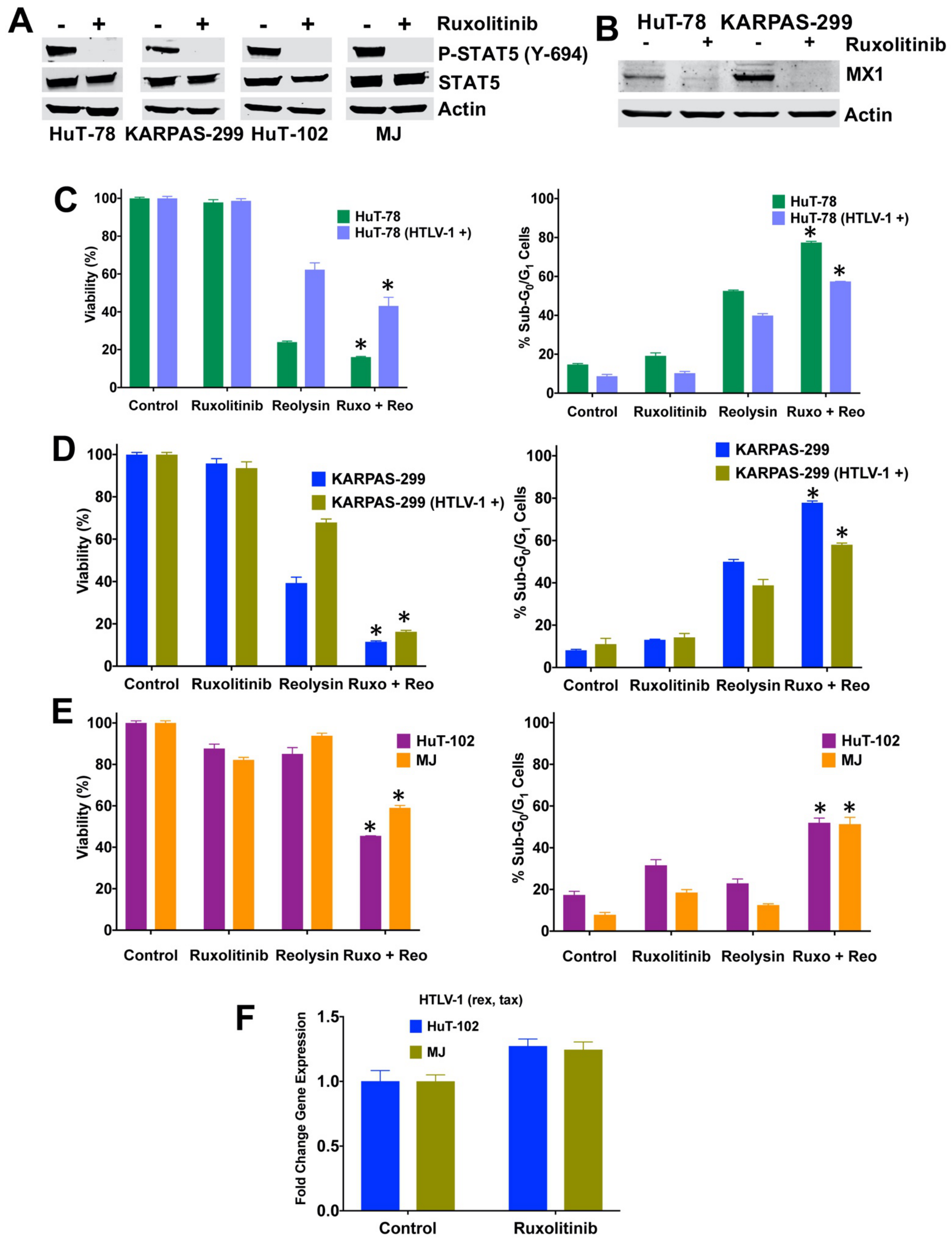
3.6. Ruxolitinib Decreases STAT5 and MX1 Expression and Significantly Enhances the Anti-TCL Activity of Reolysin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El Hajj, H.; Tsukasaki, K.; Cheminant, M.; Bazarbachi, A.; Watanabe, T.; Hermine, O. Novel Treatments of Adult t Cell Leukemia Lymphoma. Front. Microbiol. 2020, 11, 1062. [Google Scholar] [CrossRef]
- Islam, S.; Vick, E.; Huber, B.; Morales, C.; Spier, C.; Cooke, L.; Weterings, E.; Mahadevan, D. Co-targeting Aurora Kinase with Pd-l1 and Pi3k Abrogates Immune Checkpoint MEDIATED Proliferation in Peripheral t-Cell Lymphoma: A Novel Therapeutic Strategy. Oncotarget 2017, 8, 100326–100338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, P.V.; Cassar, O.; Gessain, A. Molecular Epidemiology, Genetic Variability and Evolution of Htlv-1 with Special Emphasis on African Genotypes. Retrovirology 2019, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangham, C.R.M. Human t Cell Leukemia Virus Type 1: Persistence and Pathogenesis. Annu. Rev. Immunol. 2018, 36, 43–71. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M.; Nakano, K.; Miyake, A.; Yamochi, T.; Kagami, Y.; Tsutsumi, A.; Matsuda, Y.; Sato-Otsubo, A.; Muto, S.; Utsunomiya, A.; et al. Polycomb-Mediated Loss of Mir-31 Activates Nik-Dependent Nf-Kappab Pathway in Adult t Cell Leukemia and Other Cancers. Cancer Cell 2012, 21, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Tsukasaki, K.; Tobinai, K. Biology and Treatment of Htlv-1 Associated t-Cell Lymphomas. Best Pract. Res. Clin. Haematol. 2013, 26, 3–14. [Google Scholar] [CrossRef]
- Chung, H.K.; Young, H.A.; Goon, P.K.; Heidecker, G.; Princler, G.L.; Shimozato, O.; Taylor, G.P.; Bangham, C.R.; Derse, D. Activation of Interleukin-13 Expression in t Cells from Htlv-1-Infected Individuals and in Chronically Infected Cell Lines. Blood 2003, 102, 4130–4136. [Google Scholar] [CrossRef] [Green Version]
- Waldele, K.; Schneider, G.; Ruckes, T.; Grassmann, R. Interleukin-13 Overexpression by Tax Transactivation: A Potential autocrine Stimulus in Human t-Cell Leukemia virus-Infected Lymphocytes. J. Virol. 2004, 78, 6081–6090. [Google Scholar] [CrossRef] [Green Version]
- Leonard, W.J.; O’Shea, J.J. Jaks and stats: Biological implications. Annu. Rev. Immunol. 1998, 16, 293–322. [Google Scholar] [CrossRef] [Green Version]
- Calton, C.M.; Kelly, K.R.; Anwer, F.; Carew, J.S.; Nawrocki, S.T. Oncolytic Viruses for Multiple Myeloma Therapy. Cancers 2018, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Muller, L.; Berkeley, R.; Barr, T.; Ilett, E.; Errington-Mais, F. Past, Present and Future of Oncolytic Reovirus. Cancers 2020, 12, 3219. [Google Scholar] [CrossRef]
- Hingorani, P.; Zhang, W.; Lin, J.; Liu, L.; Guha, C.; Kolb, E.A. Systemic Administration of Reovirus (REOLYSIN) inhibits Growth of Human Sarcoma Xenografts. Cancer 2011, 117, 1764–1774. [Google Scholar] [CrossRef]
- Zhao, X.; Chester, C.; Rajasekaran, N.; He, Z.; Kohrt, H.E. Strategic Combinations: The Future of Oncolytic Virotherapy with Reovirus. Mol. Cancer Ther. 2016, 15, 767–773. [Google Scholar] [CrossRef] [Green Version]
- Coffey, M.C.; Strong, J.E.; Forsyth, P.A.; Lee, P.W. Reovirus Therapy of Tumors with Activated RAS pathway. Science 1998, 282, 1332–1334. [Google Scholar] [CrossRef]
- Kelly, K.R.; Espitia, C.M.; Zhao, W.; Wu, K.; Visconte, V.; Anwer, F.; Calton, C.M.; Carew, J.S.; Nawrocki, S.T. Oncolytic Reovirus Sensitizes Multiple Myeloma Cells to Anti-Pd-l1 Therapy. Leukemia 2018, 32, 230–233. [Google Scholar] [CrossRef]
- Kelly, K.R.; Espitia, C.M.; Mahalingam, D.; Oyajobi, B.O.; Coffey, M.; Giles, F.J.; Carew, J.S.; Nawrocki, S.T. Reovirus Therapy stimulates Endoplasmic Reticular Stress, Noxa Induction, and Augments Bortezomib-Mediated Apoptosis in Multiple Myeloma. Oncogene 2012, 31, 3023–3038. [Google Scholar] [CrossRef] [Green Version]
- Vidal, L.; Pandha, H.S.; Yap, T.A.; White, C.L.; Twigger, K.; Vile, R.G.; Melcher, A.; Coffey, M.; Harrington, K.J.; DeBono, J.S. A Phase I Study of Intravenous Oncolytic Reovirus Type 3 Dearing in Patients with Advanced Cancer. Clin. Cancer Res. 2008, 14, 7127–7137. [Google Scholar] [CrossRef] [Green Version]
- Forsyth, P.; Roldan, G.; George, D.; Wallace, C.; Palmer, C.A.; Morris, D.; Cairncross, G.; Matthews, M.V.; Markert, J.; Gillespie, Y.; et al. A phase I Trial of Intratumoral Administration of Reovirus in Patients with Histologically Confirmed Recurrent malignant Gliomas. Mol. Ther. 2008, 16, 627–632. [Google Scholar] [CrossRef]
- Carew, J.S.; Espitia, C.M.; Zhao, W.; Mita, M.M.; Mita, A.C.; Nawrocki, S.T. Oncolytic Reovirus Inhibits Angiogenesis through Induction of cxcl10/ip-10 and Abrogation of Hif Activity in Soft Tissue Sarcomas. Oncotarget 2017, 8, 86769–86783. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Paek, A.L.; Hammer, M.; Rangarajan, S.; Ruijtenbeek, R.; Cooke, L.; Weterings, E.; Mahadevan, D. Drug-Induced Aneuploidy and Polyploidy Is a Mechanism of DISEASE relapse in myc/bcl2-Addicted Diffuse Large b-Cell Lymphoma. Oncotarget 2018, 9, 35875–35890. [Google Scholar] [CrossRef]
- Carew, J.S.; Espitia, C.M.; Zhao, W.; Han, Y.; Visconte, V.; Phillips, J.; Nawrocki, S.T. Disruption of Autophagic Degradation with Roc-325 Antagonizes Renal Cell Carcinoma Pathogenesis. Clin. Cancer Res. 2017, 23, 2869–2879. [Google Scholar] [CrossRef] [Green Version]
- Shigemura, T.; Shiohara, M.; Kato, M.; Furuta, S.; Kaneda, K.; Morishita, K.; Hasegawa, H.; Fujii, M.; Gorlach, A.; Koike, K.; et al. Superoxide-Generating Nox5alpha is Functionally Required for the Human t-Cell Leukemia Virus Type 1-Induced Cell Transformation Phenotype. J. Virol. 2015, 89, 9080–9089. [Google Scholar] [CrossRef] [Green Version]
- Barton, E.S.; Forrest, J.C.; Connolly, J.L.; Chappell, J.D.; Liu, Y.; Schnell, F.J.; Nusrat, A.; Parkos, C.A.; Dermody, T.S. Junction Adhesion Molecule is A Receptor for Reovirus. Cell 2001, 104, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Kelly, K.R.; Espitia, C.M.; Zhao, W.; Wendlandt, E.; Tricot, G.; Zhan, F.; Carew, J.S.; Nawrocki, S.T. Junctional Adhesion Molecule-A is Overexpressed in Advanced Multiple Myeloma and Determines Response to Oncolytic Reovirus. Oncotarget 2015, 6, 41275–41289. [Google Scholar] [CrossRef]
- Cao, S.; Strong, M.J.; Wang, X.; Moss, W.N.; Concha, M.; Lin, Z.; O’Grady, T.; Baddoo, M.; Fewell, C.; Renne, R.; et al. High-Throughput RNA Sequencing-Based Virome Analysis of 50 Lymphoma Cell Lines from the cancer Cell Line Encyclopedia Project. J. Virol. 2015, 89, 713–729. [Google Scholar] [CrossRef] [Green Version]
- Prodinger, W.M.; Larcher, C.; Schwendinger, M.; Dierich, M.P. Ligation of the FUNCTIONAL domain of Complement Receptor Type 2 (cr2, cd21) Is Relevant for Complex Formation in t Cell Lines. J. Immunol. 1996, 156, 2580–2584. [Google Scholar]
- Pinto, D.O.; DeMarino, C.; Pleet, M.L.; Cowen, M.; Branscome, H.; Al Sharif, S.; Jones, J.; Dutartre, H.; Lepene, B.; Liotta, L.A.; et al. Htlv-1 Extracellular Vesicles Promote Cell-to-Cell Contact. Front. Microbiol. 2019, 10, 2147. [Google Scholar] [CrossRef]
- Haller, S.D.; Monaco, M.L.; Essani, K. The Present Status of Immuno-Oncolytic Viruses in the Treatment of Pancreatic Cancer. Viruses 2020, 12, 1318. [Google Scholar] [CrossRef]
- Mahalingam, D.; Goel, S.; Aparo, S.; Patel Arora, S.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Gutierrez, A.; Coffey, M.; et al. A Phase II Study of Pelareorep (Reolysin((r))) in Combination with Gemcitabine for Patients with Advanced Pancreatic Adenocarcinoma. Cancers 2018, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, D.; Patel, S.; Nuovo, G.; Gill, G.; Selvaggi, G.; Coffey, M.; Nawrocki, S.T. The Combination of Intravenous Reolysin and Gemcitabine Induces Reovirus Replication and Endoplasmic Reticular Stress in a Patient with Kras-Activated Pancreatic Cancer. BMC Cancer 2015, 15, 513. [Google Scholar] [CrossRef] [Green Version]
- Kicielinski, K.P.; Chiocca, E.A.; Yu, J.S.; Gill, G.M.; Coffey, M.; Markert, J.M. Phase 1 Clinical Trial of Intratumoral Reovirus Infusion for the Treatment of Recurrent Malignant Gliomas in Adults. Mol. Ther. 2014, 22, 1056–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roulstone, V.; Khan, K.; Pandha, H.S.; Rudman, S.; Coffey, M.; Gill, G.M.; Melcher, A.A.; Vile, R.; Harrington, K.J.; de Bono, J.; et al. Phase I Trial of Cyclophosphamide as An Immune Modulator for Optimizing Oncolytic Reovirus Delivery to Solid Tumors. Clin. Cancer Res. 2015, 21, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.; Espitia, C.M.; Persky, D.O.; Carew, J.S.; Nawrocki, S.T. Resistance to Histone Deacetylase Inhibitors Confers Hypersensitivity to Oncolytic REOVIRUS therapy. Blood Adv. 2020, 4, 5297–5310. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.; Biron, V.L.; Fast, D.; Tam, R.; Carey, T.; Shmulevitz, M.; Seikaly, H. Oncolytic Activity of Reovirus in HPV Positive and Negative Head and Neck Squamous Cell Carcinoma. J. Otolaryngol. Head Neck Surg. 2015, 44, 8. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L.M.; Fredholm, S.; Joseph, C.; Nielsen, B.S.; Jonson, L.; Willerslev-Olsen, A.; Gluud, M.; Blumel, E.; Petersen, D.L.; Sibbesen, N.; et al. Stat5 Induces Mir-21 Expression in Cutaneous t Cell Lymphoma. Oncotarget 2016, 7, 45730–45744. [Google Scholar] [CrossRef] [Green Version]
- Yumeen, S.; Mirza, F.N.; Lewis, J.M.; King, A.L.O.; Kim, S.R.; Carlson, K.R.; Umlauf, S.R.; Surovtseva, Y.V.; Foss, F.M.; Girardi, M. Jak Inhibition Synergistically Potentiates Bcl2, BET, hdac, and Proteasome Inhibition in Advanced Ctcl. Blood Adv. 2020, 4, 2213–2226. [Google Scholar] [CrossRef]
- Kopp, K.L.; Ralfkiaer, U.; Gjerdrum, L.M.; Helvad, R.; Pedersen, I.H.; Litman, T.; Jonson, L.; Hagedorn, P.H.; Krejsgaard, T.; Gniadecki, R.; et al. Stat5-Mediated Expression of Oncogenic Mir-155 in Cutaneous t-Cell Lymphoma. Cell Cycle 2013, 12, 1939–1947. [Google Scholar] [CrossRef]
- Silbermann, K.; Schneider, G.; Grassmann, R. Stimulation of Interleukin-13 Expression by Human t-Cell LEUKEMIA virus Type 1 Oncoprotein Tax via a Dually Active Promoter Element Responsive to NF-kappab and Nfat. J. Gen. Virol. 2008, 89, 2788–2798. [Google Scholar] [CrossRef]
- McNamara, A.; Roebke, K.; Danthi, P. Cell Killing by Reovirus: Mechanisms and Consequences. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–20. [Google Scholar]
- Goody, R.J.; Beckham, J.D.; Rubtsova, K.; Tyler, K.L. Jak-stat Signaling Pathways Are Activated in the Brain Following Reovirus Infection. J. Neurovirol. 2007, 13, 373–383. [Google Scholar] [CrossRef]
- Felt, S.A.; Droby, G.N.; Grdzelishvili, V.Z. Ruxolitinib and Polycation Combination Treatment Overcomes Multiple Mechanisms of Resistance of Pancreatic Cancer Cells to ONCOLYTIC vesicular Stomatitis Virus. J. Virol. 2017, 91, 91. [Google Scholar] [CrossRef] [Green Version]
- Ghonime, M.G.; Cassady, K.A. Combination Therapy Using Ruxolitinib and Oncolytic HSV Renders Resistant Mpnsts Susceptible to Virotherapy. Cancer Immunol. Res. 2018, 6, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, C.; Iankov, I.D.; Anderson, S.K.; Aderca, I.; Leontovich, A.A.; Maurer, M.J.; Oberg, A.L.; Schroeder, M.A.; Giannini, C.; Greiner, S.M.; et al. Constitutive Interferon Pathway Activation in Tumors as An Efficacy Determinant Following Oncolytic Virotherapy. J. Natl. Cancer Inst. 2018, 110, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.R.; Dash, A.; Jacobson, B.A.; Ji, Y.; Baumann, D.; Ismail, K.; Kratzke, R.A. Jak/Stat Inhibition with Ruxolitinib Enhances Oncolytic Virotherapy in Non-Small Cell Lung Cancer Models. Cancer Gene Ther. 2019, 26, 411–418. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, S.; Espitia, C.M.; Persky, D.O.; Carew, J.S.; Nawrocki, S.T. Targeting JAK/STAT Signaling Antagonizes Resistance to Oncolytic Reovirus Therapy Driven by Prior Infection with HTLV-1 in Models of T-Cell Lymphoma. Viruses 2021, 13, 1406. https://doi.org/10.3390/v13071406
Islam S, Espitia CM, Persky DO, Carew JS, Nawrocki ST. Targeting JAK/STAT Signaling Antagonizes Resistance to Oncolytic Reovirus Therapy Driven by Prior Infection with HTLV-1 in Models of T-Cell Lymphoma. Viruses. 2021; 13(7):1406. https://doi.org/10.3390/v13071406
Chicago/Turabian StyleIslam, Shariful, Claudia M. Espitia, Daniel O. Persky, Jennifer S. Carew, and Steffan T. Nawrocki. 2021. "Targeting JAK/STAT Signaling Antagonizes Resistance to Oncolytic Reovirus Therapy Driven by Prior Infection with HTLV-1 in Models of T-Cell Lymphoma" Viruses 13, no. 7: 1406. https://doi.org/10.3390/v13071406
APA StyleIslam, S., Espitia, C. M., Persky, D. O., Carew, J. S., & Nawrocki, S. T. (2021). Targeting JAK/STAT Signaling Antagonizes Resistance to Oncolytic Reovirus Therapy Driven by Prior Infection with HTLV-1 in Models of T-Cell Lymphoma. Viruses, 13(7), 1406. https://doi.org/10.3390/v13071406