
Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
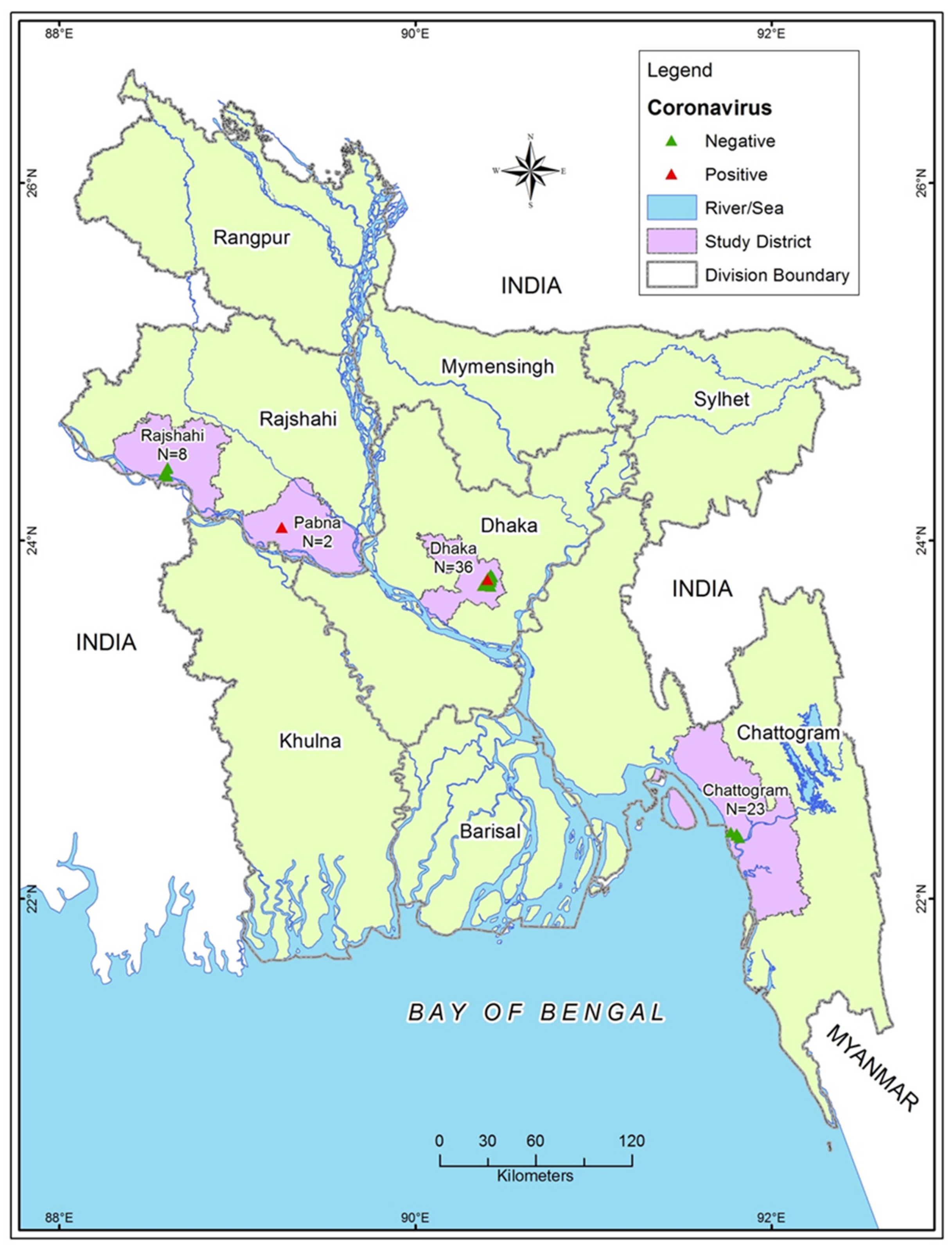
2.1. Study Location and Duration
2.2. Dog Capturing, Sample Collection and Processing, Data Collection
2.3. Laboratory Analysis
2.4. Statistical Analysis
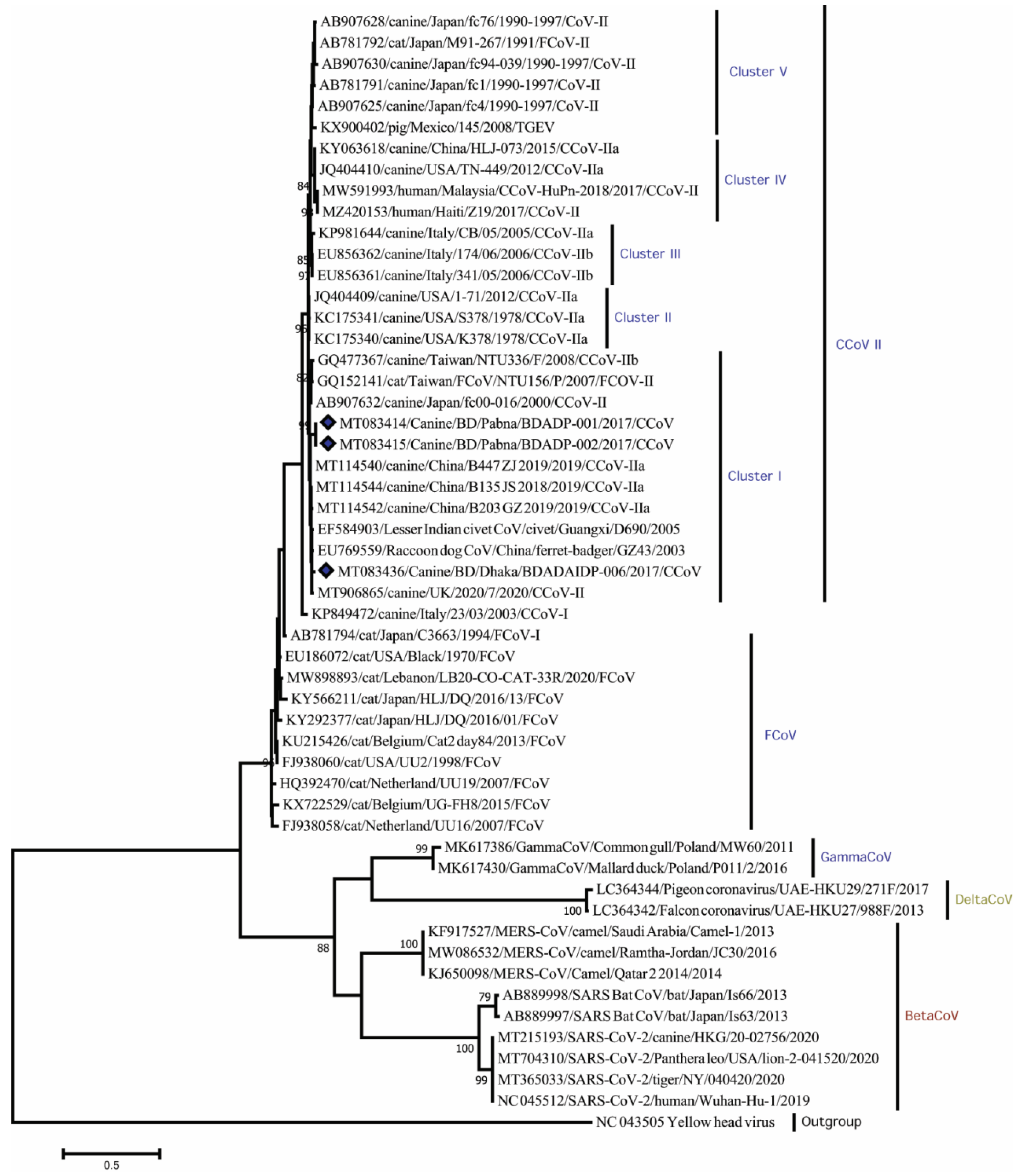
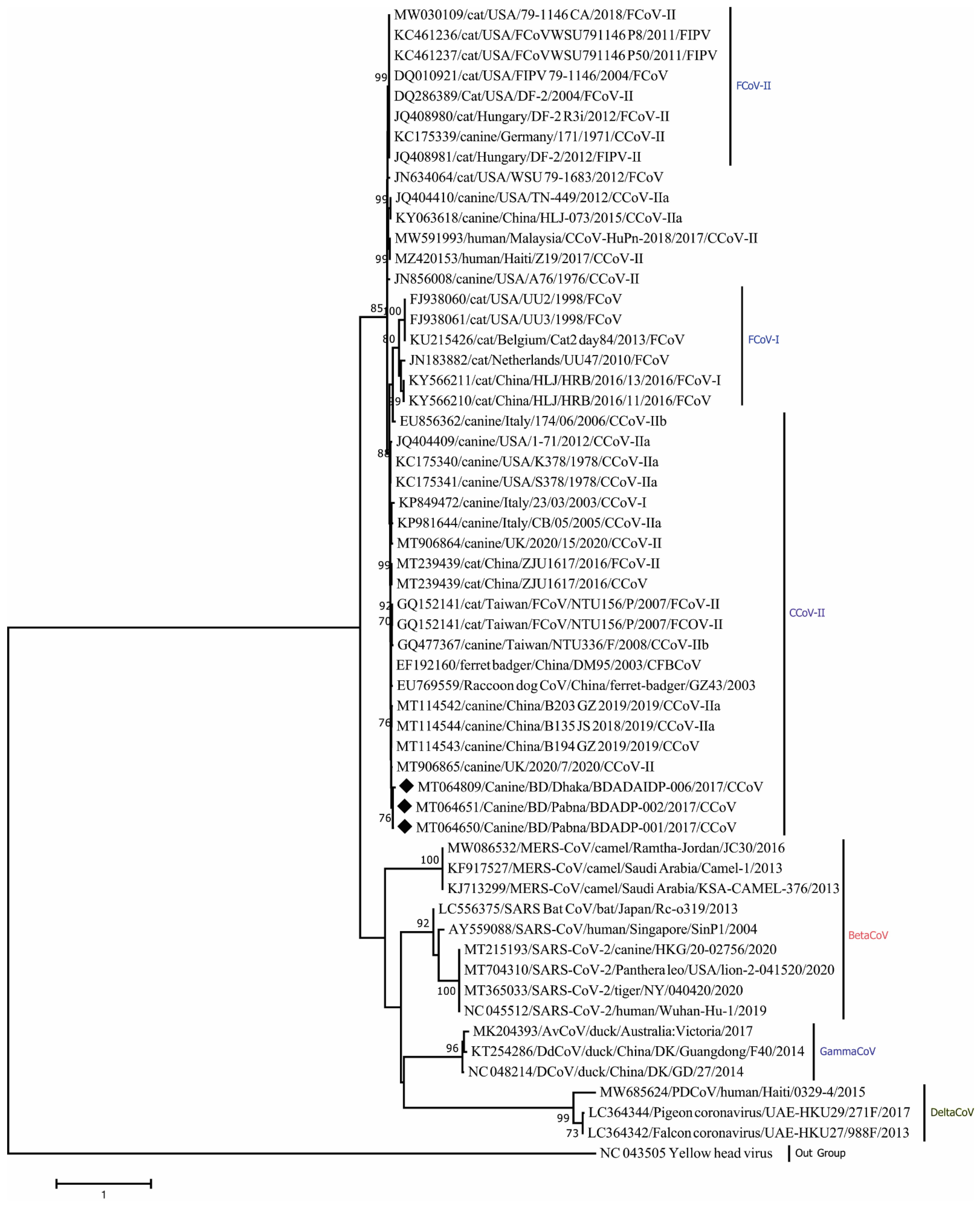
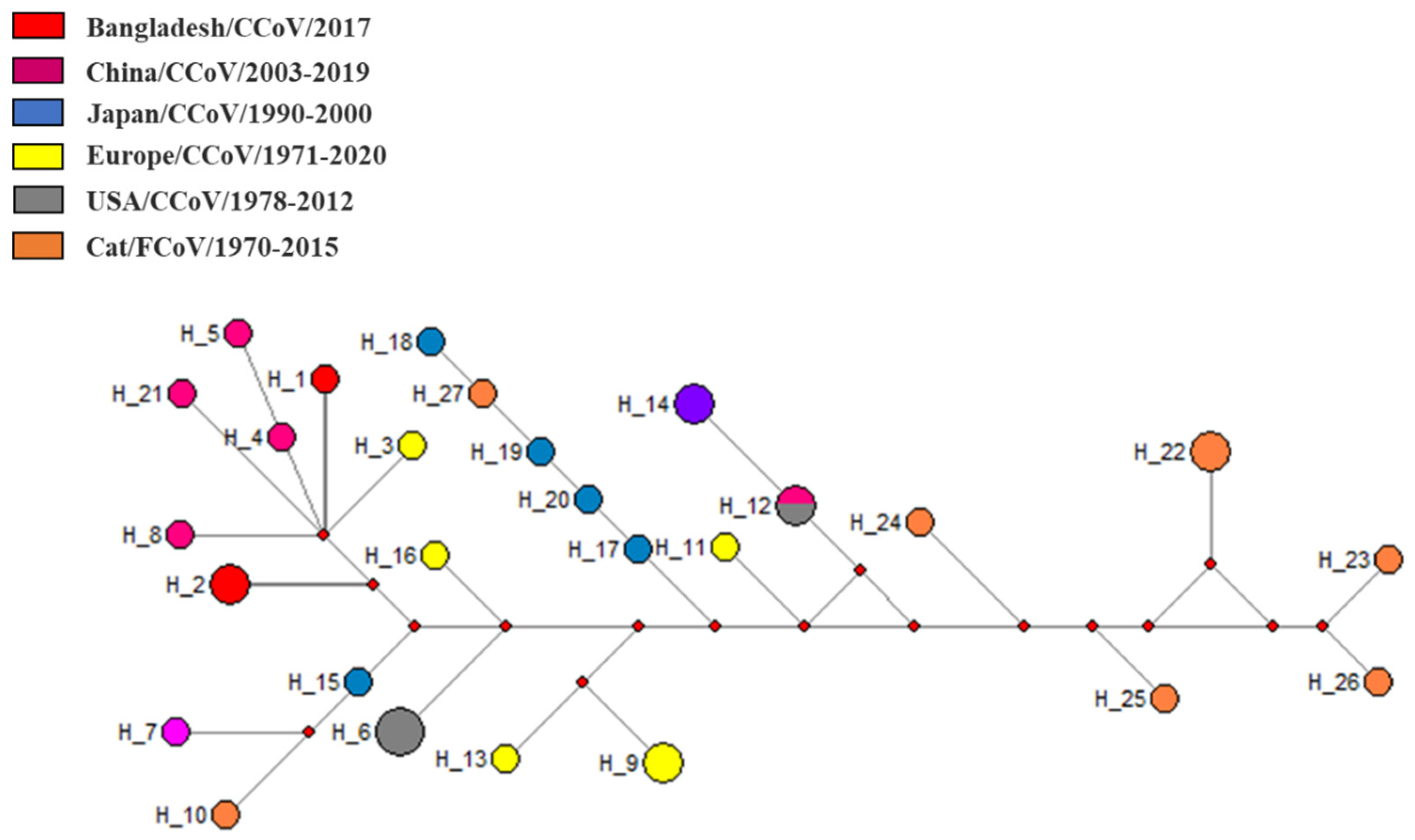
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiss, S.R.; Navas-Martin, S. Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, A.A.; Horzinek, M.C.; Rottier, P.J.; de Groot, R.J. The Genome Organization of the Nidovirales: Similarities and Differences between Arteri-, Toro-, and Coronaviruses. Semin. Virol. 1997, 8, 33–47. [Google Scholar] [CrossRef] [PubMed]
- de Groot, R.J.; Baker, S.C.; Baric, R.; Enjuanes, L.; Gorbalenya, A.E.; Holmes, K.V.; Perlman, S.; Poon, L.; Rottier, P.J.M.; Talbot, P.J.; et al. Virus taxonomy: Classification and nomenclature of viruses. In Ninth Report of the International Committee on Taxonomy of Viruses; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; pp. 806–828. [Google Scholar]
- Haake, C.; Cook, S.; Pusterla, N.; Murphy, B. Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses 2020, 12, 1023. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Sayeed, M.A.; Kalam, M.A.; Ferdous, J.; Rahman, M.K.; Abedin, J.; Islam, S.; Shano, S.; Otun Saha, O.; Tahmina Shirin, T.; et al. Molecular epidemiology of SARS-CoV-2 in diverse environmental samples globally. Microorganisms 2021, 9, 1696. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Ferdous, J.; Sayeed, A.; Rahman, K.; Saha, O.; Hassan, M.M.; Shirin, T. Transmission dynamics and susceptibility patterns of SARS-CoV-2 in domestic, farmed and wild animals: Sustainable One Health surveillance for conservation and public health to prevent future epidemics and pandemics. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhao, K.; Shi, Z.-L.; Zhou, P. Bat Coronaviruses in China. Viruses 2019, 11, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sit, T.H.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T.; et al. Evidence of exposure to SARS-CoV-2 in cats and dogs from households in Italy. Nat. Commun. 2020, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Ferdous, J.; Islam, S.; Sayeed, A.; Choudhury, S.D.; Saha, O.; Hassan, M.M.; Shirin, T. Evolutionary Dynamics and Epidemiology of Endemic and Emerging Coronaviruses in Humans, Domestic Animals, and Wildlife. Viruses 2021, 13, 1908. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Diaz, A.; Damtie, D.; Xiu, L.; Toh, T.-H.; Lee, J.S.-Y.; Saif, L.J.; Gray, G.C. Novel Canine Coronavirus Isolated from a Hospitalized Pneumonia Patient, East Malaysia. Clin. Infect. Dis. 2021, ciab456. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Blohm, G.M.; Alam, M.; Iovine, N.M.; Salemi, M.; Mavian, C.; Morris, J.G. Isolation of a Novel Recombinant Canine Coronavirus from a Visitor to Haiti: Further Evidence of Transmission of Coronaviruses of Zoonotic Origin to Humans. Clin. Infect. Dis. 2021, ciab924. [Google Scholar] [CrossRef] [PubMed]
- Licitra, B.N.; Duhamel, G.E.; Whittaker, G.R. Canine Enteric Coronaviruses: Emerging Viral Pathogens with Distinct Recombinant Spike Proteins. Viruses 2014, 6, 3363–3376. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, A.; Martella, V.; Decaro, N.; Tinelli, A.; Camero, M.; Cirone, F.; Elia, G.; Cavalli, A.; Corrente, M.; Greco, G.; et al. Genetic diversity of a canine coronavirus detected in pups with diarrhoea in Italy. J. Virol. Methods 2003, 110, 9–17. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: Viral evolution and pathobiology. Vet. Microbiol. 2008, 132, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Zappulli, V.; Caliari, D.; Cavicchioli, L.; Tinelli, A.; Castagnaro, M. Systemic fatal type II coronavirus infection in a dog: Pathological findings and immunohistochemistry. Res. Vet. Sci. 2007, 84, 278–282. [Google Scholar] [CrossRef]
- Decaro, N.; Martella, V.; Ricci, D.; Elia, G.; Desario, C.; Campolo, M.; Cavaliere, N.; Di Trani, L.; Tempesta, M.; Buonavoglia, C. Genotype-specific fluorogenic RT-PCR assays for the detection and quantitation of canine coronavirus type I and type II RNA in faecal samples of dogs. J. Virol. Methods 2005, 130, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, A.; Elia, G.; Martella, V.; Tinelli, A.; DeCaro, N.; Marsilio, F.; Buonavoglia, D.; Tempesta, M.; Buonavoglia, C. M gene evolution of canine coronavirus in naturally infected dogs. Vet. Rec. 2002, 151, 758–761. [Google Scholar]
- Decaro, N.; Desario, C.; Elia, G.; Mari, V.; Lucente, M.S.; Cordioli, P.; Colaianni, M.L.; Martella, V.; Buonavoglia, C. Serological and molecular evidence that canine respiratory coronavirus is circulating in Italy. Vet. Microbiol. 2007, 121, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Yachi, A.; Mochizuki, M. Survey of Dogs in Japan for Group 2 Canine Coronavirus Infection. J. Clin. Microbiol. 2006, 44, 2615–2618. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Yamashita, S.; Murata-Ohkubo, M.; Satoh, K.; Doki, T.; Hohdatsu, T. Prevalence of canine coronavirus (CCoV) in dog in Japan: Detection of CCoV RNA and retrospective serological analysis. J. Vet. Med Sci. 2016, 78, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erles, K.; Brownlie, J. Sequence analysis of divergent canine coronavirus strains present in a UK dog population. Virus Res. 2009, 141, 21–25. [Google Scholar] [CrossRef]
- Rahaman, K. Free roaming dogs: A threat to public health. Int. J. Epidemiol. Res. 2017, 4, 182–184. [Google Scholar] [CrossRef]
- Hossain, M.; Ahmed, K.; Marma, A.S.P.; Hossain, S.; Ali, M.A.; Shamsuzzaman, A.K.M.; Nishizono, A. A survey of the dog population in rural Bangladesh. Prev. Vet. Med. 2013, 111, 134–138. [Google Scholar] [CrossRef]
- –Tenzin, T.; Ahmed, R.; Debnath, N.C.; Ahmed, G.; Yamage, M. Free-Roaming Dog Population Estimation and Status of the Dog Population Management and Rabies Control Program in Dhaka City, Bangladesh. PLoS Negl. Trop. Dis. 2015, 9, e0003784. [Google Scholar] [CrossRef] [Green Version]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for Camel-to-Human Transmission of MERS Coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef]
- Bolles, M.; Donaldson, E.; Baric, R. SARS-CoV and emergent coronaviruses: Viral determinants of interspecies transmission. Curr. Opin. Virol. 2011, 1, 624–634. [Google Scholar] [CrossRef]
- Navarro, R.; Nair, R.; Peda, A.; Aung, M.S.; Ashwinie, G.S.; Gallagher, C.A.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Molecular characterization of canine parvovirus and canine enteric coronavirus in diarrheic dogs on the island of St. Kitts: First report from the Caribbean region. Virus Res. 2017, 240, 154–160. [Google Scholar] [CrossRef] [PubMed]
- He, H.-J.; Zhang, W.; Liang, J.; Lu, M.; Wang, R.; Li, G.; He, J.-W.; Chen, J.; Xing, G.; Chen, Y. Etiology and genetic evolution of canine coronavirus circulating in five provinces of China, during 2018–2019. Microb. Pathog. 2020, 145, 104209. [Google Scholar] [CrossRef] [PubMed]
- Haun, B.K.; Kamara, V.; Dweh, A.S.; Garalde-Machida, K.; Forkay, S.S.E.; Takaaze, M.; Namekar, M.; Wong, T.A.S.; Woto, A.E.R.B.-G.; Humphreys, P.; et al. Serological evidence of Ebola virus exposure in dogs from affected communities in Liberia: A preliminary report. PLoS Negl. Trop. Dis. 2019, 13, e0007614. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.; Kendall, A.; Url, A.; Auer, A.; Leschnik, M. The first RT-qPCR confirmed case of tick-borne encephalitis in a dog in Scandinavia. Acta Vet. Scand. 2020, 62, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Montagnaro, S.; Piantedosi, D.; Ciarcia, R.; Loponte, R.; Veneziano, V.; Fusco, G.; Amoroso, M.G.; Ferrara, G.; Damiano, S.; Iovane, G.; et al. Serological Evidence of Mosquito-Borne Flaviviruses Circulation in Hunting Dogs in Campania Region, Italy. Vector-Borne Zoonotic Dis. 2019, 19, 142–147. [Google Scholar] [CrossRef]
- Evermann, J.F.; Lincoln, J.D.; McKiernan, A.J. Isolation of a paramyxovirus from the cerebrospinal fluid of a dog with posterior paresis. J. Am. Vet. Med Assoc. 1980, 177, 1132–1134. [Google Scholar]
- Macartney, L.; Cornwell, H.J.; McCandlish, I.A.; Thompson, H. Isolation of a novel paramyxovirus from a dog with enteric disease. Vet. Record 1985, 117, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Kempf, C.; Schulz, B.S.; Strauch, C.; Sauter-Louis, C.; Truyen, U.; Hartmann, K. Virus detection, clinical signs, and laboratory findings in dogs with acute hemorrhagic diarrhea: A retrospective study of 935 cases. Tierarztl Prax Ausg K Kleintiere Heimtiere 2010, 38, 79–86. [Google Scholar] [PubMed]
- Islam, A.; Rahman, M.L.; Islam, S.; Debnath, P.; Alam, M.; Hassan, M.M. Sero-prevalence of visceral leishmaniasis (VL) among dogs in VL endemic areas of Mymensingh district, Bangladesh. J. Adv. Vet. Anim. Res. 2017, 4, 241–248. [Google Scholar] [CrossRef]
- Khan, S.; Epstein, J.; Olival, K.; Hassan, M.; Hossain, M.; Rahman, K.; Elahi, M.; Mamun, M.; Haider, N.; Yasin, G.; et al. Hematology and serum chemistry reference values of stray dogs in Bangladesh. Open Vet. J. 2011, 1, 13–20. [Google Scholar] [PubMed]
- Rakib, T.M.; Islam, S.; Azam, N.E.; Islam, S.; Al Faruq, A.; Das, T.; Akter, L.; Hassan, M.M.; Hossain, M.A. Multidrug Resistance Pattern of Salmonella Typhimurium Isolated from Rectal Swabs of Stray Dogs at Chittagong Metropolitan Area (CMA), Bangladesh. Microbiol. Res. J. Int. 2018, 25, 1–11. [Google Scholar] [CrossRef]
- Fábrega, L.; Restrepo, C.M.; Torres, A.; Smith, D.; Chan, P.; Pérez, D.; Cumbrera, A.; Caballero, E.Z. Frequency of Toxoplasma gondii and Risk Factors Associated with the Infection in Stray Dogs and Cats of Panama. Microorganisms 2020, 8, 927. [Google Scholar] [CrossRef]
- Burns, K.; Renda-Francis, L. Textbook for the Veterinary Assistant; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Gorrel, C. Veterinary Dentistry for the General Practitioner; Saunders Ltd.: St. Louis, MO, USA, 2014. [Google Scholar] [CrossRef]
- Smith, L.M. The Impact of Dog Population Management on Free-Roaming Dog Population Dynamics, Health and Welfare; University of Leeds: Leeds, UK, 2020. [Google Scholar]
- Moureau, G.; Temmam, S.; Gonzalez, J.-P.; Charrel, R.; Grard, G.; De Lamballerie, X. A Real-Time RT-PCR Method for the Universal Detection and Identification of Flaviviruses. Vector-Borne Zoonotic Dis. 2007, 7, 467–478. [Google Scholar] [CrossRef]
- Zhai, J.; Palacios, G.; Towner, J.S.; Jabado, O.; Kapoor, V.; Venter, M.; Grolla, A.; Briese, T.; Paweska, J.; Swanepoel, R.; et al. Rapid Molecular Strategy for Filovirus Detection and Characterization. J. Clin. Microbiol. 2007, 45, 224–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Chern, S.-W.W.; Li, Y.; Pallansch, M.A.; Anderson, L.J. Sensitive and Broadly Reactive Reverse Transcription-PCR Assays to Detect Novel Paramyxoviruses. J. Clin. Microbiol. 2008, 46, 2652–2658. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.-L.; Firth, C.; Street, C.; Henriquez, J.A.; Petrosov, A.; Tashmukhamedova, A.; Hutchison, S.K.; Egholm, M.; Osinubi, M.O.V.; Niezgoda, M.; et al. Identification of a Severe Acute Respiratory Syndrome Coronavirus-Like Virus in a Leaf-Nosed Bat in Nigeria. mBio 2010, 1, e00208-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Masangkay, J.S.; Nagata, N.; Morikawa, S.; Mizutani, T.; Fukushi, S.; Alviola, P.; Omatsu, T.; Ueda, N.; Iha, K.; et al. Bat Coronaviruses and Experimental Infection of Bats, the Philippines. Emerg. Infect. Dis. 2010, 16, 1217–1223. [Google Scholar] [CrossRef]
- Anthony, S.J.; Johnson, C.K.; Greig, D.J.; Kramer, S.; Che, X.; Wells, H.; Hicks, A.L.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; et al. Global patterns in coronavirus diversity. Virus Evol. 2017, 3, vex012. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Hasegawa, M.; Kishino, H.; Yano, T.-A. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Ntafis, V.; Mari, V.; Decaro, N.; Papanastassopoulou, M.; Pardali, D.; Rallis, T.S.; Kanellos, T.; Buonavoglia, C.; Xylouri, E. Canine coronavirus, Greece. Molecular analysis and genetic diversity characterization. Infect. Genet. Evol. 2013, 16, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Bandai, C.; Ishiguro, S.; Masuya, N.; Hohdatsu, T.; Mochizuki, M. Canine Coronavirus Infections in Japan: Virological and Epidemiological Aspects. J. Vet. Med. Sci. 1999, 61, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeCaro, N.; Mari, V.; Elia, G.; Addie, D.D.; Camero, M.; Lucente, M.S.; Martella, V.; Buonavoglia, C. Recombinant Canine Coronaviruses in Dogs, Europe. Emerg. Infect. Dis. 2010, 16, 41–47. [Google Scholar] [CrossRef]
- Yesilbag, K.; Yilmaz, Z.; Torun, S.; Pratelli, A. Canine Coronavirus Infection in Turkish Dog Population. J. Vet. Med. Ser. B 2004, 51, 353–355. [Google Scholar] [CrossRef]
- Lu, S.; Zhang, D.; Zhou, J.; Xia, Z.; Lin, D.; Lou, Y.; Tan, W.; Qin, K. Prevalence and phylogenetic characterization of canine coronavirus from diseased pet dogs in Beijing, China. Sci. China Life Sci. 2016, 59, 860–862. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, C.; Guo, D.; Wei, S.; Geng, Y.; Wang, E.; Wang, Z.; Zhao, X.; Su, M.; Liu, Q.; et al. Co-Circulation of Canine Coronavirus I and IIa/b with High Prevalence and Genetic Diversity in Heilongjiang Province, Northeast China. PLoS ONE 2016, 11, e0146975. [Google Scholar] [CrossRef] [PubMed]
- Sakulwira, K.; Vanapongtipagorn, P.; Theamboonlers, A.; Oraveerakul, K.; Poovorawan, Y. Prevalence of canine coronavirus and parvovirus infections in dogs with gastroenteritis in Thailand. Vet. Med. 2003, 48, 163–168. [Google Scholar] [CrossRef]
- Terada, Y.; Matsui, N.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Soma, T.; Mochizuki, M.; Maeda, K. Emergence of Pathogenic Coronaviruses in Cats by Homologous Recombination between Feline and Canine Coronaviruses. PLoS ONE 2014, 9, e106534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.L.; Thi, H.H.P. Genome-wide comparison of coronaviruses derived from veterinary animals: A canine and feline perspective. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101654. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-N.; Chang, R.-Y.; Su, B.-L.; Chueh, L.-L. Full genome analysis of a novel type II feline coronavirus NTU156. Virus Genes 2013, 46, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Mullis, L.B.; Pereira, O.; Saif, L.J.; Vlasova, A.; Zhang, X.; Owens, R.J.; Paulson, D.; Taylor, D.; Haynes, L.M.; et al. Human Respiratory Coronaviruses Detected In Patients with Influenza-Like Illness in Arkansas, USA. Virol. Mycol. 2014, 2014, 004. [Google Scholar] [CrossRef] [PubMed]
- Xiu, L.; Binder, R.A.; Alarja, N.A.; Kochek, K.; Coleman, K.K.; Than, S.T.; Bailey, E.S.; Bui, V.N.; Toh, T.-H.; Erdman, D.D.; et al. A RT-PCR assay for the detection of coronaviruses from four genera. J. Clin. Virol. 2020, 128, 104391. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, G.; Marshall, J.; Morrell, J.; Robb, D.; McCauley, J.W.; Perez, D.R.; Parrish, C.R.; Murcia, P.R. Infection and Pathogenesis of Canine, Equine, and Human Influenza Viruses in Canine Tracheas. J. Virol. 2014, 88, 9208–9219. [Google Scholar] [CrossRef] [Green Version]
- Delong, J. Dutch Minister Confirms Dog, Three Cats Have Caught Novel Coronavirus|Reuters. 2020. Available online: https://www.reuters.com/article/us-health-coronavirus-netherlands-pets-idUSKBN22R2EN (accessed on 17 September 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Categories | N | CCoV Positive n (%) | 95% CI |
|---|---|---|---|---|
| District | Chattogram | 23 | - | - |
| Dhaka | 36 | 1 (2.8) | 0.07–14.5 | |
| Pabna | 2 | 2 (100) | 15.8–100 | |
| Rajshahi | 8 | - | - | |
| Sampling time | 2009–2010 | 53 | - | - |
| 2016–2017 | 16 | 3 (18.8) | 4.0–45.6 | |
| Sampling season | Dry | 16 | 3 (18.8) | 4.0–45.6 |
| Wet | 53 | - | - | |
| Sex | Male | 36 | 1 (2.8) | 0.07–14.5 |
| Female | 33 | 2 (6.1) | 0.7–20.2 | |
| Age | 1–1.5 years | 32 | - | - |
| 1.6–3 years | 24 | - | - | |
| >3 years | 13 | 3 (23.1) | 5.0–53.8 | |
| Type of ownership | Stray dog | 58 | 2 (3.5) | 0.4–11.9 |
| Sheltered dog | 11 | 1 (9.1) | 0.2–41.2 | |
| Health condition | Sick | 13 | 1 (7.7) | 0.2–36.0 |
| Apparently healthy | 56 | 2 (3.6) | 0.4–12.3 | |
| BCS | Poor | 22 | 2 (9.1) | 1.1–29.1 |
| Fair | 29 | 1 (3.5) | 0.08–17.8 | |
| Good | 18 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.E.; Islam, A.; Islam, S.; Rahman, M.K.; Miah, M.; Alam, M.S.; Rahman, M.Z. Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh. Viruses 2022, 14, 67. https://doi.org/10.3390/v14010067
Hossain ME, Islam A, Islam S, Rahman MK, Miah M, Alam MS, Rahman MZ. Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh. Viruses. 2022; 14(1):67. https://doi.org/10.3390/v14010067
Chicago/Turabian StyleHossain, Mohammad Enayet, Ariful Islam, Shariful Islam, Md Kaisar Rahman, Mojnu Miah, Md Shaheen Alam, and Mohammed Ziaur Rahman. 2022. "Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh" Viruses 14, no. 1: 67. https://doi.org/10.3390/v14010067
APA StyleHossain, M. E., Islam, A., Islam, S., Rahman, M. K., Miah, M., Alam, M. S., & Rahman, M. Z. (2022). Detection and Molecular Characterization of Canine Alphacoronavirus in Free-Roaming Dogs, Bangladesh. Viruses, 14(1), 67. https://doi.org/10.3390/v14010067