
Serological Evidence of Widespread Zika Transmission across the Philippines

 , , , and
, , , and
Abstract
:1. Introduction
2. Methods
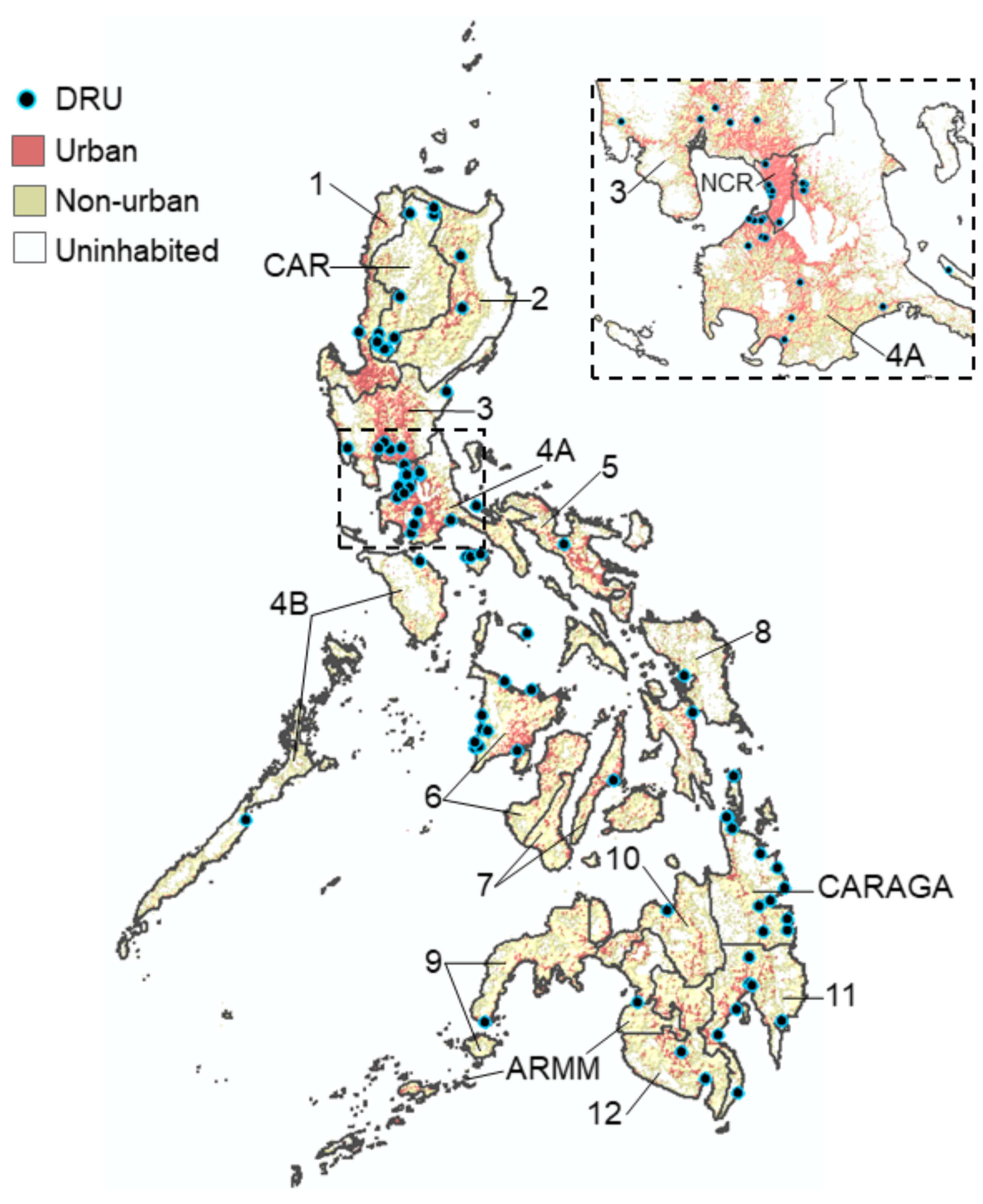
2.1. Flavivirus Surveillance in the Philippines
2.2. Data Collection
2.3. Laboratory Analysis
2.4. Data Analysis
3. Results
3.1. Data Description
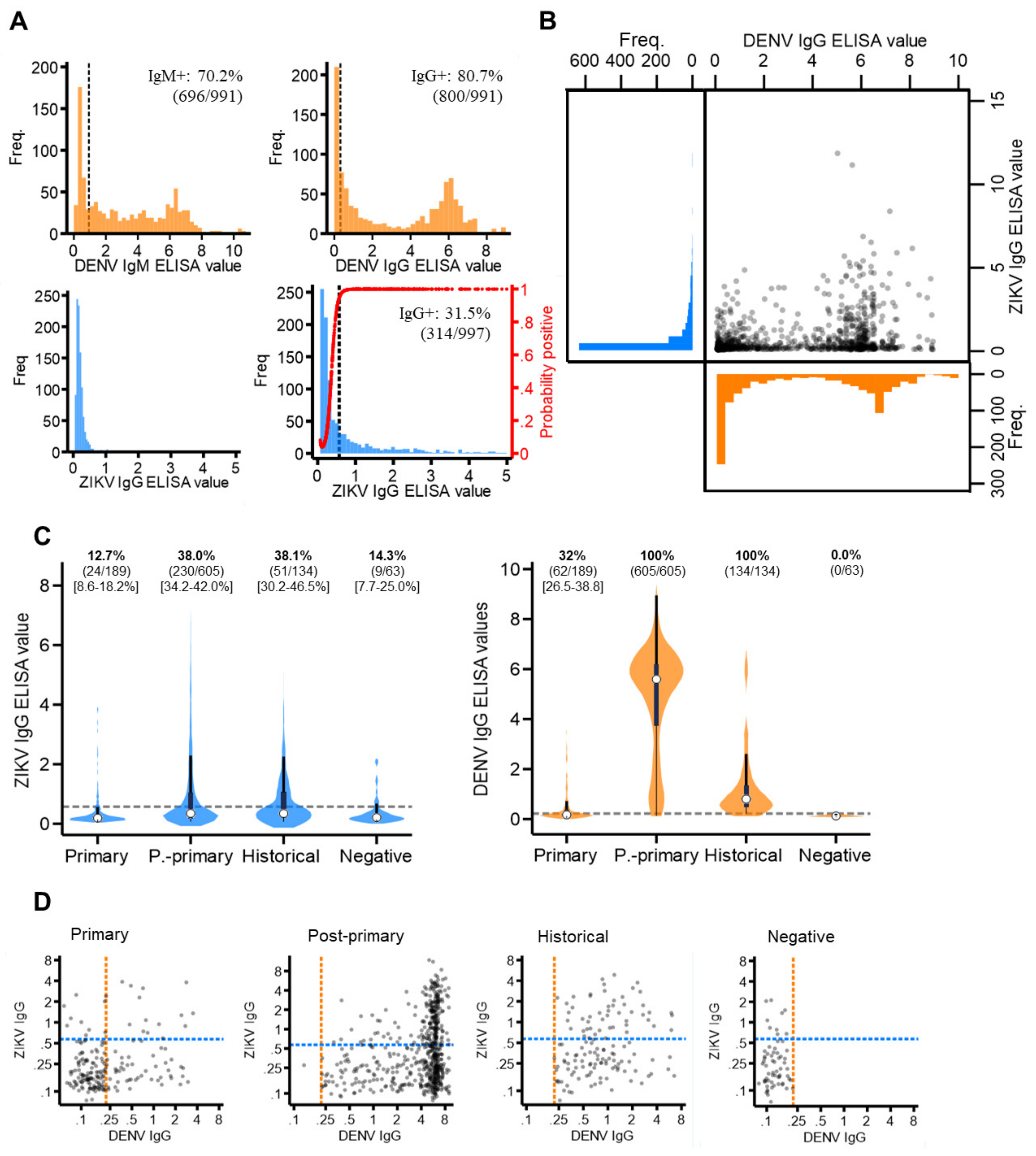
3.2. ZIKV and DENV Cross-Reactive Antibody Responses
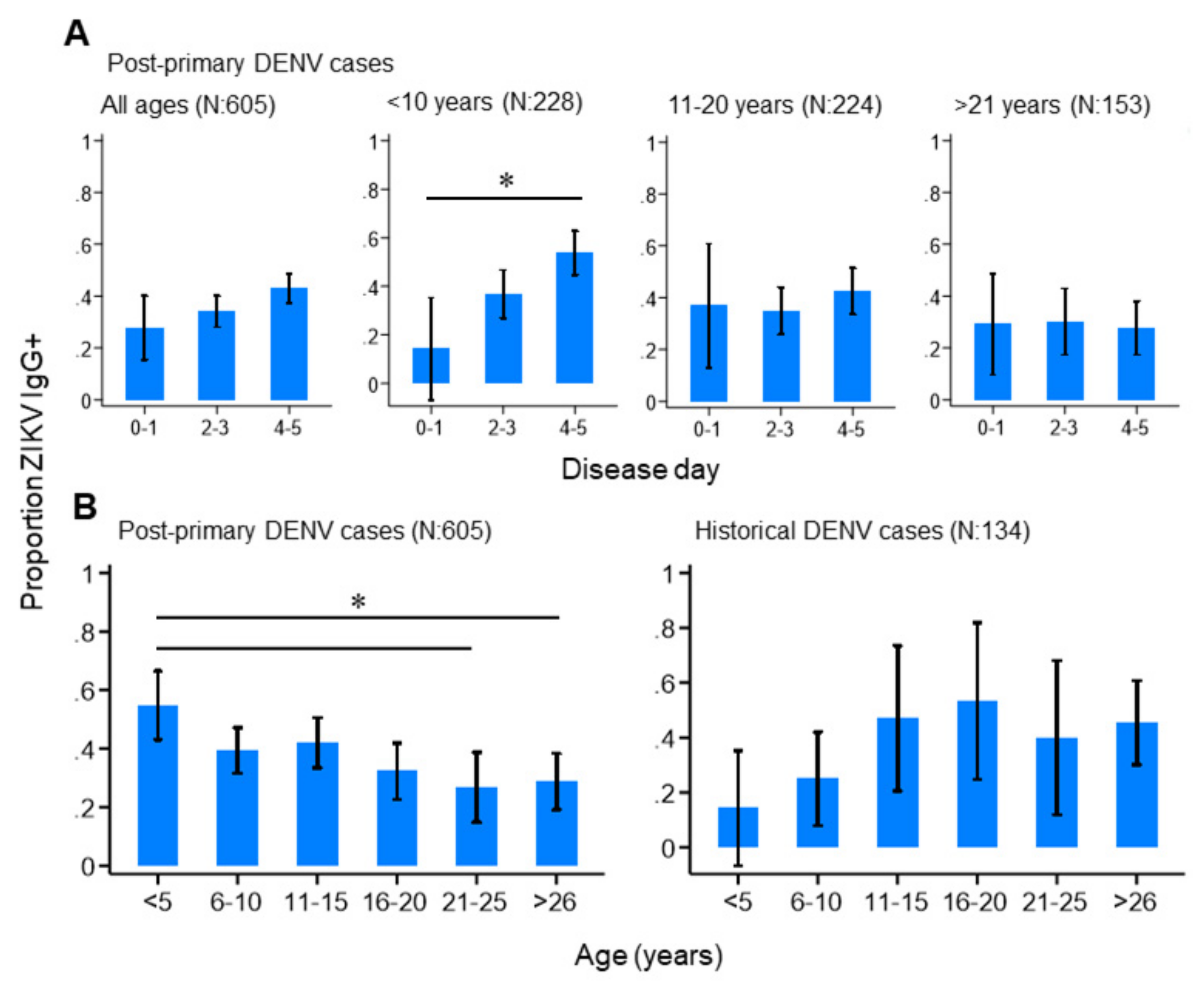
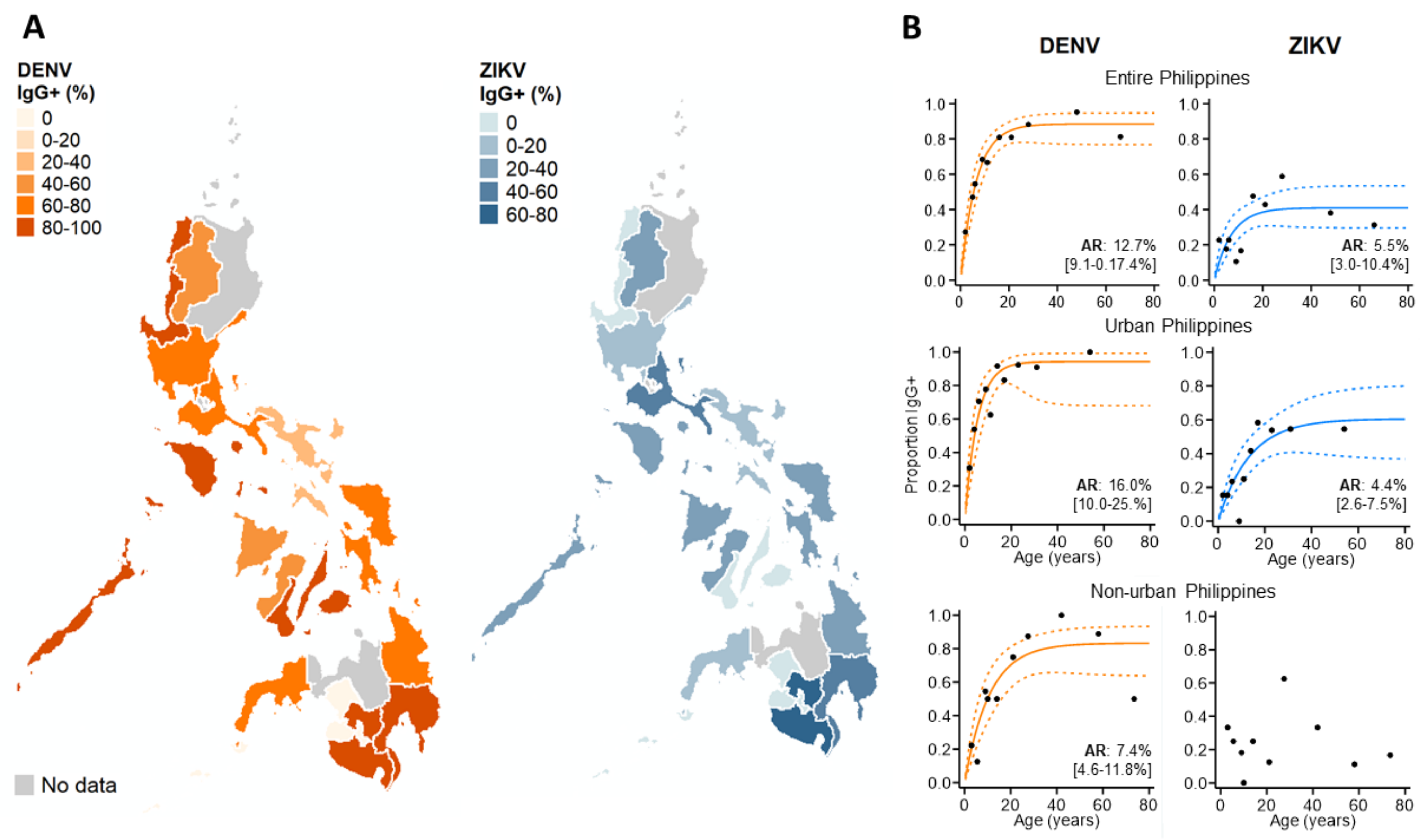
3.3. ZIKV Immunoepidemiology in the Philippines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Zika Epidemiological Update Report. Available online: https://www.who.int/emergencies/diseases/zika/zika-epidemiology-update-july-2019.pdf (accessed on 14 July 2021).
- Lowe, R.; Barcellos, C.; Brasil, P.; Cruz, O.G.; Honório, N.A.; Kuper, H.; Carvalho, M.S. The Zika Virus Epidemic in Brazil: From Discovery to Future Implications. Int. J. Environ. Res. Public Health 2018, 15, 96. [Google Scholar] [CrossRef] [Green Version]
- Cao-Lormeau, V.-M.; Blake, A.; Mons, S.; Lastère, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Osgood-Zimmerman, A.; Kassebaum, N.J.; Ray, S.E.; de Araújo, V.E.M.; Da Nóbrega, A.A.; Frutuoso, L.C.V.; Lecca, R.C.R.; Stevens, A.; Zoca de Oliveira, B.; et al. The association between Zika virus infection and microcephaly in Brazil 2015-2017: An observational analysis of over 4 million births. PLoS Med. 2019, 16, e1002755. [Google Scholar] [CrossRef]
- Lessler, J.; Chaisson, L.H.; Kucirka, L.M.; Bi, Q.; Grantz, K.; Salje, H.; Carcelen, A.C.; Ott, C.T.; Sheffield, J.S.; Ferguson, N.M.; et al. Assessing the global threat from Zika virus. Science 2016, 353, aaf8160. [Google Scholar] [CrossRef] [Green Version]
- Ruchusatsawat, K.; Wongjaroen, P.; Posanacharoen, A.; Rodriguez-Barraquer, I.; Sangkitporn, S.; Cummings, D.A.T.; Salje, H. Long-term circulation of Zika virus in Thailand: An observational study. Lancet Infect. Dis. 2019, 19, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Barraquer, I.; Costa, F.; Nascimento, E.J.M.; Nery, N.; Castanha, P.M.S.; Sacramento, G.A.; Cruz, J.; Carvalho, M.; De Olivera, D.; Hagan, J.E.; et al. Impact of preexisting dengue immunity on Zika virus emergence in a dengue endemic region. Science 2019, 363, 607–610. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.H.L.; Ho, H.J.; Chow, A.; Wong, J.; Kyaw, W.M.; Tan, A.; Chia, P.Y.; Choy, C.Y.; Tan, G.; Yeo, T.W.; et al. Correlation of clinical illness with viremia in Zika virus disease during an outbreak in Singapore. BMC Infect. Dis. 2018, 18, 1–7. [Google Scholar] [CrossRef]
- Okafor, I.I. Zika Virus: The Emerging Global Health Challenge. Divers. Equal. Health Care 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Stone, M.; Bakkour, S.; Lanteri, M.C.; Brambilla, D.; Simmons, G.; Bruhn, R.; Kaidarova, Z.; Lee, T.H.; Orlando Alsina, J.; Williamson, P.C.; et al. Zika virus RNA and IgM persistence in blood compartments and body fluids: A prospective observational study. Lancet Infect. Dis. 2020, 20, 1446–1456. [Google Scholar] [CrossRef]
- Pasquier, C.; Joguet, G.; Mengelle, C.; Chapuy-Regaud, S.; Pavili, L.; Prisant, N.; Izopet, J.; Bujan, L.; Mansuy, J.M. Kinetics of anti-ZIKV antibodies after Zika infection using two commercial enzyme-linked immunoassays. Diagn. Microbiol. Infect. Dis. 2018, 90, 26–30. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.S. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue Antibody-Dependent Enhancement: Knowns and Unknowns. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Martín-Acebes, M.A.; Saiz, J.-C.; Jiménez de Oya, N. Antibody-Dependent Enhancement and Zika: Real Threat or Phantom Menace? Front. Cell. Infect. Microbiol. 2018, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Culshaw, A.; Mongkolsapaya, J.; Screaton, G.R. The immunopathology of dengue and Zika virus infections. Curr. Opin. Immunol. 2017, 48, 1–6. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Narvaez, C.; Arguello, S.; Lopez Mercado, B.; Collado, D.; Ampie, O.; Elizondo, D.; Miranda, T.; Bustos Carillo, F.; Mercado, J.C.; et al. Zika virus infection enhances future risk of severe dengue disease. Science 2020, 369, 1123–1128. [Google Scholar] [CrossRef]
- George, J.; Valiant, W.G.; Mattapallil, M.J.; Walker, M.; Huang, Y.-J.S.; Vanlandingham, D.L.; Misamore, J.; Greenhouse, J.; Weiss, D.E.; Verthelyi, D.; et al. Prior Exposure to Zika Virus Significantly Enhances Peak Dengue-2 Viremia in Rhesus Macaques. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Ariën, K.K.; Michiels, J.; Foqué, N.; Heyndrickx, L.; Van Esbroeck, M. Can Zika virus antibodies cross-protect against dengue virus? Lancet Glob. Health 2018, 6, e494. [Google Scholar] [CrossRef] [Green Version]
- Valiant, W.G.; Huang, Y.-J.S.; Vanlandingham, D.L.; Higgs, S.; Lewis, M.G.; Mattapallil, J.J. Zika convalescent macaques display delayed induction of anamnestic cross-neutralizing antibody responses after dengue infection. Emerg. Microbes Infect. 2018, 7, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elisa, N.S. Zika Virus Infections EUROIMMUN Test Systems for the Diagnosis of Zika Virus Infections. Available online: https//:www.euroimmun.com/documents/Indications/Infections/Zika-virus/HI_2668_I_UK_B.pdf (accessed on 14 July 2021).
- Huzly, D.; Hanselmann, I.; Schmidt-Chanasit, J.; Panning, M. High specificity of a novel Zika virus ELISA in European patients after exposure to different flaviviruses. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Steinhagen, K.; Probst, C.; Radzimski, C.; Schmidt-Chanasit, J.; Emmerich, P.; Van Esbroeck, M.; Schinkel, J.; Grobusch, M.P.; Goorhuis, A.; Warnecke, J.M.; et al. Serodiagnosis of Zika virus (ZIKV) infections by a novel NS1-based ELISA devoid of cross-reactivity with dengue virus antibodies: A multicohort study of assay performance, 2015 to 2016. Eurosurveillance 2016, 21, 30426. [Google Scholar] [CrossRef] [Green Version]
- Medialdea-Carrera, R.; Levy, F.; Castanha, P.; de Sequeira, P.C.; Brasil, P.; Lewis-Ximenez, L.L.; Turtle, L.; Solomon, T.; Bispo de Filippis, A.M.; Brown, D.W.; et al. A systematic evaluation of IgM and IgG antibody assay accuracy in diagnosing acute Zika Virus infection in Brazil; lessons relevant to emerging infections. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kikuti, M.; Tauro, L.B.; Moreira, P.S.S.; Campos, G.S.; Paploski, I.A.D.; Weaver, S.C.; Reis, M.G.; Kitron, U.; Ribeiro, G.S. Diagnostic performance of commercial IgM and IgG enzyme-linked immunoassays (ELISAs) for diagnosis of Zika virus infection. Virol. J. 2018, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- Morales, I.; Rosenberger, K.D.; Magalhaes, T.; Morais, C.N.L.; Braga, C.; Marques, E.T.A.; Calvet, G.A.; Damasceno, L.; Brasil, P.; Bispo de Filippis, A.M.; et al. Diagnostic performance of anti-Zika virus IgM, IgAM and IgG ELISAs during co-circulation of Zika, dengue, and chikungunya viruses in Brazil and Venezuela. PLoS Negl. Trop. Dis. 2021, 15, e0009336. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.; Wang, X.; Donolato, M.; Harris, E.; Cruz, M.M.; Balmaseda, A.; Wang, R.Y.L. Evaluation of ViroTrack Sero Zika IgG/IgM, a New Rapid and Quantitative Zika Serological Diagnostic Assay. Diagnostics 2020, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Buerano, C.C.; Pangilinan, L.A.S.; Dimamay, M.T.A.; Mapua, C.A.; Dimamay, M.P.S.; Matias, R.R.; Natividad, F.F.; Daroy, M.L.G.; Hasebe, F.; Morita, K.; et al. Zika virus infection, philippines, 2012. Emerg. Infect. Dis. 2020, 26, 2300–2301. [Google Scholar] [CrossRef] [PubMed]
- Alera, M.T.; Hermann, L.; Tac-An, I.A.; Klungthong, C.; Rutvisuttinunt, W.; Manasatienkij, W.; Villa, D.; Thaisomboonsuk, B.; Velasco, J.M.; Chinnawirotpisan, P.; et al. Zika virus infection, philippines, 2012. Emerg. Infect. Dis. 2015, 21, 722–724. [Google Scholar] [CrossRef]
- Lonogan, K.; de Guzman, A.; Delos Reyes, V.C.; Sucaldito, M.N.; Avelino, F. The enhanced Zika surveillance in the Philippines, November 14, 2016–February 28, 2017. Int. J. Infect. Dis. 2020, 101, 232–233. [Google Scholar] [CrossRef]
- Department of Health (DoH). Philippine Integrated Disease Surveillance and Response; National Epidemiology Centre: Madrid, Spain, 2014. [Google Scholar]
- Johnson, B.W.; Russell, B.J.; Lanciotti, R.S. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J. Clin. Microbiol. 2005, 43, 4977–4983. [Google Scholar] [CrossRef] [Green Version]
- Biggs, J.R.; Sy, A.K.; Brady, O.J.; Kucharski, A.J.; Funk, S.; Reyes, M.A.J.; Quinones, M.A.; Jones-Warner, W.; Tu, Y.-H.; Avelino, F.L.; et al. A serological framework to investigate acute primary and post-primary dengue cases reporting across the Philippines. BMC Med. 2020, 18, 364. [Google Scholar] [CrossRef]
- Sepúlveda, N.; Stresman, G.; White, M.T.; Drakeley, C.J. Current Mathematical Models for Analyzing Anti-Malarial Antibody Data with an Eye to Malaria Elimination and Eradication. J. Immunol. Res. 2015, 2015, 738030. [Google Scholar] [CrossRef]
- Kucharski, A.J.; Kama, M.; Watson, C.H.; Aubry, M.; Funk, S.; Henderson, A.D.; Brady, O.J.; Vanhomwegen, J.; Manuguerra, J.-C.; Lau, C.L.; et al. Using paired serology and surveillance data to quantify dengue transmission and control during a large outbreak in Fiji. Elife 2018, 7, e34848. [Google Scholar] [CrossRef]
- Henderson, A.D.; Aubry, M.; Kama, M.; Vanhomwegen, J.; Teissier, A.; Mariteragi-Helle, T.; Paoaafaite, T.; Teissier, Y.; Manuguerra, J.-C.; Edmunds, J.; et al. Zika seroprevalence declines and neutralizing antibodies wane in adults following outbreaks in French Polynesia and Fiji. Elife 2020, 9, e48460. [Google Scholar] [CrossRef]
- Rodriguez-Barraquer, I.; Salje, H.; Cummings, D.A. Opportunities for improved surveillance and control of dengue from age-specific case data. Elife 2019, 8, e45474. [Google Scholar] [CrossRef]
- Wikramaratna, P.S.; Simmons, C.P.; Gupta, S.; Recker, M. The Effects of Tertiary and Quaternary Infections on the Epidemiology of Dengue. PLoS ONE 2010, 5, e12347. [Google Scholar] [CrossRef] [Green Version]
- Salje, H.; Paul, K.K.; Paul, R. Nationally-representative serostudy of dengue in Bangladesh allows generalizable disease burden estimates. Elife 2019, 8, e42869. [Google Scholar] [CrossRef]
- Henderson, A.D.; Kama, M.; Aubry, M.; Hue, S.; Teissier, A.; Naivalu, T.; Bechu, V.D.; Kailawadoko, J.; Rabukawaqa, I.; Sahukhan, A.; et al. Interactions between timing and transmissibility explain diverse flavivirus dynamics in Fiji. Nat. Commun. 2021, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ho, Z.J.M.; Hapuarachchi, H.C.; Barkham, T.; Chow, A.; Ng, L.C.; Lee, J.M.V.; Leo, Y.S.; Prem, K.; Lim, Y.H.G.; de Sessions, P.F.; et al. Outbreak of Zika virus infection in Singapore: An epidemiological, entomological, virological, and clinical analysis. Lancet Infect. Dis. 2017, 17, 813–821. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reported DENV/ZIKV | N | Adverse Clinical Symptoms | Severe Symptoms | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Immune Status | % | OR | [95% CI] | p-Value | % | OR | [95% CI] | p-Value | |
| Primary DENV | 131 | 73.3 | 1 (ref) | 9.2 | 1 (ref) | ||||
| (ZIKV IgG-) | |||||||||
| Post-primary DENV | 306 | 80.7 | 1.53 | [0.94–2.47] | 0.084 | 6.9 | 0.69 | [0.33–1.34] | 0.254 |
| (ZIKV IgG-) | |||||||||
| Post-primary DENV | 190 | 87.4 | 2.52 | [1.42–4.49] | 0.002 | 12.6 | 1.31 | [0.66–2.60] | 0.438 |
| (ZIKV IgG+) | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biggs, J.R.; Sy, A.K.; Brady, O.J.; Kucharski, A.J.; Funk, S.; Tu, Y.-H.; Reyes, M.A.J.; Quinones, M.A.; Jones-Warner, W.; Ashall, J.; et al. Serological Evidence of Widespread Zika Transmission across the Philippines. Viruses 2021, 13, 1441. https://doi.org/10.3390/v13081441
Biggs JR, Sy AK, Brady OJ, Kucharski AJ, Funk S, Tu Y-H, Reyes MAJ, Quinones MA, Jones-Warner W, Ashall J, et al. Serological Evidence of Widespread Zika Transmission across the Philippines. Viruses. 2021; 13(8):1441. https://doi.org/10.3390/v13081441
Chicago/Turabian StyleBiggs, Joseph R., Ava Kristy Sy, Oliver J. Brady, Adam J. Kucharski, Sebastian Funk, Yun-Hung Tu, Mary Anne Joy Reyes, Mary Ann Quinones, William Jones-Warner, James Ashall, and et al. 2021. "Serological Evidence of Widespread Zika Transmission across the Philippines" Viruses 13, no. 8: 1441. https://doi.org/10.3390/v13081441
APA StyleBiggs, J. R., Sy, A. K., Brady, O. J., Kucharski, A. J., Funk, S., Tu, Y. -H., Reyes, M. A. J., Quinones, M. A., Jones-Warner, W., Ashall, J., Avelino, F. L., Sucaldito, N. L., Tandoc, A. O., Cutiongco-de la Paz, E., Capeding, M. R. Z., Padilla, C. D., Hibberd, M. L., & Hafalla, J. C. R. (2021). Serological Evidence of Widespread Zika Transmission across the Philippines. Viruses, 13(8), 1441. https://doi.org/10.3390/v13081441