
Evaluating the Impact of Anthropogenic Factors on the Dissemination of Contemporary Cosmopolitan, Arctic, and Arctic-like Rabies Viruses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
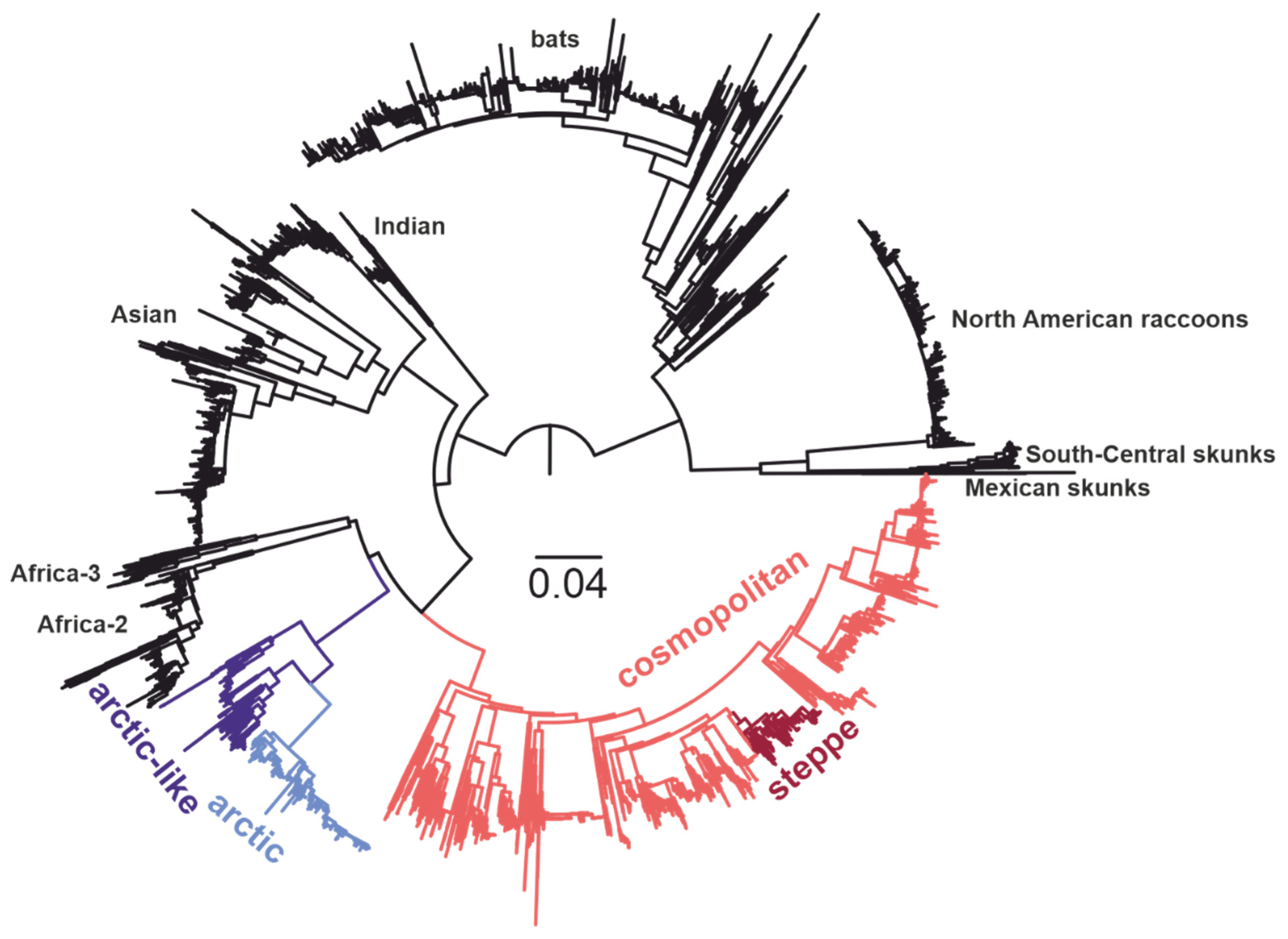
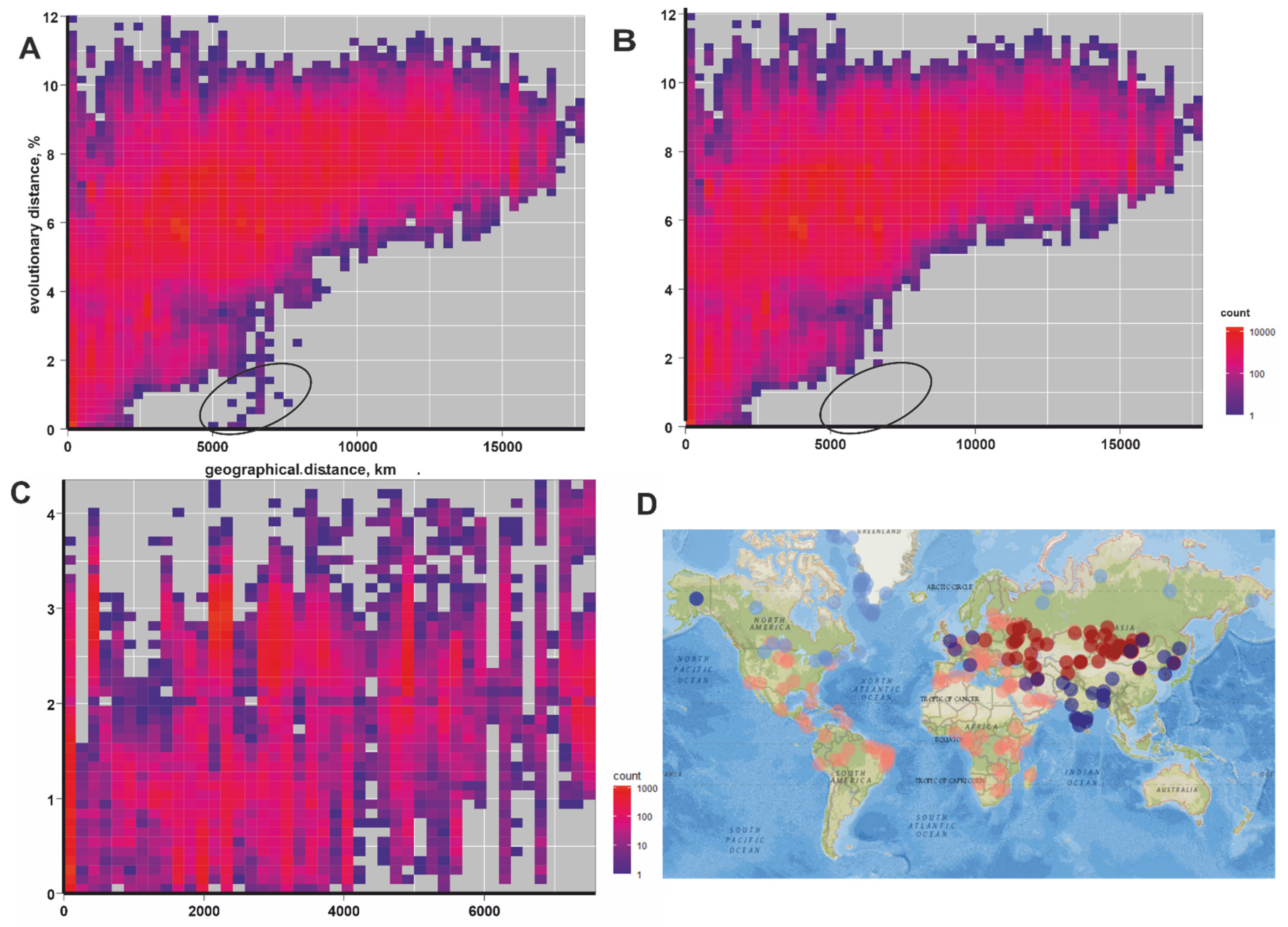
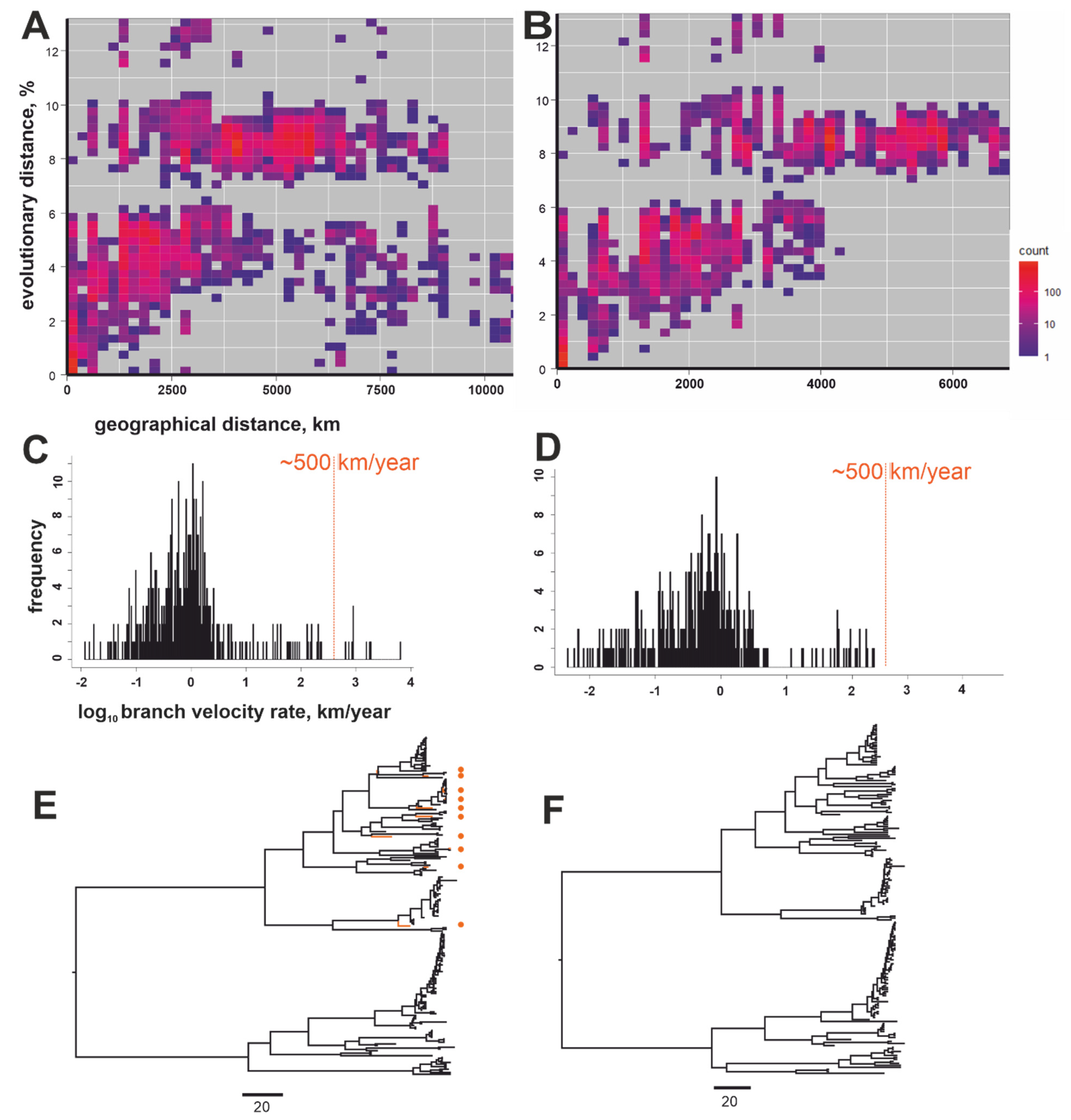
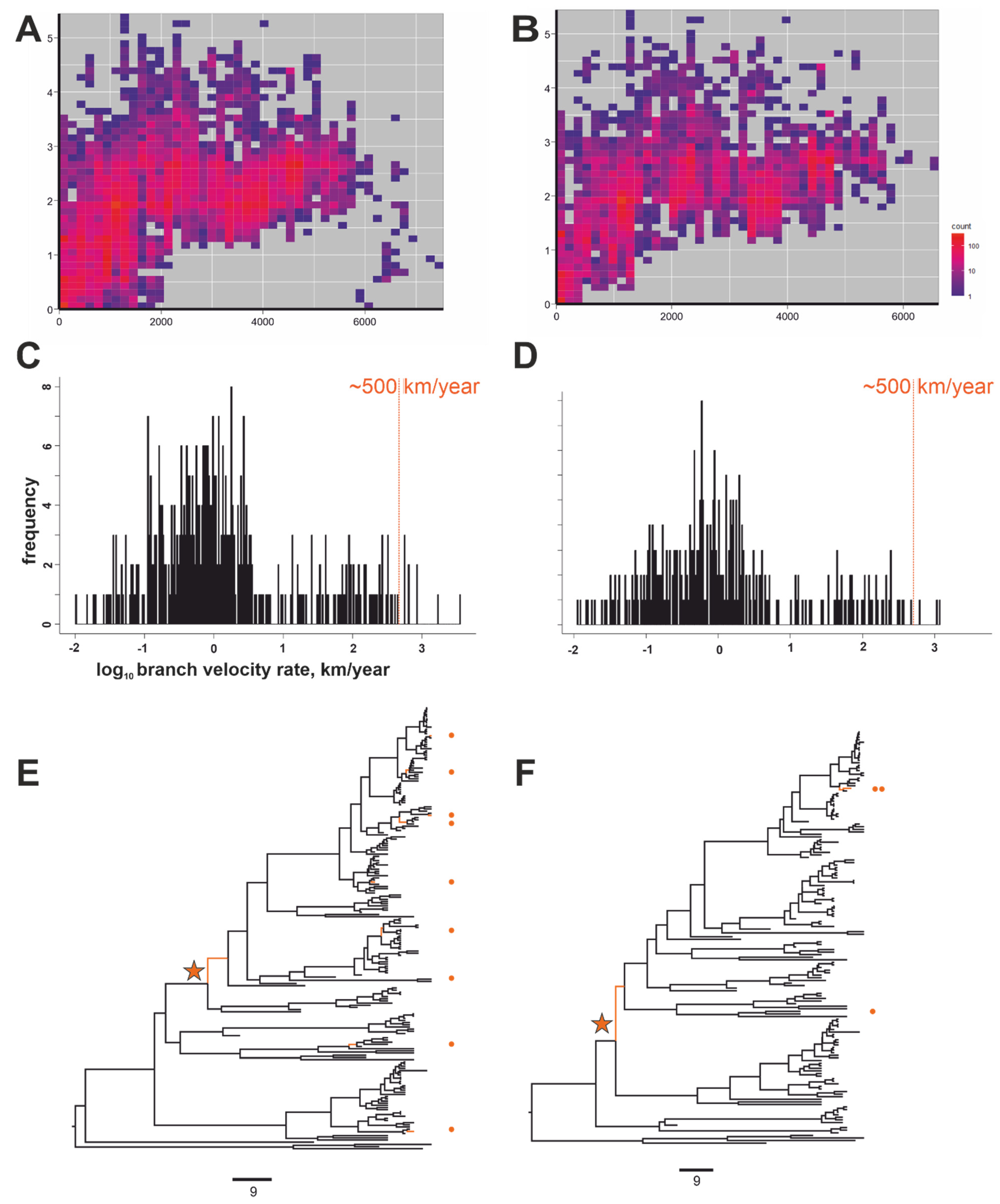
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hayman, D.T.S.; Fooks, A.R.; Marston, D.; Garcia-R, J.C. The Global Phylogeography of Lyssaviruses—Challenging the ’Out of Africa’ Hypothesis. PLoS Negl. Trop. Dis. 2016, 10, e0005266. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Expert Consultation on Rabies: Third Report; WHO Technical Report Series 1012; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Troupin, C.; Dacheux, L.; Tanguy, M.; Sabeta, C.; Blanc, H.; Bouchier, C.; Vignuzzi, M.; Duchene, S.; Holmes, E.; Bourhy, H. Large-Scale Phylogenomic Analysis Reveals the Complex Evolutionary History of Rabies Virus in Multiple Carnivore Hosts. PLoS Pathog. 2016, 12, e1006041. [Google Scholar] [CrossRef] [PubMed]
- Bourhy, H.; Reynes, J.-M.; Dunham, E.J.; Dacheux, L.; Larrous, F.; Huong, V.T.Q.; Xu, G.; Yan, J.; Miranda, M.E.G.; Holmes, E. The origin and phylogeography of dog rabies virus. J. Gen. Virol. 2008, 89, 2673–2681. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Freuling, C.M.; Müller, T.; Pfaff, F.; Bodenhofer, U.; Höper, D.; Fischer, M.; Marston, D.A.; Fooks, A.R.; Mettenleiter, T.C.; et al. Defining objective clusters for rabies virus sequences using affinity propagation clustering. PLoS Negl. Trop. Dis. 2018, 12, e0006182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco-Villa, A.; Mauldin, M.R.; Shi, M.; Escobar, L.E.; Gallardo-Romero, N.F.; Damon, I.; Olson, V.A.; Streicker, D.G.; Emerson, G. The history of rabies in the Western Hemisphere. Antivir. Res. 2017, 146, 221–232. [Google Scholar] [CrossRef]
- Mey, C.; Metlin, A.; Duong, V.; Ong, S.; In, S.; Horwood, P.F.; Reynes, J.-M.; Bourhy, H.; Tarantola, A.; Buchy, P. Evidence of two distinct phylogenetic lineages of dog rabies virus circulating in Cambodia. Infect. Genet. Evol. 2016, 38, 55–61. [Google Scholar] [CrossRef]
- Shchelkanov, M.Y.; Deviatkin, A.; Ananiev, V.Y.; Frolov, E.V.; Dombrovskaya, I.E.; Dedkov, V.G.; Ardashev, A.V.; Kolomeets, S.A.; Korotkova, I.P.; Lyubchenko, E.N.; et al. Isolation and complete genome sequencing of rabies virus strain isolated from a brown bear (Ursus arctos) that attacked a human in Primorsky krai (November 2014). Probl. Virol. 2016, 61, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Boland, T.A.; McGuone, D.; Jindal, J.; Rocha, M.; Cumming, M.; Rupprecht, C.E.; Barbosa, T.F.S.; Oliveira, R.D.N.; Chu, C.J.; Cole, A.J.; et al. Phylogenetic and epidemiologic evidence of multiyear incubation in human rabies. Ann. Neurol. 2013, 75, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Turcitu, M.; Barboi, G.; Vuta, V.; Mihai, I.; Boncea, D.; Dumitrescu, F.; Codreanu, M.; Johnson, N.; Fooks, A.; Müller, T.; et al. Molecular epidemiology of rabies virus in Romania provides evidence for a high degree of heterogeneity and virus diversity. Virus Res. 2010, 150, 28–33. [Google Scholar] [CrossRef]
- Anderson, R.; Jackson, H.C.; May, R.M.; Smith, A.M. Population dynamics of fox rabies in Europe. Nat. Cell Biol. 1981, 289, 765–771. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Botvinkin, A.D.; McElhinney, L.; Smith, J.S.; Orciari, L.A.; Hughes, G.J.; Fooks, A.R.; Rupprecht, C.E. Molecular epidemiology of terrestrial rabies in the former soviet Union. J. Wildl. Dis. 2004, 40, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Fuglei, E.; Tarroux, A. Arctic fox dispersal from Svalbard to Canada: One female’s long run across sea ice. Polar Res. 2019, 38, 3512. [Google Scholar] [CrossRef]
- Nadin-Davis, S.A.; Falardeau, E.; Flynn, A.; Whitney, H.; Marshall, H.D. Relationships between fox populations and rabies virus spread in northern Canada. PLoS ONE 2021, 16, e0246508. [Google Scholar] [CrossRef]
- Joly, K.; Gurarie, E.; Sorum, M.S.; Kaczensky, P.; Cameron, M.D.; Jakes, A.F.; Borg, B.L.; Nandintsetseg, D.; Hopcraft, J.G.C.; Buuveibaatar, B.; et al. Longest terrestrial migrations and movements around the world. Sci. Rep. 2019, 9, 15333. [Google Scholar] [CrossRef]
- Deviatkin, A.A.; Lukashev, A.N.; Poleshchuk, E.; Dedkov, V.G.; Tkachev, S.; Sidorov, G.N.; Karganova, G.G.; Galkina, I.V.; Shchelkanov, M.Y.; Shipulin, G. The phylodynamics of the rabies virus in the Russian Federation. PLoS ONE 2017, 12, e0171855. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, K.; Racloz, V.; McElhinney, L.; Marston, D.; Johnson, N.; Rønsholt, L.; Christensen, L.; Neuvonen, E.; Botvinkin, A.; Rupprecht, C.; et al. Molecular epidemiological study of Arctic rabies virus isolates from Greenland and comparison with isolates from throughout the Arctic and Baltic regions. Virus Res. 2006, 116, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hanke, D.; Freuling, C.M.; Fischer, S.; Hueffer, K.; Hundertmark, K.; Nadin-Davis, S.; Marston, D.; Fooks, A.R.; Bøtner, A.; Mettenleiter, T.C.; et al. Spatio-temporal Analysis of the Genetic Diversity of Arctic Rabies Viruses and Their Reservoir Hosts in Greenland. PLoS Negl. Trop. Dis. 2016, 10, e0004779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmin, I.V.; Hughes, G.J.; Botvinkin, A.D.; Gribencha, S.G.; Rupprecht, C.E. Arctic and Arctic-like rabies viruses: Distribution, phylogeny and evolutionary history. Epidemiol. Infect. 2008, 136, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Nadin-Davis, S.A.; Turner, G.; Paul, J.P.V.; Madhusudana, S.N.; Wandeler, A.I. Emergence of Arctic-like Rabies Lineage in India. Emerg. Infect. Dis. 2007, 13, 111–116. [Google Scholar] [CrossRef]
- Freuling, C.M.; Hampson, K.; Selhorst, T.; Schröder, R.; Meslin, F.X.; Mettenleiter, T.C.; Müller, T. The elimination of fox rabies from Europe: Determinants of success and lessons for the future. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120142. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.; Un, H.; Fooks, A.R.; Freuling, C.; Müller, T.; Aylan, O.; Vos, A. Rabies epidemiology and control in Turkey: Past and present. Epidemiol. Infect. 2009, 138, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Fèvre, E.M.; Bronsvoort, M.; Hamilton, K.A.; Cleaveland, S. Animal movements and the spread of infectious diseases. Trends Microbiol. 2006, 14, 125–131. [Google Scholar] [CrossRef]
- Hutchison, D.W.; Templeton, A.R. Correlation of Pairwise Genetic and Geographic Distance Measures: Inferring the Relative Influences of Gene Flow and Drift on the Distribution of Genetic Variability. Evolution 1999, 53, 1898. [Google Scholar] [CrossRef]
- Deviatkin, A.A.; Kholodilov, I.S.; Belova, O.A.; Bugmyrin, S.V.; Bespyatova, L.A.; Ivannikova, A.Y.; Vakulenko, Y.A.; Lukashev, A.N.; Karganova, G.G. Baltic Group Tick-Borne Encephalitis Virus Phylogeography: Systemic Inconsistency Pattern between Genetic and Geographic Distances. Microorganisms 2020, 8, 1589. [Google Scholar] [CrossRef]
- Faria, N.R.; Suchard, M.A.; Rambaut, A.; Lemey, P. Toward a quantitative understanding of viral phylogeography. Curr. Opin. Virol. 2011, 1, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Talbi, C.; Lemey, P.; Suchard, M.A.; Abdelatif, E.; Elharrak, M.; Jalal, N.; Faouzi, A.; Echevarría, J.E.; Morón, S.V.; Rambaut, A.; et al. Phylodynamics and Human-Mediated Dispersal of a Zoonotic Virus. PLoS Pathog. 2010, 6, e1001166. [Google Scholar] [CrossRef] [Green Version]
- Dellicour, S.; Rose, R.; Faria, N.R.; Vieira, L.F.P.; Bourhy, H.; Gilbert, M.; Lemey, P.; Pybus, O. Using Viral Gene Sequences to Compare and Explain the Heterogeneous Spatial Dynamics of Virus Epidemics. Mol. Biol. Evol. 2017, 34, 2563–2571. [Google Scholar] [CrossRef]
- Yamada, K.D.; Tomii, K.; Katoh, K. Application of the MAFFT sequence alignment program to large data—Reexamination of the usefulness of chained guide trees. Bioinformatics 2016, 32, 3246–3251. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayres, D.L.; Darling, A.; Zwickl, D.J.; Beerli, P.; Holder, M.; Lewis, P.O.; Huelsenbeck, J.P.; Ronquist, F.; Swofford, D.L.; Cummings, M.P.; et al. BEAGLE: An Application Programming Interface and High-Performance Computing Library for Statistical Phylogenetics. Syst. Biol. 2011, 61, 170–173. [Google Scholar] [CrossRef]
- Rambaut, A.; Lam, T.T.; Max Carvalho, L.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed Phylogenetics and Dating with Confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FigTree v1.4.4. Available online: https://github.com/rambaut/figtree/releases (accessed on 28 December 2021).
- Vaiente, M.A.; Scotch, M. Going back to the roots: Evaluating Bayesian phylogeographic models with discrete trait uncertainty. Infect. Genet. Evol. 2020, 85, 104501. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, L.M.; Marston, D.; Freuling, C.M.; Cragg, W.; Stankov, S.; Lalošević, D.; Müller, T.; Fooks, A.R. Molecular diversity and evolutionary history of rabies virus strains circulating in the Balkans. J. Gen. Virol. 2011, 92, 2171–2180. [Google Scholar] [CrossRef] [Green Version]
- Talbi, C.; Holmes, E.; De Benedictis, P.; Faye, O.; Nakouné, E.; Gamatié, D.; Diarra, A.; Elmamy, B.O.; Sow, A.; Adjogoua, E.V.; et al. Evolutionary history and dynamics of dog rabies virus in western and central Africa. J. Gen. Virol. 2009, 90, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Tao, X.; Yin, C.; Han, N.; Yu, J.; Li, H.; Liu, H.; Fang, W.; Adams, J.; Wang, J.; et al. National Borders Effectively Halt the Spread of Rabies: The Current Rabies Epidemic in China Is Dislocated from Cases in Neighboring Countries. PLoS Negl. Trop. Dis. 2013, 7, e2039. [Google Scholar] [CrossRef] [Green Version]
- Biek, R.; Henderson, J.C.; Waller, L.A.; Rupprecht, C.E.; Real, L.A. A high-resolution genetic signature of demographic and spatial expansion in epizootic rabies virus. Proc. Natl. Acad. Sci. USA 2007, 104, 7993–7998. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Ma, J.; Nie, J.; Li, J.; Cao, S.; Wang, L.; Yu, C.; Huang, W.; Li, Y.; Yu, Y.; et al. Antigenic variations of recent street rabies virus. Emerg. Microbes Infect. 2019, 8, 1584–1592. [Google Scholar] [CrossRef]
- Wang, L.; Wu, X.; Bao, J.; Song, C.; Du, J. Phylodynamic and transmission pattern of rabies virus in China and its neighboring countries. Arch. Virol. 2019, 164, 2119–2129. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Villa, A.; Reeder, S.A.; Orciari, L.A.; Yager, P.A.; Franka, R.; Blanton, J.D.; Zuckero, L.; Hunt, P.; Oertli, E.H.; Robinson, L.E.; et al. Enzootic Rabies Elimination from Dogs and Reemergence in Wild Terrestrial Carnivores, United States. Emerg. Infect. Dis. 2008, 14, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.R.; Winkler, W.G. Descriptive epidemiology from an epizootic of raccoon rabies in the middle Atlantic states, 1982–1983. Am. J. Epidemiol. 1987, 126, 429–437. [Google Scholar] [CrossRef]
- Horton, D.; McElhinney, L.M.; Freuling, C.M.; Marston, D.; Banyard, A.C.; Goharrriz, H.; Wise, E.; Breed, A.; Saturday, G.; Kolodziejek, J.; et al. Complex Epidemiology of a Zoonotic Disease in a Culturally Diverse Region: Phylogeography of Rabies Virus in the Middle East. PLoS Negl. Trop. Dis. 2015, 9, e0003569. [Google Scholar] [CrossRef]
- Carnieli, P.; Oliveira, R.D.N.; Macedo, C.I.; Castilho, J.G. Phylogeography of rabies virus isolated from dogs in Brazil between 1985 and 2006. Arch. Virol. 2011, 156, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Brunker, K.; Marston, D.; Horton, D.L.; Cleaveland, S.; Fooks, A.R.; Kazwala, R.; Ngeleja, C.; Lembo, T.; Sambo, M.; Mtema, Z.J.; et al. Elucidating the phylodynamics of endemic rabies virus in eastern Africa using whole-genome sequencing. Virus Evol. 2015, 1, vev011. [Google Scholar] [CrossRef] [Green Version]
- Dibia, I.N.; Sumiarto, B.; Susetya, H.; Putra, A.A.G.; Scott-Orr, H.; Mahardika, G.N. Phylogeography of the current rabies viruses in Indonesia. J. Veter-Sci. 2015, 16, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakulenko, Y.; Deviatkin, A.; Lukashev, A. The Effect of Sample Bias and Experimental Artefacts on the Statistical Phylogenetic Analysis of Picornaviruses. Viruses 2019, 11, 1032. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deviatkin, A.A.; Vakulenko, Y.A.; Dashian, M.A.; Lukashev, A.N. Evaluating the Impact of Anthropogenic Factors on the Dissemination of Contemporary Cosmopolitan, Arctic, and Arctic-like Rabies Viruses. Viruses 2022, 14, 66. https://doi.org/10.3390/v14010066
Deviatkin AA, Vakulenko YA, Dashian MA, Lukashev AN. Evaluating the Impact of Anthropogenic Factors on the Dissemination of Contemporary Cosmopolitan, Arctic, and Arctic-like Rabies Viruses. Viruses. 2022; 14(1):66. https://doi.org/10.3390/v14010066
Chicago/Turabian StyleDeviatkin, Andrei A., Yulia A. Vakulenko, Mariia A. Dashian, and Alexander N. Lukashev. 2022. "Evaluating the Impact of Anthropogenic Factors on the Dissemination of Contemporary Cosmopolitan, Arctic, and Arctic-like Rabies Viruses" Viruses 14, no. 1: 66. https://doi.org/10.3390/v14010066
APA StyleDeviatkin, A. A., Vakulenko, Y. A., Dashian, M. A., & Lukashev, A. N. (2022). Evaluating the Impact of Anthropogenic Factors on the Dissemination of Contemporary Cosmopolitan, Arctic, and Arctic-like Rabies Viruses. Viruses, 14(1), 66. https://doi.org/10.3390/v14010066