
The Molecular Evolution of Type 2 Vaccine-Derived Polioviruses in Individuals with Primary Immunodeficiency Diseases


Abstract
:1. Introduction
2. Definition and Classification of Vaccine-Derived Polioviruses
3. Status of Polio Outbreaks Due to cVDPV2
4. Prevalence of iVDPV2-Positive Cases
5. Genetic Analysis and Molecular Characterization of iVDPV2 Isolates
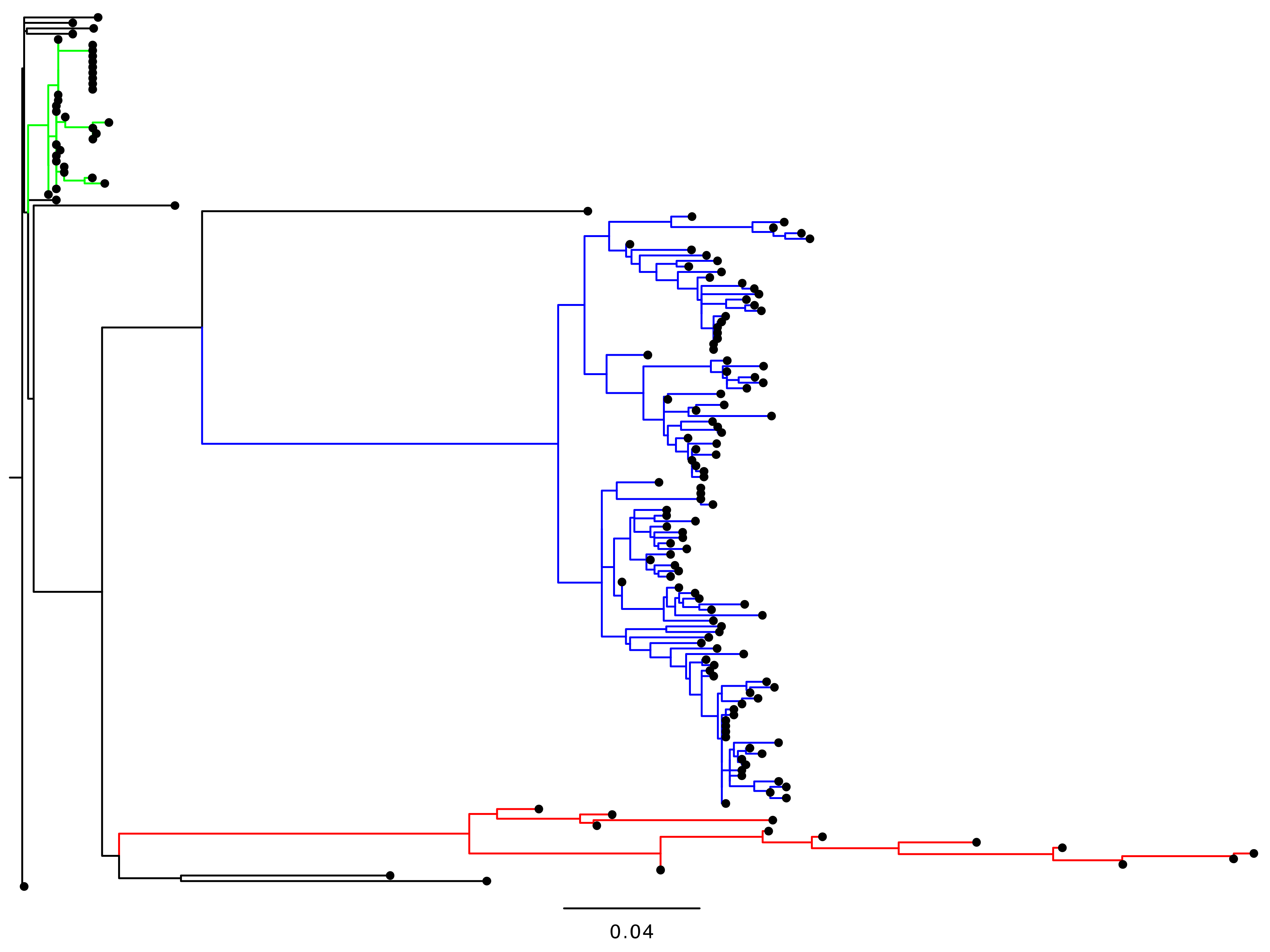
5.1. Available Sequence Dataset of iVDPV2 Isolates
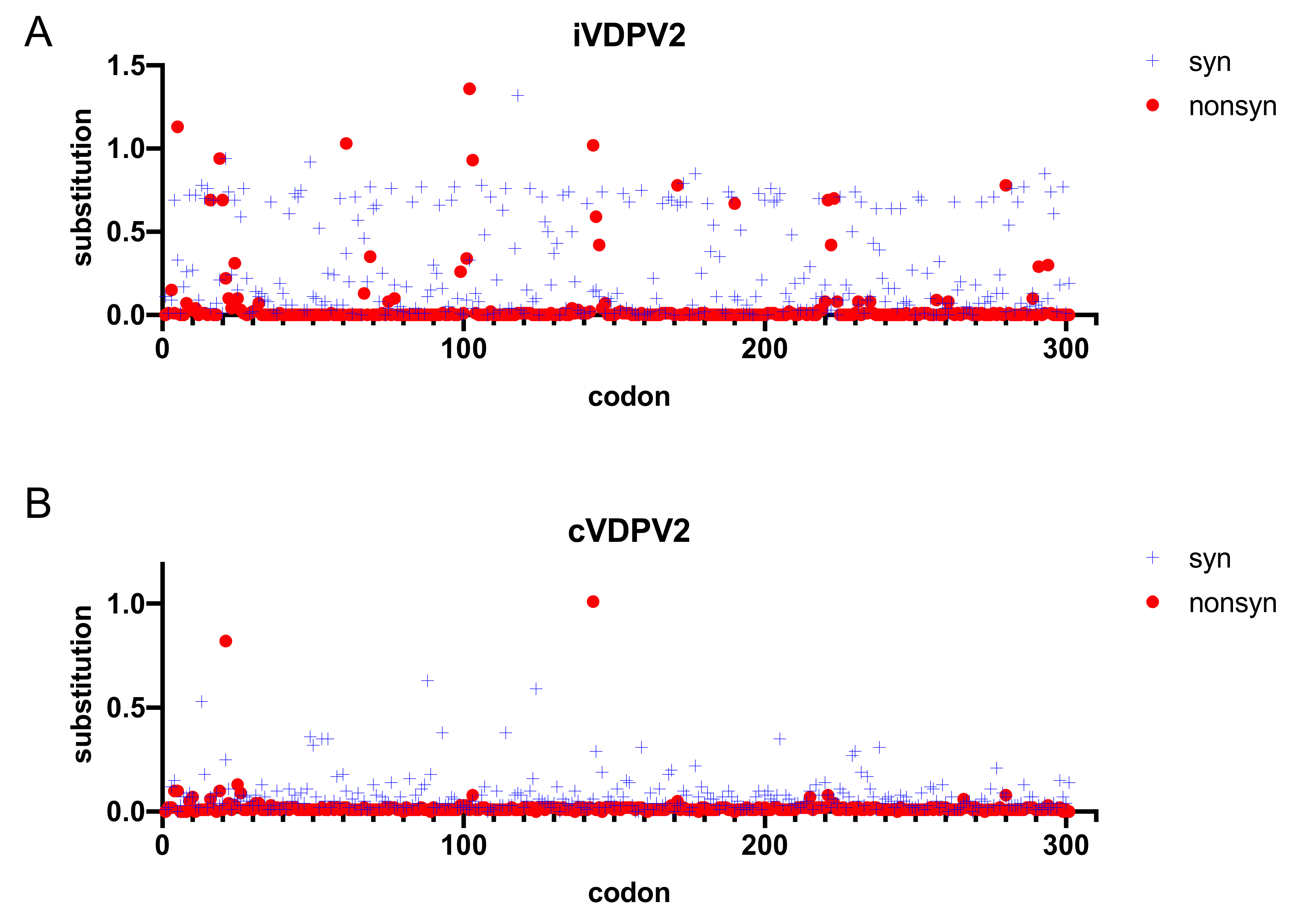
5.2. Molecular Evolution of iVDPV2
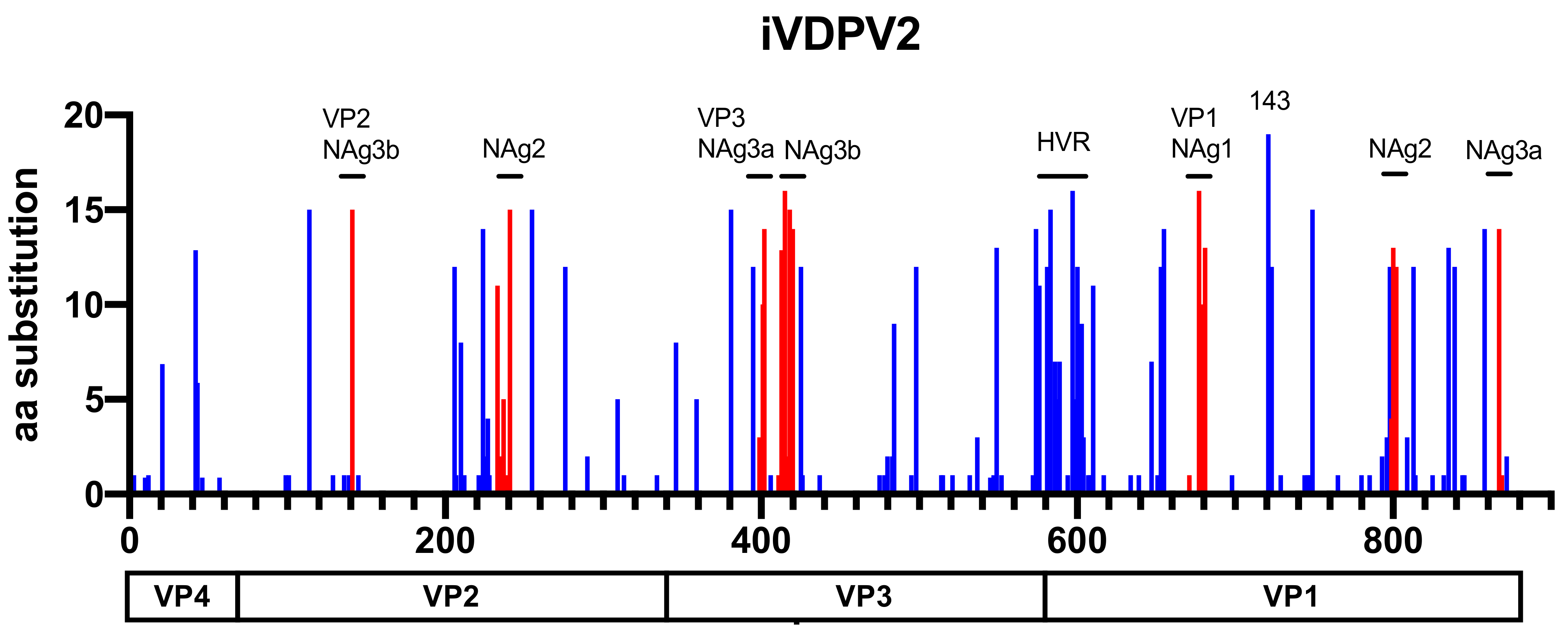
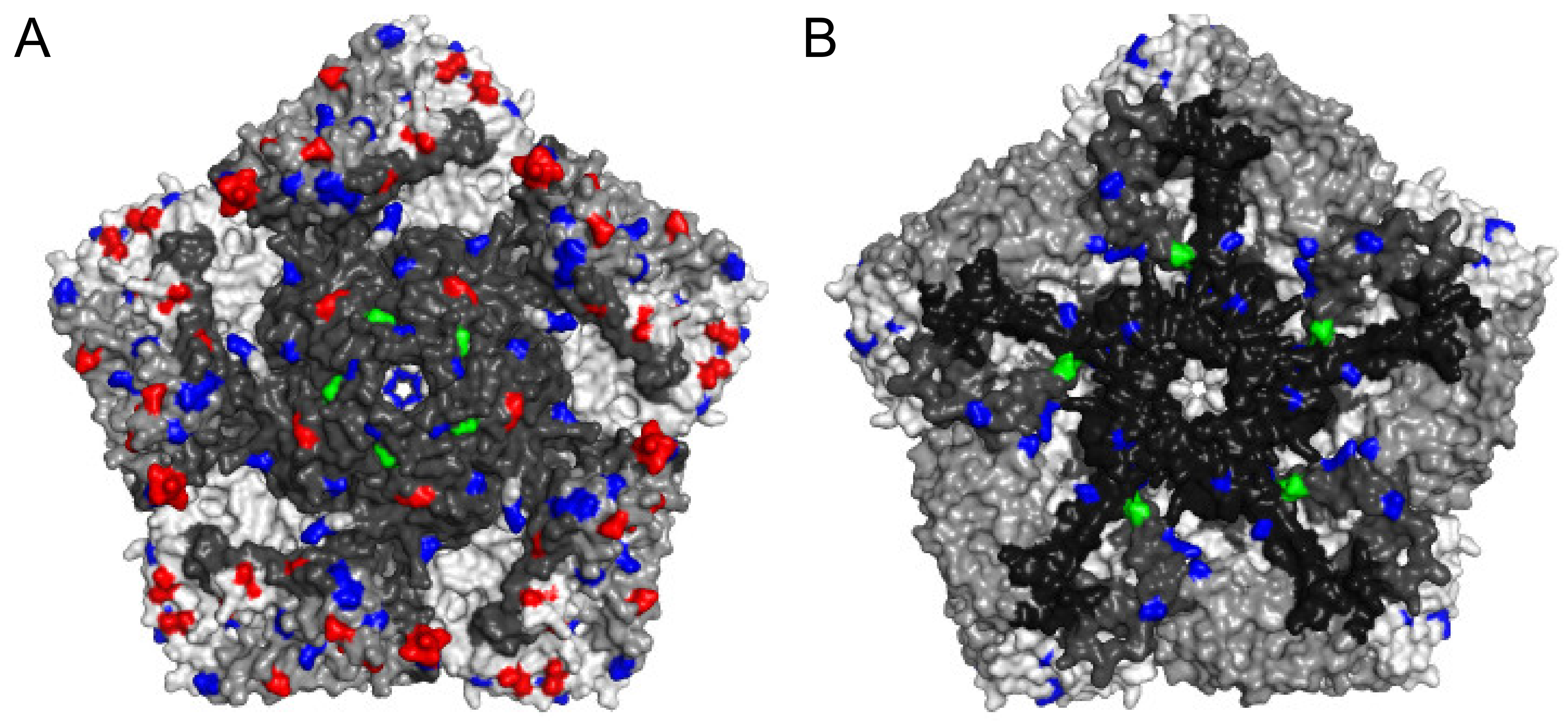
5.3. Amino Acid Substitutions in Phenotypic Determinants
5.4. Genomic Recombination
5.5. In Vivo Fitness and Quasi-Species
6. Current and Future Risk of iVDPV2
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, A.; Salisbury, D.M. Eradicating polio. Science 2015, 350, 609. [Google Scholar] [CrossRef] [Green Version]
- Makoni, M. Africa eradicates wild polio. Lancet Microbe 2020, 1, e243. [Google Scholar] [CrossRef]
- Tuma, J.N.; Wilkinson, A.L.; Diop, O.M.; Jorba, J.; Gardner, T.; Snider, C.J.; Anand, A.; Ahmed, J. Surveillance to track progress toward polio eradication—Worldwide, 2019–2020. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 667–673. [Google Scholar] [CrossRef]
- Global Polio Eradication Initiative. Wild Poliovirus List. Available online: https://polioeradication.org/wp-content/uploads/2021/06/weekly-polio-analyses-WPV-20210615.pdf (accessed on 2 July 2021).
- Cochi, S.L.; Pallansch, M.A. The long and winding road to eradicate vaccine-related polioviruses. J. Infect. Dis. 2021, 223, 7–9. [Google Scholar] [CrossRef]
- Chumakov, K.; Ehrenfeld, E.; Agol, V.I.; Wimmer, E. Polio eradication at the crossroads. Lancet Glob. Health 2021. [Google Scholar] [CrossRef]
- Global Polio Eradication Initiative. Circulating Vaccine-Derived Poliovirus. Available online: http://polioeradication.org/polio-today/polio-now/this-week/circulating-vaccine-derived-poliovirus/ (accessed on 2 July 2021).
- Alleman, M.M.; Jorba, J.; Greene, S.A.; Diop, O.M.; Iber, J.; Tallis, G.; Goel, A.; Wiesen, E.; Wassilak, S.G.F.; Burns, C.C. Update on vaccine-derived poliovirus outbreaks—Worldwide, July 2019–February 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Burns, C.C.; Diop, O.M.; Sutter, R.W.; Kew, O.M. Vaccine-derived polioviruses. J. Infect. Dis. 2014, 210 (Suppl. S1), S283–S293. [Google Scholar] [CrossRef]
- Yan, D.; Wang, D.; Zhang, Y.; Li, X.; Tang, H.; Guan, J.; Song, Y.; Zhu, S.; Xu, W. Implication of a High Risk for Type 2 Vaccine-Derived Poliovirus Emergence and Transmission After the Switch From Trivalent to Bivalent Oral Poliovirus Vaccine. J. Infect. Dis. 2021, 223, 113–118. [Google Scholar] [CrossRef]
- Meyer, E.; Sikka, N.; Durry, E.; Datta, D. Notes from the field: CDC polio surge response to expanding outbreaks of Type 2 circulating vaccine-derived poliovirus—Africa and Philippines, September 2019–March 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1182–1183. [Google Scholar] [CrossRef]
- Macklin, G.; Diop, O.M.; Humayun, A.; Shahmahmoodi, S.; El-Sayed, Z.A.; Triki, H.; Rey, G.; Avagyan, T.; Grabovac, V.; Jorba, J.; et al. Update on immunodeficiency-associated vaccine-derived polioviruses—Worldwide, July 2018–December 2019. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 913–917. [Google Scholar] [CrossRef]
- Aghamohammadi, A.; Abolhassani, H.; Kutukculer, N.; Wassilak, S.G.; Pallansch, M.A.; Kluglein, S.; Quinn, J.; Sutter, R.W.; Wang, X.; Sanal, O.; et al. Patients with primary immunodeficiencies are a reservoir of poliovirus and a risk to polio eradication. Front. Immunol. 2017, 8, 685. [Google Scholar] [CrossRef] [PubMed]
- Mbala-Kingebeni, P.; Pratt, C.; Mutafali-Ruffin, M.; Pauthner, M.G.; Bile, F.; Nkuba-Ndaye, A.; Black, A.; Kinganda-Lusamaki, E.; Faye, M.; Aziza, A.; et al. Ebola virus transmission initiated by relapse of systemic Ebola virus disease. N. Engl. J. Med. 2021, 384, 1240–1247. [Google Scholar] [CrossRef]
- Kemp, S.A.; Collier, D.A.; Datir, R.P.; Ferreira, I.; Gayed, S.; Jahun, A.; Hosmillo, M.; Rees-Spear, C.; Mlcochova, P.; Lumb, I.U.; et al. SARS-CoV-2 evolution during treatment of chronic infection. Nature 2021, 592, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Perelygina, L.; Chen, M.H.; Suppiah, S.; Adebayo, A.; Abernathy, E.; Dorsey, M.; Bercovitch, L.; Paris, K.; White, K.P.; Krol, A.; et al. Infectious vaccine-derived rubella viruses emerge, persist, and evolve in cutaneous granulomas of children with primary immunodeficiencies. PLoS Pathog. 2019, 15, e1008080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, K.S.; Stevens-Ayers, T.; Campbell, A.P.; Englund, J.A.; Pergam, S.A.; Boeckh, M.; Bloom, J.D. Parallel evolution of influenza across multiple spatiotemporal scales. Elife 2017, 6. [Google Scholar] [CrossRef]
- Burns, C.C.; Shaw, J.; Jorba, J.; Bukbuk, D.; Adu, F.; Gumede, N.; Pate, M.A.; Abanida, E.A.; Gasasira, A.; Iber, J.; et al. Multiple independent emergences of Type 2 vaccine-derived polioviruses during a large outbreak in northern Nigeria. J. Virol. 2013, 87, 4907–4922. [Google Scholar] [CrossRef] [Green Version]
- Kew, O.M.; Sutter, R.W.; de Gourville, E.M.; Dowdle, W.R.; Pallansch, M.A. Vaccine-derived polioviruses and the endgame strategy for global polio eradication. Annu. Rev. Microbiol. 2005, 59, 587–635. [Google Scholar] [CrossRef] [Green Version]
- Kew, O.M.; Wright, P.F.; Agol, V.I.; Delpeyroux, F.; Shimizu, H.; Nathanson, N.; Pallansch, M.A. Circulating vaccine-derived polioviruses: Current state of knowledge. Bull. World Health Organ. 2004, 82, 16–23. [Google Scholar]
- Global Polio Eradication Initiative. Polio Eradication and Endgame Strategic Plan 2013–2018. Available online: https://polioeradication.org/wp-content/uploads/2016/07/PEESP_EN_A4.pdf (accessed on 2 July 2021).
- Garon, J.; Seib, K.; Orenstein, W.A.; Ramirez Gonzalez, A.; Chang Blanc, D.; Zaffran, M.; Patel, M. Polio endgame: The global switch from tOPV to bOPV. Expert Rev. Vaccines 2016, 15, 693–708. [Google Scholar] [CrossRef]
- Global Polio Eradication Initiative. Introduction of inactivated poliovirus vaccine and switch from trivalent to bivalent oral poliovirus vaccine—Worldwide, 2013–2016. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 699–702. [Google Scholar]
- Global Polio Eradication Initiative. Global Circulating Vaccine-Derived Poliovirus (cVDPV) as of 01 June 2021. Available online: https://polioeradication.org/wp-content/uploads/2021/06/weekly-polio-analyses-cVDPV-20210601.pdf (accessed on 2 July 2021).
- Macklin, G.R.; O’Reilly, K.M.; Grassly, N.C.; Edmunds, W.J.; Mach, O.; Santhana Gopala Krishnan, R.; Voorman, A.; Vertefeuille, J.F.; Abdelwahab, J.; Gumede, N.; et al. Evolving epidemiology of poliovirus serotype 2 following withdrawal of the serotype 2 oral poliovirus vaccine. Science 2020, 368, 401–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez-Llorens, X.; Bandyopadhyay, A.S.; Gast, C.; Leon, T.; DeAntonio, R.; Jimeno, J.; Caballero, M.I.; Aguirre, G.; Oberste, M.S.; Weldon, W.C.; et al. Safety and immunogenicity of two novel type 2 oral poliovirus vaccine candidates compared with a monovalent type 2 oral poliovirus vaccine in children and infants: Two clinical trials. Lancet 2021, 397, 27–38. [Google Scholar] [CrossRef]
- Yeh, M.T.; Bujaki, E.; Dolan, P.T.; Smith, M.; Wahid, R.; Konz, J.; Weiner, A.J.; Bandyopadhyay, A.S.; Van Damme, P.; De Coster, I.; et al. Engineering the live-attenuated polio vaccine to prevent reversion to virulence. Cell Host Microbe 2020, 27, 736–751.e8. [Google Scholar] [CrossRef]
- Van Damme, P.; De Coster, I.; Bandyopadhyay, A.S.; Revets, H.; Withanage, K.; De Smedt, P.; Suykens, L.; Oberste, M.S.; Weldon, W.C.; Costa-Clemens, S.A.; et al. The safety and immunogenicity of two novel live attenuated monovalent (serotype 2) oral poliovirus vaccines in healthy adults: A double-blind, single-centre phase 1 study. Lancet 2019, 394, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Zomahoun, D.J.; Burman, A.L.; Snider, C.J.; Chauvin, C.; Gardner, T.; Lickness, J.S.; Ahmed, J.A.; Diop, O.; Gerber, S.; Anand, A. Impact of COVID-19 pandemic on global poliovirus surveillance. MMWR Morb. Mortal. Wkly. Rep. 2021, 69, 1648–1652. [Google Scholar] [CrossRef] [PubMed]
- WHO. Statement Following the Twenty-Eighth IHR Emergency Committee for Polio. Available online: https://www.who.int/news/item/21-05-2021-statement-following-the-twenty-eighth-ihr-emergency-committee-for-polio (accessed on 2 July 2021).
- Jorba, J.; Diop, O.M.; Iber, J.; Henderson, E.; Zhao, K.; Sutter, R.W.; Wassilak, S.G.F.; Burns, C.C. Update on vaccine-derived polioviruses—Worldwide, January 2017–June 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1189–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaghaghi, M.; Soleyman-Jahi, S.; Abolhassani, H.; Yazdani, R.; Azizi, G.; Rezaei, N.; Barbouche, M.R.; McKinlay, M.A.; Aghamohammadi, A. New insights into physiopathology of immunodeficiency-associated vaccine-derived poliovirus infection; systematic review of over 5 decades of data. Vaccine 2018, 36, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Update on vaccine-derived polioviruses. MMWR Morb. Mortal. Wkly. Rep. 2006, 55, 1093–1097. [Google Scholar]
- Jorgensen, D.; Pons-Salort, M.; Shaw, A.G.; Grassly, N.C. The role of genetic sequencing and analysis in the polio eradication programme. Virus Evol. 2020, 6, veaa040. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.; Yeh, M.T.; Zinger, T.; Smith, M.; Wright, C.; Ling, G.; Nielsen, R.; Macadam, A.; Andino, R. The evolutionary pathway to virulence of an RNA virus. Cell 2017, 169, 35–46.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, S.L.; Weldon, W.C.; Wei, L.; Chen, Q.; Shaw, J.; Zhao, K.; Jorba, J.; Kew, O.M.; Pallansch, M.A.; Burns, C.C.; et al. Neutralization capacity of highly divergent type 2 vaccine-derived polioviruses from immunodeficient patients. Vaccine 2020, 38, 3042–3049. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Jorba, J.; Shaw, J.; Iber, J.; Chen, Q.; Bullard, K.; Kew, O.M.; Burns, C.C. Are circulating type 2 vaccine-derived polioviruses (VDPVs) genetically distinguishable from immunodeficiency-associated VDPVs? Comput. Struct. Biotechnol. J. 2017, 15, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Schubert, A.; Böttcher, S.; Eis-Hübinger, A.M. Two cases of vaccine-derived poliovirus infection in an Oncology Ward. N. Engl. J. Med. 2016, 374, 1296–1298. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.S.; Harper, J.; Murray, A.; Lexau, C.; Bahta, L.; Christensen, J.; Cebelinski, E.; Fuller, S.; Kline, S.; Wallace, G.S.; et al. Vaccine-derived poliomyelitis 12 years after infection in Minnesota. N. Engl. J. Med. 2011, 364, 2316–2323. [Google Scholar] [CrossRef] [PubMed]
- Yakovenko, M.L.; Korotkova, E.A.; Ivanova, O.E.; Eremeeva, T.P.; Samoilovich, E.; Uhova, I.; Gavrilin, G.V.; Agol, V.I. Evolution of the Sabin vaccine into pathogenic derivatives without appreciable changes in antigenic properties: Need for improvement of current poliovirus surveillance. J. Virol. 2009, 83, 3402–3406. [Google Scholar] [CrossRef] [Green Version]
- Adu, F.; Iber, J.; Bukbuk, D.; Gumede, N.; Yang, S.J.; Jorba, J.; Campagnoli, R.; Sule, W.F.; Yang, C.F.; Burns, C.; et al. Isolation of recombinant type 2 vaccine-derived poliovirus (VDPV) from a Nigerian child. Virus Res. 2007, 127, 17–25. [Google Scholar] [CrossRef]
- Buttinelli, G.; Donati, V.; Fiore, S.; Marturano, J.; Plebani, A.; Balestri, P.; Soresina, A.R.; Vivarelli, R.; Delpeyroux, F.; Martin, J.; et al. Nucleotide variation in Sabin type 2 poliovirus from an immunodeficient patient with poliomyelitis. J. Gen. Virol. 2003, 84, 1215–1221. [Google Scholar] [CrossRef]
- Hovi, T.; Paananen, A.; Blomqvist, S.; Savolainen-Kopra, C.; Al-Hello, H.; Smura, T.; Shimizu, H.; Nadova, K.; Sobotova, Z.; Gavrilin, E.; et al. Characteristics of an environmentally monitored prolonged Type 2 vaccine derived poliovirus shedding episode that stopped without intervention. PLoS ONE 2013, 8, e66849. [Google Scholar] [CrossRef] [Green Version]
- Shulman, L.M.; Manor, Y.; Handsher, R.; Delpeyroux, F.; McDonough, M.J.; Halmut, T.; Silberstein, I.; Alfandari, J.; Quay, J.; Fisher, T.; et al. Molecular and antigenic characterization of a highly evolved derivative of the type 2 oral poliovaccine strain isolated from sewage in Israel. J. Clin. Microbiol. 2000, 38, 3729–3734. [Google Scholar] [CrossRef] [Green Version]
- Dunn, G.; Klapsa, D.; Wilton, T.; Stone, L.; Minor, P.D.; Martin, J. Twenty-eight years of poliovirus replication in an immunodeficient individual: Impact on the global polio eradication initiative. PLoS Pathog. 2015, 11, e1005114. [Google Scholar] [CrossRef]
- Weil, M.; Rahav, G.; Somech, R.; Stauber, T.; Alfandari, J.; Weiss, L.; Silberstein, I.; Indenbaum, V.; Or, I.B.; Mendelson, E.; et al. First report of a persistent oropharyngeal infection of type 2 vaccine-derived poliovirus (iVDPV2) in a primary immune deficient (PID) patient after eradication of wild type 2 poliovirus. Int. J. Infect. Dis. 2019, 83, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weil, M.; Shulman, L.M.; Heiman, S.; Stauber, T.; Alfandari, J.; Weiss, L.; Silberstein, I.; Indenbaum, V.; Mendelson, E.; Sofer, D. Prolonged excretion of type-2 poliovirus from a primary immune deficient patient during the transition to a type-2 poliovirus-free world, Israel, 2016. Euro Surveill. 2016, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Jorba, J.; Campagnoli, R.; De, L.; Kew, O. Calibration of multiple poliovirus molecular clocks covering an extended evolutionary range. J. Virol. 2008, 82, 4429–4440. [Google Scholar] [CrossRef] [Green Version]
- Shaghaghi, M.; Irannejad, M.; Abolhassani, H.; Shahmahmoodi, S.; Hamidieh, A.A.; Soleyman-Jahi, S.; Yazdani, R.; Azizi, G.; Aghamohammadi, A. Clearing vaccine-derived poliovirus infection following hematopoietic stem cell transplantation: A case report and review of literature. J. Clin. Immunol. 2018, 38, 610–616. [Google Scholar] [CrossRef]
- Odoom, J.K.; Yunus, Z.; Dunn, G.; Minor, P.D.; Martin, J. Changes in population dynamics during long-term evolution of Sabin type 1 poliovirus in an immunodeficient patient. J. Virol. 2008, 82, 9179–9190. [Google Scholar] [CrossRef] [Green Version]
- Valesano, A.L.; Taniuchi, M.; Fitzsimmons, W.J.; Islam, M.O.; Ahmed, T.; Zaman, K.; Haque, R.; Wong, W.; Famulare, M.; Lauring, A.S. The early evolution of oral poliovirus vaccine is shaped by strong positive selection and tight transmission bottlenecks. Cell Host Microbe 2021, 29, 32–43.e4. [Google Scholar] [CrossRef]
- Famulare, M.; Chang, S.; Iber, J.; Zhao, K.; Adeniji, J.A.; Bukbuk, D.; Baba, M.; Behrend, M.; Burns, C.C.; Oberste, M.S. Sabin vaccine reversion in the field: A comprehensive analysis of Sabin-like poliovirus isolates in Nigeria. J. Virol. 2016, 90, 317–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korber, B. HIV signature and sequence variation analysis. In Computational Analysis of HIV Molecular Sequences; Rodrigo, A.G., Learn, G.H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 55–72. [Google Scholar]
- Shaw, J.; Jorba, J.; Zhao, K.; Iber, J.; Chen, Q.; Adu, F.; Adeniji, A.; Bukbuk, D.; Baba, M.; Henderson, E.; et al. Dynamics of evolution of poliovirus neutralizing antigenic sites and other capsid functional domains during a large and prolonged outbreak. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Muslin, C.; Joffret, M.L.; Pelletier, I.; Blondel, B.; Delpeyroux, F. Evolution and emergence of enteroviruses through intra- and inter-species recombination: Plasticity and phenotypic impact of modular genetic exchanges in the 5′ untranslated region. PLoS Pathog. 2015, 11, e1005266. [Google Scholar] [CrossRef]
- Jegouic, S.; Joffret, M.L.; Blanchard, C.; Riquet, F.B.; Perret, C.; Pelletier, I.; Colbere-Garapin, F.; Rakoto-Andrianarivelo, M.; Delpeyroux, F. Recombination between polioviruses and co-circulating coxsackie A viruses: Role in the emergence of pathogenic vaccine-derived polioviruses. PLoS Pathog. 2009, 5, e1000412. [Google Scholar] [CrossRef]
- Rakoto-Andrianarivelo, M.; Guillot, S.; Iber, J.; Balanant, J.; Blondel, B.; Riquet, F.; Martin, J.; Kew, O.; Randriamanalina, B.; Razafinimpiasa, L.; et al. Co-circulation and evolution of polioviruses and species C enteroviruses in a district of Madagascar. PLoS Pathog. 2007, 3, e1911. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Zhu, S.L.; Yoshida, H.; Yoneyama, T.; Miyamura, T.; Shimizu, H. A Sabin 3-derived poliovirus recombinant contained a sequence homologous with indigenous human enterovirus species C in the viral polymerase coding region. J. Virol. 2005, 79, 12650–12657. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, H.; Thorley, B.; Paladin, F.J.; Brussen, K.A.; Stambos, V.; Yuen, L.; Utama, A.; Tano, Y.; Arita, M.; Yoshida, H.; et al. Circulation of type 1 vaccine-derived poliovirus in the Philippines in 2001. J. Virol. 2004, 78, 13512–13521. [Google Scholar] [CrossRef] [Green Version]
- Kew, O.; Morris-Glasgow, V.; Landaverde, M.; Burns, C.; Shaw, J.; Garib, Z.; Andre, J.; Blackman, E.; Freeman, C.J.; Jorba, J.; et al. Outbreak of poliomyelitis in Hispaniola associated with circulating type 1 vaccine-derived poliovirus. Science 2002, 296, 356–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuervo, N.S.; Guillot, S.; Romanenkova, N.; Combiescu, M.; Aubert-Combiescu, A.; Seghier, M.; Caro, V.; Crainic, R.; Delpeyroux, F. Genomic features of intertypic recombinant Sabin poliovirus strains excreted by primary vaccinees. J. Virol. 2001, 75, 5740–5751. [Google Scholar] [CrossRef] [Green Version]
- Gavrilin, G.V.; Cherkasova, E.A.; Lipskaya, G.Y.; Kew, O.M.; Agol, V.I. Evolution of circulating wild poliovirus and of vaccine-derived poliovirus in an immunodeficient patient: A unifying model. J. Virol. 2000, 74, 7381–7390. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.F.; Chen, H.Y.; Jorba, J.; Sun, H.C.; Yang, S.J.; Lee, H.C.; Huang, Y.C.; Lin, T.Y.; Chen, P.J.; Shimizu, H.; et al. Intratypic recombination among lineages of type 1 vaccine-derived poliovirus emerging during chronic infection of an immunodeficient patient. J. Virol. 2005, 79, 12623–12634. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, A.; Brodsky, L.; Andino, R. Mutational and fitness landscapes of an RNA virus revealed through population sequencing. Nature 2014, 505, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Vignuzzi, M.; Wendt, E.; Andino, R. Engineering attenuated virus vaccines by controlling replication fidelity. Nat. Med. 2008, 14, 154–161. [Google Scholar] [CrossRef]
- Vignuzzi, M.; Stone, J.K.; Arnold, J.J.; Cameron, C.E.; Andino, R. Quasispecies diversity determines pathogenesis through cooperative interactions in a viral population. Nature 2006, 439, 344–348. [Google Scholar] [CrossRef]
- Kalkowska, D.A.; Pallansch, M.A.; Cochi, S.L.; Kovacs, S.D.; Wassilak, S.G.F.; Thompson, K.M. Updated characterization of post-OPV cessation risks: Lessons from 2019 Serotype 2 outbreaks and implications for the probability of OPV restart. Risk Anal. 2021, 41, 320–328. [Google Scholar] [CrossRef]
- Duintjer Tebbens, R.J.; Thompson, K.M. Comprehensive screening for immunodeficiency-associated vaccine-derived poliovirus: An essential oral poliovirus vaccine cessation risk management strategy. Epidemiol. Infect. 2017, 145, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Gurung, S.; Harris, J.B.; Eltayeb, A.O.; Hampton, L.M.; Diorditsa, S.; Avagyan, T.; Schluter, W.W. Experience with inactivated polio vaccine introduction and the “switch” from trivalent to bivalent oral polio vaccine in the World Health Organization’s western Pacific region. J. Infect. Dis. 2017, 216 (Suppl. S1), S101–S108. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.; Boualam, L.; Diorditsa, S.; Gregory, C.; Jee, Y.; Mendoza-Aldana, J.; Roesel, S. Maintaining polio-free certification in the World Health Organization western Pacific Region for over a decade. J. Infect. Dis. 2014, 210 (Suppl. S1), S259–S267. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.M.; Zhang, Y.; Wang, X.Q.; Yu, W.Z.; Wen, N.; Yan, D.M.; Wang, H.Q.; Wushouer, F.; Wang, H.B.; Xu, A.Q.; et al. Identification and control of a poliomyelitis outbreak in Xinjiang, China. N. Engl. J. Med. 2013, 369, 1981–1990. [Google Scholar] [CrossRef]
- Bauri, M.; Wilkinson, A.L.; Ropa, B.; Feldon, K.; Snider, C.J.; Anand, A.; Tallis, G.; Boualam, L.; Grabovac, V.; Avagyan, T.; et al. Notes from the field: Circulating vaccine-derived poliovirus Type 1 and outbreak response—Papua New Guinea, 2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yan, D.; Zhu, S.; Nishimura, Y.; Ye, X.; Wang, D.; Jorba, J.; Zhu, H.; An, H.; Shimizu, H.; et al. An insight into recombination with Enterovirus species C and Nucleotide G-480 reversion from the viewpoint of neurovirulence of vaccine-derived polioviruses. Sci. Rep. 2015, 5, 17291. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case No. | Year Detected | Localization | PID Type | Serotype | Maximum VP1 Divergence (%) | Estimated Replication Period (years) | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 1981 | USA | CVID | 1 | 10 † | 7.6 † | [32] |
| 2 § | 1986 1992 | USA | CVID | 1 2 | 5.4 † 11.8 † | 4.7 ‡ 9.6 † | [32,33] |
| 3 | 1990 | Germany | CVID | 1 | 8.3 † | 9.5 † | [32] |
| 4 | 1995 | UK | CVID | 2 | 17.9 † | 27.83 † | [32] |
| 5 | 2000 | Germany | CVID | 1 | 8.5 † | 8.5 † | [32] |
| 6 | 2002 | UK | CVID | 2 | 6.3 † | 6.3 † | [32] |
| 7 | 2009 | India | CVID | 1 | 5.2 † | 5 † | [32] |
| 8 | 2009 | USA | CVID | 2 | 12.3 † | 11.9 † | [32] |
| 9 | 2015 | India | SCID | 3 | 10.2 † | 6 † | [32] |
| 10 | 2019 | Philippines | Hypokalemia and infectious diarrhea | 2 | 7.6 || | 5.0 || | [12] |
| GenBank Accession No. | Genome Region | Year Detected | Localization | Source of Samples | VP1 Divergence (%) | Estimated Replication Period (years) | Reference |
|---|---|---|---|---|---|---|---|
| GU390704 | P1 | 1992 | USA | NA | 10.4 | NA | [39] |
| KR817050-817060 AJ544513 | P1 Full genome | 1995-2013 | UK | Chronic excretor (Case 4 in Table 1) | 17.9 | 28 | [45] |
| AJ288062 | P1 | 1998 | Israel | Environmental samples | 9.4 | NA | [44] |
| AY177685 | Full genome | 2000 | Italy | iVDPV2 case | 0.88 | 1.42 | [42] |
| DQ890387 | Full genome | 2002 | Nigeria | iVDPV2 case | 2.5 | 1.5 | [41] |
| JX913541-913647 | VP1 | 2003-2005 | Slovakia | Environmental samples | 3.4 | NA | [43] |
| FJ517648 | P1 | 2007 | Belarus | iVDPV2 case (AFP) | 1.88 | 1.6 | [40] |
| FJ517649 | P1 | 2007 | Russia | iVDPV2 case (AFP) | 1.44 | NA (no OPV history) | [40] |
| GU390707 | Full genome | 2009 | USA | Chronic excretor (Case 8 in Table 1) | 11.83 | 11.9 | [39] |
| KR709241 | VP1 | 2013 | Germany | iVDPV2 case | 1.0 | 0.5–0.9 | [38] |
| KR709242 | VP1 | 2013 | Germany | iVDPV2 case | 4.4 | 2.4–2.8 | [38] |
| MK660464-660492 | VP1 | 2015 | Israel | iVDPV2 case | >0.8 | NA | [46,47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitamura, K.; Shimizu, H. The Molecular Evolution of Type 2 Vaccine-Derived Polioviruses in Individuals with Primary Immunodeficiency Diseases. Viruses 2021, 13, 1407. https://doi.org/10.3390/v13071407
Kitamura K, Shimizu H. The Molecular Evolution of Type 2 Vaccine-Derived Polioviruses in Individuals with Primary Immunodeficiency Diseases. Viruses. 2021; 13(7):1407. https://doi.org/10.3390/v13071407
Chicago/Turabian StyleKitamura, Kouichi, and Hiroyuki Shimizu. 2021. "The Molecular Evolution of Type 2 Vaccine-Derived Polioviruses in Individuals with Primary Immunodeficiency Diseases" Viruses 13, no. 7: 1407. https://doi.org/10.3390/v13071407
APA StyleKitamura, K., & Shimizu, H. (2021). The Molecular Evolution of Type 2 Vaccine-Derived Polioviruses in Individuals with Primary Immunodeficiency Diseases. Viruses, 13(7), 1407. https://doi.org/10.3390/v13071407