
Yellow Fever Outbreak in Eastern Senegal, 2020–2021

 , , , ,
, , , ,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Surveillance
2.2. Serology Assays
2.3. Molecular Diagnostics
2.4. Mosquito Collections and Processing
2.5. Virus Isolation and Identification from Mosquito Pools
2.6. Next Generation Sequencing of Yellow Fever Virus
2.7. Phylogenetic Analysis
3. Results
3.1. First Detected Yellow Fever Human Cases
3.2. Yellow Fever Virus Circulation in the Human Population in Kedougou
3.3. Yellow Fever Virus Circulation in Mosquitoes from Kedougou
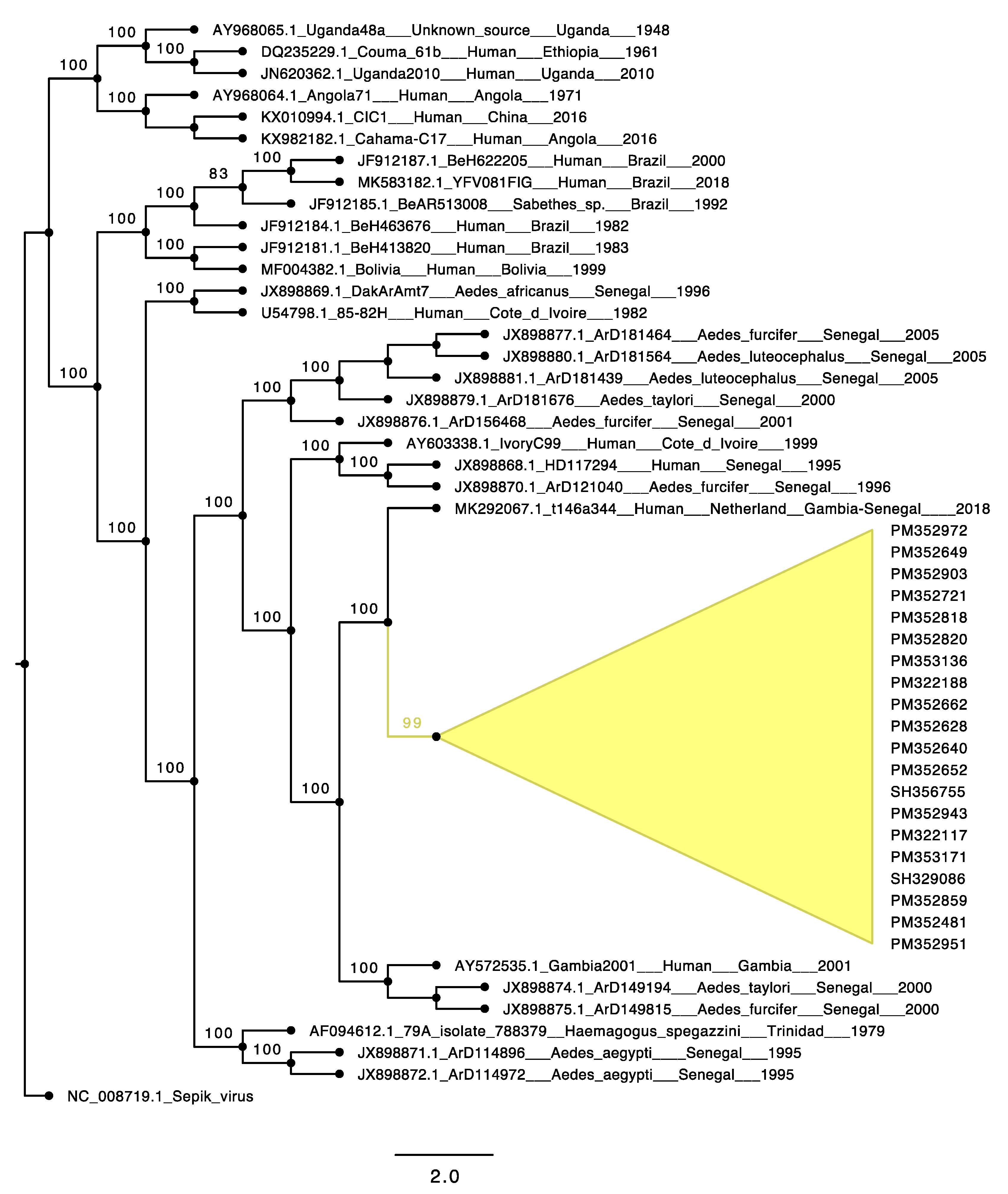
3.4. Phylogenetic Analyses of the Yellow Fever Virus Strain Causing the Outbreak in Eastern Senegal
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Staples, J.E.; Monath, T.P. Yellow fever: 100 years of discovery. JAMA 2008, 300, 960–962. [Google Scholar] [CrossRef]
- Chambers, T.J.; McCourt, D.W.; Rice, C.M. Production of yellow fever virus proteins in infected cells: Identification of discrete polyprotein species and analysis of cleavage kinetics using region-specific polyclonal antisera. Virology 1990, 177, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide sequence of yellow fever virus: Implications for flavivirus gene expression and evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Lorente, S.; Romero-López, C.; Berzal-Herranz, A. Information Encoded by the Flavivirus Genomes beyond the Nucleotide Sequence. Int. J. Mol. Sci. 2021, 22, 3738. [Google Scholar] [CrossRef] [PubMed]
- Douam, F.; Ploss, A. Yellow fever Virus: Knowledge Gaps Impeding the Fight Against an Old Foe. Trends Microbiol. 2018, 26, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.A.; Vasconcelos, P.F.; Staples, J.E. The whole iceberg: Estimating the incidence of yellow fever virus infection from the number of severe cases. Trans. R Soc. Trop. Med. Hyg. 2014, 108, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P.; Vasconcelos, P.F. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- Diallo, D.; Fall, G.; Diagne, C.T.; Gaye, A.; Ba, Y.; Dia, I.; Faye, O.; Diallo, M. Concurrent amplification of Zika, chikungunya, and yellow fever virus in a sylvatic focus of arboviruses in Southeastern Senegal, 2015. BMC Microbiol. 2020, 20, 181. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Wilson, M.E. Yellow fever control: Current epidemiology and vaccination strategies. Trop. Dis. Travel Med. Vaccines 2020, 6, 1. [Google Scholar] [CrossRef]
- Sall, A.A.; Faye, O.; Diallo, M.; Firth, C.; Kitchen, A.; Holmes, E.C. Yellow fever virus exhibits slower evolutionary dynamics than dengue virus. J. Virol. 2010, 84, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Traoré-Lamizana, M.; Fontenille, D.; Zeller, H.G.; Mondo, M.; Diallo, M.; Adam, F.; Eyraud, M.; Maiga, A.; Digoutte, J.P. Surveillance for yellow fever virus in eastern Senegal during 1993. J. Med. Entomol. 1996, 33, 760–765. [Google Scholar] [CrossRef] [Green Version]
- Thonnon, J.; Fontenille, D.; Tall, A.; Diallo, M.; Renaudineau, Y.; Baudez, B.; Raphenon, G. Re-emergence of yellow fever in Senegal in 1995. Am. J. Trop. Med. Hyg. 1998, 59, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Althouse, B.M.; Hanley, K.A.; Diallo, M.; Sall, A.A.; Ba, Y.; Faye, O.; Diallo, D.; Watts, D.M.; Weaver, S.C.; Cummings, D.A. Impact of climate and mosquito vector abundance on sylvatic arbovirus circulation dynamics in Senegal. Am. J. Trop. Med. Hyg. 2015, 92, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Sow, A.; Loucoubar, C.; Diallo, D.; Faye, O.; Ndiaye, Y.; Senghor, C.S.; Dia, A.T.; Faye, O.; Weaver, S.C.; Diallo, M.; et al. Concurrent malaria and arbovirus infections in Kedougou, southeastern Senegal. Malar. J. 2016, 15, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Recommended Surveillance Standards, 2nd ed.; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Barry, M.A.; Arinal, F.; Talla, C.; Hedible, B.G.; Sarr, F.D.; Ba, I.O.; Diop, B.; Dia, N.; Vray, M. Performance of case definitions and clinical predictors for influenza surveillance among patients followed in a rural cohort in Senegal. BMC Infect. Dis. 2021, 21, 31. [Google Scholar] [CrossRef]
- Sow, A.; Faye, O.; Diallo, M.; Diallo, D.; Chen, R.; Faye, O.; Diagne, C.T.; Guerbois, M.; Weidmann, M.; Ndiaye, Y.; et al. Chikungunya Outbreak in Kedougou, Southeastern Senegal in 2009-2010. Open Forum. Infect. Dis. 2017, 5, ofx259. [Google Scholar] [CrossRef]
- Diallo, M.; Ba, Y.; Sall, A.A.; Diop, O.M.; Ndione, J.A.; Mondo, M.; Girault, L.; Mathiot, C. Amplification of the sylvatic cycle of dengue virus type 2, Senegal, 1999-2000: Entomologic findings and epidemiologic considerations. Emerg. Infect. Dis. 2003, 362–367. [Google Scholar] [CrossRef]
- Weidmann, M.; Faye, O.; Faye, O.; Kranaster, R.; Marx, A.; Nunes, M.R.; Vasconcelos, P.F.; Hufert, F.T.; Sall, A.A. Improved LNA probe-based assay for the detection of African and South American yellow fever virus strains. J. Clin. Virol. 2010, 48, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Diallo, D.; Sall, A.A.; Diagne, C.T.; Faye, O.; Hanley, K.A.; Buenemann, M.; Ba, Y.; Faye, O.; Weaver, S.C.; Diallo, M. Patterns of a sylvatic yellow fever virus amplification in southeastern Senegal, 2010. Am. J. Trop. Med. Hyg. 2014, 90, 1003–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, D.; Sall, A.A.; Buenemann, M.; Chen, R.; Faye, O.; Diagne, C.T.; Faye, O.; Ba, Y.; Dia, I.; Watts, D.; et al. Landscape ecology of sylvatic chikungunya virus and mosquito vectors in southeastern Senegal. PLoS Negl. Trop. Dis. 2012, 6, e1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traore-Lamizana, M.; Zeller, H.; Monlun, E.; Mondo, M.; Hervy, J.P.; Adam, F.; Digoutte, J.P. Dengue 2 outbreak in southeastern Senegal during 1990: Virus isolations from mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 623–627. [Google Scholar] [CrossRef]
- Ndiaye, E.H.; Diallo, D.; Fall, G.; Ba, Y.; Faye, O.; Dia, I.; Diallo, M. Arboviruses isolated from the Barkedji mosquito-based surveillance system, 2012-2013. BMC Infect. Dis. 2018, 18, 642. [Google Scholar] [CrossRef]
- Digoutte, J.P.; Calvo-Wilson, M.A.; Mondo, M.; Traore-Lamizana, M.; Adam, F. Continuous cell lines and immune ascitic fluid pools in arbovirus detection. Res. Virol. 1992, 143, 417–422. [Google Scholar] [CrossRef]
- Li, P.E.; Lo, C.C.; Anderson, J.J.; Davenport, K.W.; Bishop-Lilly, K.A.; Xu, Y.; Ahmed, S.; Feng, S.; Mokashi, V.P.; Chain, P.S. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017, 45, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.M.; et al. Genome Detective: An automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Rambaut, A. Figtree v1.4.4. 2019. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 7 June 2021).
- Phan, M.V.; Murad, S.D.; van der Eijk, A.A.; Metselaar, H.J.; Hartog, H.; Harinck, F.; GeurtsvanKessel, C.H.; Molenkamp, R.; Cotten, M.; Koopmans, M.P. Genomic sequence of yellow fever virus from a Dutch traveller returning from the Gambia-Senegal region, the Netherlands, November 2018. Euro. Surveill. 2019, 24, 1800684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colebunders, R.; Mariage, J.L.; Coche, J.C.; Pirenne, B.; Kempinaire, S.; Hantson, P.; Van Gompel, A.; Niedrig, M.; Van Esbroeck, M.; Bailey, R.; et al. A Belgian traveler who acquired yellow fever in the Gambia. Clin. Infect. Dis. 2002, 35, e113–e116. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.G.; Drosten, C.; Emmerich, P.; Colebunders, R.; Hantson, P.; Pest, S.; Parent, M.; Schmitz, H.; Warnat, M.A.; Niedrig, M. Analysis of two imported cases of yellow fever infection from Ivory Coast and The Gambia to Germany and Belgium. J. Clin. Virol. 2005, 33, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.C.; Soto-Acosta, R.; Bradrick, S.S.; Garcia-Blanco, M.A.; Ooi, E.E. The 5′ and 3′ Untranslated Regions of the Flaviviral Genome. Viruses 2017, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandl, C.W.; Holzmann, H.; Meixner, T.; Rauscher, S.; Stadler, P.F.; Allison, S.L.; Heinz, F.X. Spontaneous and engineered deletions in the 3′ noncoding region of tick-borne encephalitis virus: Construction of highly attenuated mutants of a flavivirus. J. Virol. 1998, 72, 2132–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, M.; Yoshii, K.; Sunden, Y.; Yokozawa, K.; Hirano, M.; Kariwa, H. Variable region of the 3′ UTR is a critical virulence factor in the Far-Eastern subtype of tick-borne encephalitis virus in a mouse model. J. Gen. Virol. 2014, 95, 823–835. [Google Scholar] [CrossRef]
- Proutski, V.; Gritsun, T.S.; Gould, E.A.; Holmes, E.C. Biological consequences of deletions within the 3′-untranslated region of flaviviruses may be due to rearrangements of RNA secondary structure. Virus Res. 1999, 64, 107–123. [Google Scholar] [CrossRef]
- Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Luo, H.; Xie, X.; Medeiros, D.B.A.; Wakamiya, M.; Tesh, R.B.; Barrett, A.D.; Wang, T.; et al. A live-attenuated Zika virus vaccine candidate induces sterilizing immunity in mouse models. Nat. Med. 2017, 23, 763–767. [Google Scholar] [CrossRef]
- Slonchak, A.; Khromykh, A.A. Subgenomic flaviviral RNAs: What do we know after the first decade of research. Antivir. Res. 2018, 159, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, F.M.; Longbottom, J.; Browne, A.J.; Pigott, D.M.; Brady, O.J.; Kraemer, M.U.G.; Marinho, F.; Yactayo, S.; de Araújo, V.E.M.; da Nóbrega, A.A.; et al. Existing and potential infection risk zones of yellow fever worldwide: A modelling analysis. Lancet Glob. Health 2018, 6, e270–e278. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Age | Sex | Locality | Date of Onset of Symptoms (dd/mm/yyyy) | Sampling Date (dd/mm/yyyy) | Symptoms | YF Vaccination Status | PCR YF | Differencial PCR | IgM YF | Differencial ELISA | PRNT90 YF (Titer) | Differencial PRNT90 Assays | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient 1 | 40 | F | Kidira | NA | 18/10/2020 | Fever | Not vaccinated | - | - | + | - | + (1/320) | - |
| Patient 2 | 8 | M | Bakel | 13/10/2020 | 29/10/2020 | Not vaccinated | Not vaccinated | + (ct: 34.9) | - | + | - | + (1/80) | - |
| Patient 3 | 23 | M | Kidira | 02/11/2020 | 04/11/2020 | Fever + jaundice | Not vaccinated | + (ct: 33.4) | - | + | - | + (1/80) | - |
| Patient 4 | 15 | F | Kidira | NA | 02/11/2020 | Headache | Not vaccinated | - | - | + | - | + (1/160) | - |
| Patient 5 | 24 | F | Matam | NA | 19/11/2020 | Fever | Not vaccinated | - | - | + | - | + (1/40) | - |
| Health Districts | No. of Patients | No. of YFV Positive Case (%) | YFV/Malaria Co-Infection (%) | No. of Acute Infection (Positive YFV RT-PCR) | No. of Recently Acquired Infection (Positive YFV Serology Assays *) |
|---|---|---|---|---|---|
| Kedougou | 421 | 13 (3.1%) | 8 (1.9%) | 1 | 12 |
| Saraya | 120 | 4 (3.3%) | 2 (1.7%) | 1 | 3 |
| Sites | Month | Species | No of Mosquitoes Collected | No of Pools Tested | Infected Species (No Positive Pools) |
|---|---|---|---|---|---|
| Kedougou | August | Aedes aegypti | 661 | 97 | Ae. furcifer (6), Ae. luteocephalus (3), Ae. taylori (1) |
| Ae. africanus | 92 | 9 | |||
| Ae. furcifer | 1025 | 108 | |||
| Ae. luteocephalus | 419 | 58 | |||
| Ae. taylori | 112 | 33 | |||
| Ae. vittatus | 495 | 86 | |||
| September | Ae. aegypti | 478 | 65 | Ae. furcifer (7), Ae. luteocephalus (5), Ae. taylori (1), Ae. vittatus (1), Ae. africanus (1) | |
| Ae. africanus | 69 | 11 | |||
| Ae. furcifer | 515 | 74 | |||
| Ae. luteocephalus | 141 | 33 | |||
| Ae. taylori | 25 | 15 | |||
| Ae. vittatus | 339 | 64 | |||
| Kidira | November | Ae. aegypti | 19 | 4 | |
| Ae. africanus | 0 | 0 | |||
| Ae. furcifer | 0 | 0 | |||
| Ae. luteocephalus | 0 | 0 | |||
| Ae. taylori | 0 | 0 | |||
| Ae. vittatus | 0 | 0 | |||
| All | Ae. aegypti | 1158 | 166 | Ae. furcifer (13), Ae. luteocephalus (8), Ae. taylori (2), Ae. vittatus (1), Ae. africanus (1) | |
| Ae. africanus | 161 | 20 | |||
| Ae. furcifer | 1540 | 182 | |||
| Ae. luteocephalus | 560 | 91 | |||
| Ae. taylori | 137 | 48 | |||
| Ae. vittatus | 834 | 150 | |||
| others | 3084 | 511 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diagne, M.M.; Ndione, M.H.D.; Gaye, A.; Barry, M.A.; Diallo, D.; Diallo, A.; Mwakibete, L.L.; Diop, M.; Ndiaye, E.H.; Ahyong, V.; et al. Yellow Fever Outbreak in Eastern Senegal, 2020–2021. Viruses 2021, 13, 1475. https://doi.org/10.3390/v13081475
Diagne MM, Ndione MHD, Gaye A, Barry MA, Diallo D, Diallo A, Mwakibete LL, Diop M, Ndiaye EH, Ahyong V, et al. Yellow Fever Outbreak in Eastern Senegal, 2020–2021. Viruses. 2021; 13(8):1475. https://doi.org/10.3390/v13081475
Chicago/Turabian StyleDiagne, Moussa Moïse, Marie Henriette Dior Ndione, Alioune Gaye, Mamadou Aliou Barry, Diawo Diallo, Amadou Diallo, Lusajo L. Mwakibete, Mamadou Diop, El Hadji Ndiaye, Vida Ahyong, and et al. 2021. "Yellow Fever Outbreak in Eastern Senegal, 2020–2021" Viruses 13, no. 8: 1475. https://doi.org/10.3390/v13081475
APA StyleDiagne, M. M., Ndione, M. H. D., Gaye, A., Barry, M. A., Diallo, D., Diallo, A., Mwakibete, L. L., Diop, M., Ndiaye, E. H., Ahyong, V., Diouf, B., Mhamadi, M., Diagne, C. T., Danfakha, F., Diop, B., Faye, O., Loucoubar, C., Fall, G., Tato, C. M., ... Faye, O. (2021). Yellow Fever Outbreak in Eastern Senegal, 2020–2021. Viruses, 13(8), 1475. https://doi.org/10.3390/v13081475