
Astrovirus in Reunion Free-Tailed Bat (Mormopterus francoismoutoui)

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
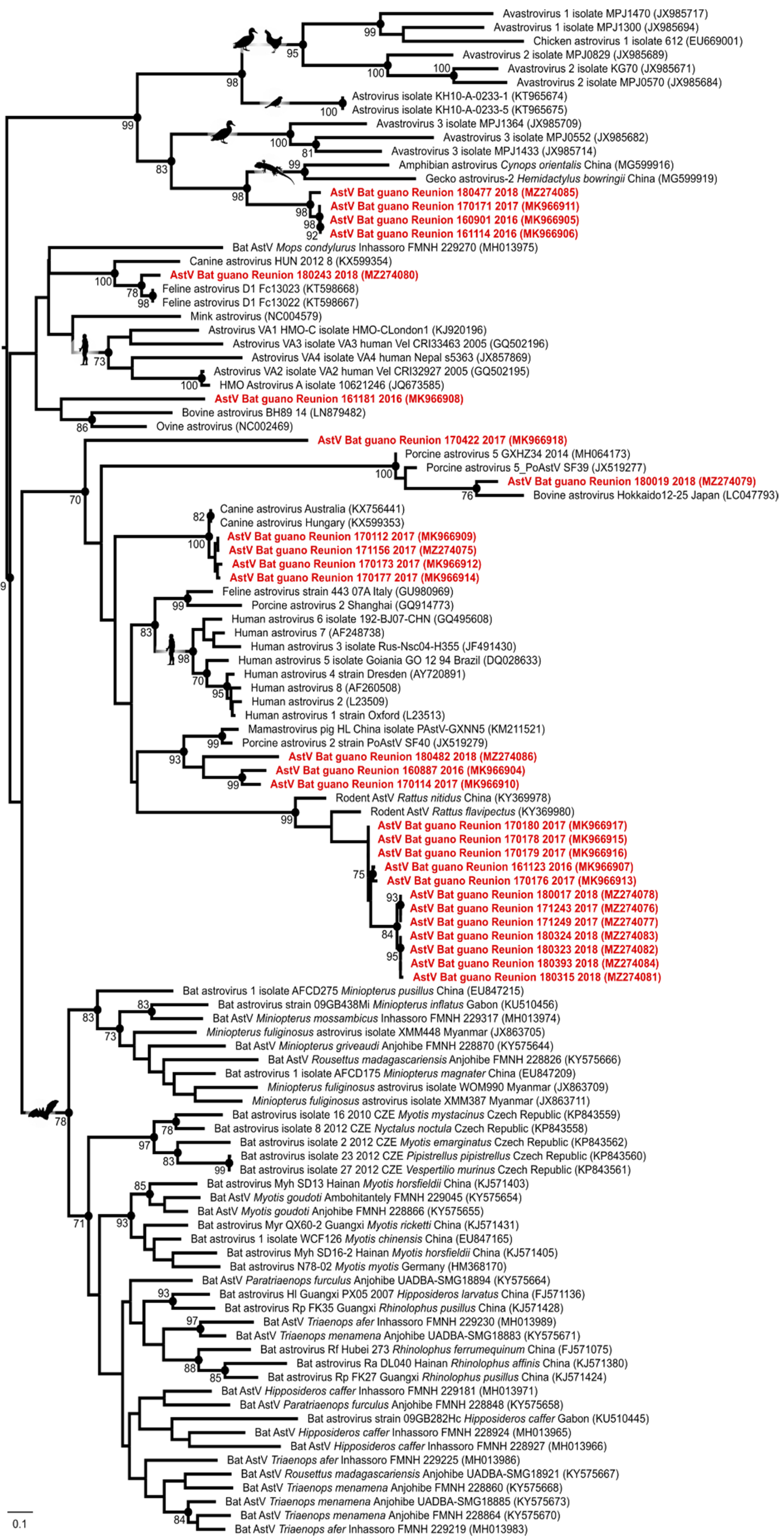
3. Results and Discussion
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals-molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef]
- Bosch, A.; Guix, S.; Krishna, N.K.; Méndez, E.; Monroe, S.S.; Pantin-Jackwood, M.; Schultz-Cherry, S. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E., Eds.; Elsevier: San Diego, CA, USA, 2011. [Google Scholar]
- Fischer, K.; dos Reis, V.P.; Balkema-buschmann, A. Bat Astroviruses: Towards understanding the transmission dynamics of a neglected virus family. Viruses 2017, 9, 34. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Leung, C.Y.H.; Perera, H.K.K.; Ng, E.M.; Gilbert, M.; Joyner, P.H.; Poon, L.L.M. A novel group of avian astroviruses in wild aquatic birds. J. Virol. 2012, 86, 13772–13778. [Google Scholar] [CrossRef] [Green Version]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.D.; Wellehan, J.F.X. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166–173. [Google Scholar] [CrossRef]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, F.A.; San, S.; Schultz-Cherry, S. Non-human primates harbor diverse mammalian and avian astroviruses including those associated with human infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Smith, G.J.D.; Dhanasekaran, V. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; Pinto, R.M.; Guix, S. Human Astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [Green Version]
- Hoarau, F.; Le Minter, G.; Joffrin, L.; Schoeman, M.C.; Lagadec, E.; Ramasindrazana, B.; Lebarbenchon, C. Bat astrovirus in Mozambique. Virol. J. 2018, 15, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Rougeron, V.; Suquet, E.; Maganda, G.D.; Jiolle, D.; Mombo, I.M.; Bourgarel, M.; Leroy, E.M. Characterization and phylogenetic analysis of new bat astroviruses detected in Gabon, Central Africa. Acta Virol. 2016, 60, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Waruhiu, C.; Ommeh, S.; Obanda, V.; Agwanda, B.; Gakuya, F.; Ge, X.Y.; Shi, Z.L. Molecular detection of viruses in Kenyan bats and discovery of novel astroviruses, caliciviruses and rotaviruses. Virol. Sin. 2017, 32, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Yinda, C.K.; Ghogomu, S.M.; Conceição-Neto, N.; Beller, L.; Deboutte, W.; Vanhulle, E.; Matthijnssens, J. Cameroonian fruit bats harbor divergent viruses, including rotavirus H, bastroviruses, and picobirnaviruses using an alternative genetic code. Virus Evol. 2018, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Drexler, J.F.; Corman, V.M.; Wegner, T.; Tateno, A.F.; Zerbinati, R.M.; Gloza-Rausch, F.; Drosten, C. Amplification of emerging viruses in a bat colony. Emerg. Infect. Dis. 2011, 17, 449–456. [Google Scholar] [CrossRef]
- Mendenhall, I.H.; Skiles, M.M.; Neves, E.S.; Borthwick, S.A.; Low, D.H.W.; Liang, B.; Smith, G.J.D. Influence of age and body condition on astrovirus infection of bats in Singapore: An evolutionary and epidemiological analysis. One Health 2017, 4, 27–33. [Google Scholar] [CrossRef]
- Seltmann, A.; Corman, V.; Rasche, A.; Drosten, C.; Czirjak, G.; Bernard, H.; Voigt, C. Seasonal fluctuations of astrovirus, but not coronavirus shedding in bats inhabiting human-modified tropical forests. Ecohealth 2017, 14, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Goodman, S.M. Les Chauves-Souris de Madagascar; Association Vahatra: Antananarivo, Madagascar, 2011. [Google Scholar]
- Goodman, S.M.; Schoeman, M.C.; Rakotoarivelo, A.; Willows-Munro, S. How many species of Hipposideros have occurred on Madagascar since the Late Pleistocene? Zool. J. Linn. Soc. 2016, 177, 428–449. [Google Scholar] [CrossRef] [Green Version]
- Foley, N.M.; Goodman, S.M.; Whelan, C.V.; Puechmaille, S.J.; Teeling, E. Towards navigating the Minotaur’s labyrinth: Cryptic diversity and taxonomic revision within the speciose genus Hipposideros (Hipposideridae). Acta Chiropterologica 2017, 19, 1–18. [Google Scholar] [CrossRef]
- Goodman, S.M.; Jansen van Vuuren, B.; Ratrimomanarivo, F.; Probst, J.; Bowie, R.; Group, E.G.; Re, L. Specific status of populations in the mascarene islands referred to Mormopterus acetabulosus (chiroptera: Molossidae), with description of a new species. J. Mammal. 2008, 89, 1316–1327. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, M.; Wilkinson, D.A.; Benlali, A.; Lagadec, E.; Ramasindrazana, B.; Dellagi, K.; Tortosa, P. Leptospira and paramyxovirus infection dynamics in a bat maternity enlightens pathogen maintenance in wildlife. Environ. Microbiol. 2015, 17, 4280–4289. [Google Scholar] [CrossRef]
- Lebarbenchon, C.; Ramasindrazana, B.; Joffrin, L.; Bos, S.; Lagadec, E.; Le Minter, G.; Mavingui, P. Astroviruses in bats, Madagascar. Emerg. Microbes Infect. 2017, 6, e58. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Keane, T.M.; Creevey, C.J.; Pentony, M.M.; Naughton, T.J.; Mclnerney, J.O. Assessment of methods for amino acid matrix selection and their use on empirical data shows that ad hoc assumptions for choice of matrix are not justified. BMC Evol. Biol. 2006, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Mildenstein, T.; de Jong, C. Natural history, ecological and socio-economic value of bats. In Investigating the Role of Bats in Emerging Zoonoses; Newman, S.H., Field, H., Epstein, J., de Jong, C., Eds.; FAO Animal; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2011; pp. 15–28. [Google Scholar]
- Joffrin, L.; Dietrich, M.; Mavingui, P.; Lebarbenchon, C. Bat pathogens hit the road: But which one? PLoS Pathog. 2018, 14, e1007134. [Google Scholar] [CrossRef]
- Donato, C.; Vijaykrishna, D. The broad host range and genetic diversity of mammalian and avian astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Niu, J.; Wang, H.; Dong, G.; Guo, Y.; Dong, H.; Hu, G. Molecular characterization of feline astrovirus in domestic cats from Northeast China. PLoS ONE 2018, 13, e0205441. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joffrin, L.; Hoarau, A.O.G.; Lagadec, E.; Köster, M.; Ramanantsalama, R.V.; Mavingui, P.; Lebarbenchon, C. Astrovirus in Reunion Free-Tailed Bat (Mormopterus francoismoutoui). Viruses 2021, 13, 1524. https://doi.org/10.3390/v13081524
Joffrin L, Hoarau AOG, Lagadec E, Köster M, Ramanantsalama RV, Mavingui P, Lebarbenchon C. Astrovirus in Reunion Free-Tailed Bat (Mormopterus francoismoutoui). Viruses. 2021; 13(8):1524. https://doi.org/10.3390/v13081524
Chicago/Turabian StyleJoffrin, Léa, Axel O. G. Hoarau, Erwan Lagadec, Marie Köster, Riana V. Ramanantsalama, Patrick Mavingui, and Camille Lebarbenchon. 2021. "Astrovirus in Reunion Free-Tailed Bat (Mormopterus francoismoutoui)" Viruses 13, no. 8: 1524. https://doi.org/10.3390/v13081524
APA StyleJoffrin, L., Hoarau, A. O. G., Lagadec, E., Köster, M., Ramanantsalama, R. V., Mavingui, P., & Lebarbenchon, C. (2021). Astrovirus in Reunion Free-Tailed Bat (Mormopterus francoismoutoui). Viruses, 13(8), 1524. https://doi.org/10.3390/v13081524