
An Update on Detection Technologies for SARS-CoV-2 Variants of Concern



 ,
,  and
and
Abstract
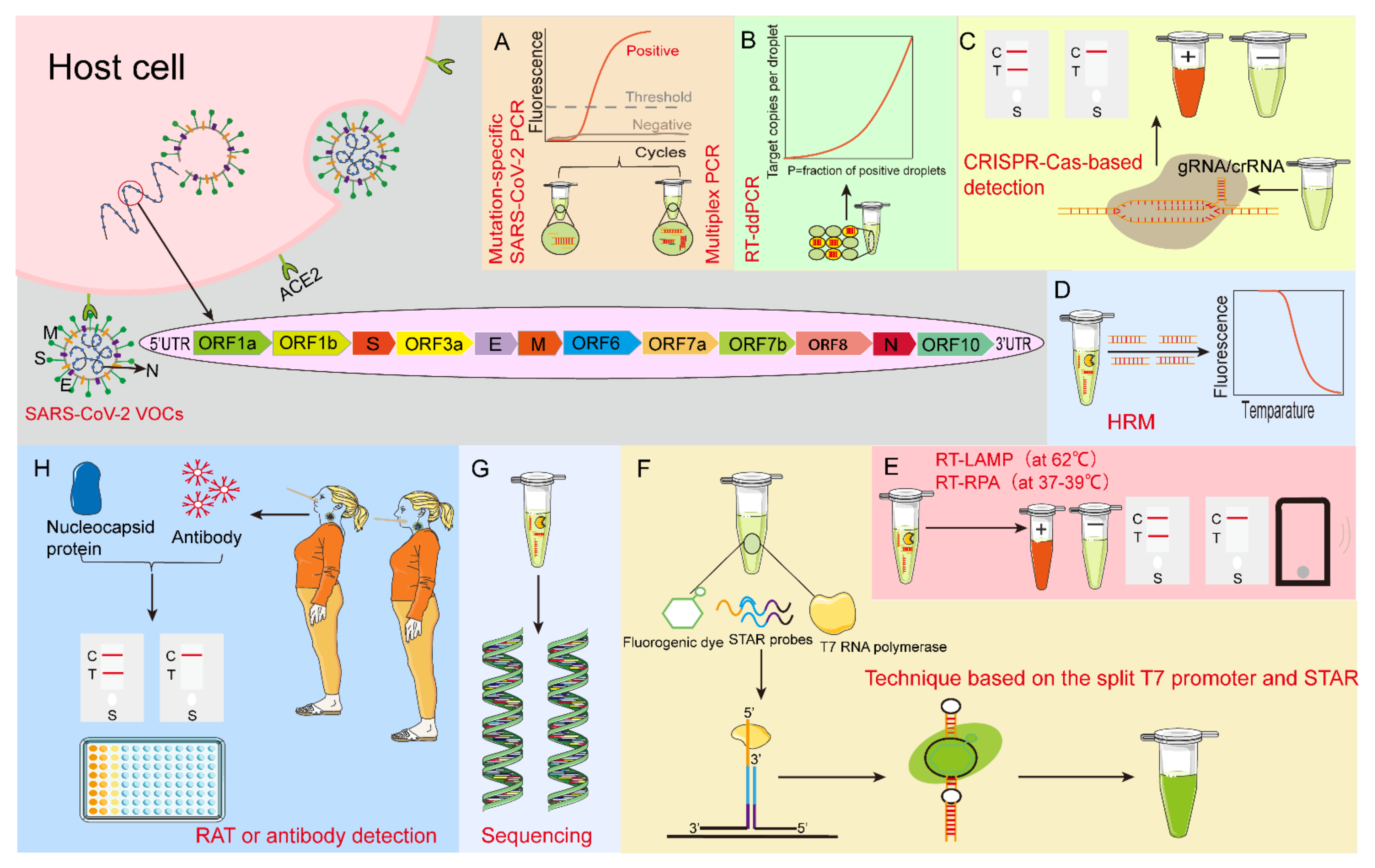
:1. Introduction
2. Nucleic Acid Detection-Based Test
2.1. Mutation-Specific SARS-CoV-2 PCR
2.2. Multiplex PCR
2.3. LAMP Assay
2.4. CRISPR-Cas-Based Detection Technology
2.5. Other Nucleic Acid Detection-Based Test
3. Viral Genome Sequencing
3.1. Generation Sequencing: Sanger Sequencing
3.2. NGS: Illumina Sequencing
3.3. Third Generation Sequencing: Nanopore Sequencing
4. Protein-Based SARS-CoV-2 Variants Detection
4.1. Antigen-Based Detection
4.2. Antibody-Based Detection
5. Combined Detection Technology
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants/ (accessed on 31 May 2021).
- Dispinseri, S.; Marzinotto, I.; Brigatti, C.; Pirillo, M.F.; Tolazzi, M.; Bazzigaluppi, E.; Canitano, A.; Borghi, M.; Gallinaro, A.; Caccia, R.; et al. Seasonal Betacoronavirus Antibodies’ Expansion Post-BNT161b2 Vaccination Associates with Reduced SARS-CoV-2 VoC Neutralization. J. Clin. Immunol. 2022, 42, 448–458. [Google Scholar] [CrossRef]
- Rubin, R. COVID-19 Vaccines vs Variants—Determining How Much Immunity Is Enough. JAMA 2021, 325, 1241–1243. [Google Scholar] [CrossRef]
- Filchakova, O.; Dossym, D.; Ilyas, A.; Kuanysheva, T.; Abdizhamil, A.; Bukasov, R. Review of COVID-19 testing and diagnostic methods. Talanta 2022, 244, 123409. [Google Scholar] [CrossRef]
- Ko, K.K.K.; Abdul Rahman, N.B.; Tan, S.Y.L.; Chan, K.X.L.; Goh, S.S.; Sim, J.H.C.; Lim, K.L.; Tan, W.L.; Chan, K.S.; Oon, L.L.E.; et al. SARS-CoV-2 N Gene G29195T Point Mutation May Affect Diagnostic Reverse Transcription-PCR Detection. Microbiol. Spectr. 2022, 10, e0222321. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eur. Surveill. 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- WHO. Methods for the Detection and Identification of SARS-CoV-2 Variants. Available online: https://www.who.int/europe/publications/i/item/WHO-EURO-2021-2148-41903-57493 (accessed on 10 March 2021).
- Baker, D.J.; Aydin, A.; Le-Viet, T.; Kay, G.L.; Rudder, S.; de Oliveira Martins, L.; Tedim, A.P.; Kolyva, A.; Diaz, M.; Alikhan, N.F.; et al. CoronaHiT: High-throughput sequencing of SARS-CoV-2 genomes. Genome Med. 2021, 13, 21. [Google Scholar] [CrossRef]
- Rosato, A.E.; Msiha, E.; Weng, B.; Mesisca, M.; Gnass, R.; Gnass, S.; Bol, C.; Tabuenca, A.; Rosato, R.R. Rapid detection of the widely circulating B.1.617.2 (Delta) SARS-CoV-2 variant. Pathology 2022, 54, 351–356. [Google Scholar] [CrossRef]
- Lu, R.J.; Zhao, L.; Huang, B.Y.; Ye, F.; Wang, W.L.; Tan, W.J. Real-time reverse transcription-polymerase chain reaction assay panel for the detection of severe acute respiratory syndrome coronavirus 2 and its variants. Chin. Med. J. 2021, 134, 2048–2053. [Google Scholar] [CrossRef]
- Rodino, K.G.; Peaper, D.R.; Kelly, B.J.; Bushman, F.; Marques, A.; Adhikari, H.; Tu, Z.J.; Marrero Rolon, R.; Westblade, L.F.; Green, D.A.; et al. Partial ORF1ab Gene Target Failure with Omicron BA.2.12.1. J. Clin. Microbiol. 2022, 60, e0060022. [Google Scholar] [CrossRef]
- Tsui, W.N.T.; Hamill, V.; Noll, L.; Lu, N.; Porter, E.P.; Harbidge, D.; Cox, E.; Richardson, C.; Gray, M.; Sebhatu, T.; et al. Molecular detection of SARS-CoV-2 and differentiation of Omicron and Delta variant strains. Transbound. Emerg. Dis. 2022, 69, 2879–2889. [Google Scholar] [CrossRef]
- Bal, A.; Destras, G.; Gaymard, A.; Stefic, K.; Marlet, J.; Eymieux, S.; Regue, H.; Semanas, Q.; d’Aubarede, C.; Billaud, G.; et al. Two-step strategy for the identification of SARS-CoV-2 variant of concern 202012/01 and other variants with spike deletion H69-V70, France, August to December 2020. Eur. Surveill. 2021, 26, 2100008. [Google Scholar] [CrossRef]
- Matic, N.; Lowe, C.F.; Ritchie, G.; Stefanovic, A.; Lawson, T.; Jang, W.; Young, M.; Dong, W.; Brumme, Z.L.; Brumme, C.J.; et al. Rapid Detection of SARS-CoV-2 Variants of Concern, Including B.1.1.28/P.1, British Columbia, Canada. Emerg. Infect. Dis. 2021, 27, 1673–1676. [Google Scholar] [CrossRef]
- Yaniv, K.; Ozer, E.; Shagan, M.; Lakkakula, S.; Plotkin, N.; Bhandarkar, N.S.; Kushmaro, A. Direct RT-qPCR assay for SARS-CoV-2 variants of concern (Alpha, B.1.1.7 and Beta, B.1.351) detection and quantification in wastewater. Environ. Res. 2021, 201, 111653. [Google Scholar] [CrossRef] [PubMed]
- Lownik, J.C.; Farrar, J.S.; Way, G.W.; McKay, A.; Roychoudhury, P.; Greninger, A.L.; Martin, R.K. Fast SARS-CoV-2 Variant Detection Using Snapback Primer High-Resolution Melting. Diagnostics 2021, 11, 1788. [Google Scholar] [CrossRef]
- Yaniv, K.; Ozer, E.; Lewis, Y.; Kushmaro, A. RT-qPCR assays for SARS-CoV-2 variants of concern in wastewater reveals compromised vaccination-induced immunity. Water Res. 2021, 207, 117808. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.W.; Lidder, R.; Daigle, J.; Wonitowy, Q.; Dueck, C.; Nagasawa, A.; Mulvey, M.R.; Mangat, C.S. RT-qPCR detection of SARS-CoV-2 mutations S 69–70 del, S N501Y and N D3L associated with variants of concern in Canadian wastewater samples. Sci. Total Environ. 2022, 810, 151283. [Google Scholar] [CrossRef]
- Buchta, C.; Camp, J.V.; Jovanovic, J.; Radler, U.; Benka, B.; Puchhammer-Stöckl, E.; Müller, M.M.; Griesmacher, A.; Aberle, S.W.; Görzer, I. Inadequate design of mutation detection panels prevents interpretation of variants of concern: Results of an external quality assessment for SARS-CoV-2 variant detection. Clin. Chem. Lab. Med. 2022, 60, 291–298. [Google Scholar] [CrossRef]
- Bechtold, P.; Wagner, P.; Hosch, S.; Siegrist, D.; Ruiz-Serrano, A.; Gregorini, M.; Mpina, M.; Ondó, F.A.; Obama, J.; Ayekaba, M.O.; et al. Rapid Identification of SARS-CoV-2 Variants of Concern Using a Portable peakPCR Platform. Anal. Chem. 2021, 93, 16350–16359. [Google Scholar] [CrossRef]
- D’Agostino, Y.; Rocco, T.; Ferravante, C.; Porta, A.; Tosco, A.; Cappa, V.M.; Lamberti, J.; Alexandrova, E.; Memoli, D.; Terenzi, I.; et al. Rapid and sensitive detection of SARS-CoV-2 variants in nasopharyngeal swabs and wastewaters. Diagn. Microbiol. Infect. Dis. 2022, 102, 115632. [Google Scholar] [CrossRef]
- Xiong, D.; Zhang, X.; Shi, M.; Wang, N.; He, P.; Dong, Z.; Zhong, J.; Luo, J.; Wang, Y.; Yu, J.; et al. Developing an Amplification Refractory Mutation System-Quantitative Reverse Transcription-PCR Assay for Rapid and Sensitive Screening of SARS-CoV-2 Variants of Concern. Microbiol. Spectr. 2022, 10, e0143821. [Google Scholar] [CrossRef]
- Subramoney, K.; Mtileni, N.; Bharuthram, A.; Davis, A.; Kalenga, B.; Rikhotso, M.; Maphahlele, M.; Giandhari, J.; Naidoo, Y.; Pillay, S.; et al. Network for Genomics Surveillance in South, A. Identification of SARS-CoV-2 Omicron variant using spike gene target failure and genotyping assays, Gauteng, South Africa, 2021. J. Med. Virol. 2022, 94, 3676–3684. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.L.; Armas, F.; Guarneri, F.; Gu, X.; Formenti, N.; Wu, F.; Chandra, F.; Parisio, G.; Chen, H.; Xiao, A.; et al. Rapid displacement of SARS-CoV-2 variant Delta by Omicron revealed by allele-specific PCR in wastewater. Water Res. 2022, 221, 118809. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Deng, Y.; Ding, J.; Zheng, X.; Li, S.; Liu, L.; Chui, H.K.; Poon, L.L.M.; Zhang, T. Real-time allelic assays of SARS-CoV-2 variants to enhance sewage surveillance. Water Res. 2022, 220, 118686. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Zelyas, N.; Wong, A.; Croxen, M.A.; Lynch, T.; Buss, E.; Murphy, S.; Shokoples, S.; Kanji, J.; Tipples, G. Evolving strategy for an evolving virus: Development of real-time PCR assays for detecting all SARS-CoV-2 variants of concern. J. Virol. Methods 2022, 307, 114553. [Google Scholar] [CrossRef] [PubMed]
- Dikdan, R.J.; Marras, S.A.E.; Field, A.P.; Brownlee, A.; Cironi, A.; Hill, D.A.; Tyagi, S. Multiplex PCR Assays for Identifying all Major SARS-CoV-2 Variants. J. Mol. Diagn. JMD 2022, 24, 309–319. [Google Scholar] [CrossRef]
- Durand, M.; Thibault, P.; Lévesque, S.; Brault, A.; Carignan, A.; Valiquette, L.; Martin, P.; Labbé, S. Detection of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and its first variants in fourplex real-time quantitative reverse transcription-PCR assays. Microb. Cell 2022, 9, 1–20. [Google Scholar] [CrossRef]
- Hale, R.; Crowley, P.; Dervisevic, S.; Coupland, L.; Cliff, P.R.; Ebie, S.; Snell, L.B.; Paul, J.; Williams, C.; Randell, P.; et al. Development of a Multiplex Tandem PCR (MT-PCR) Assay for the Detection of Emerging SARS-CoV-2 Variants. Viruses 2021, 13, 2028. [Google Scholar] [CrossRef]
- Chung, H.Y.; Jian, M.J.; Chang, C.K.; Lin, J.C.; Yeh, K.M.; Chen, C.W.; Hsieh, S.S.; Hung, K.S.; Tang, S.H.; Perng, C.L.; et al. Emergency SARS-CoV-2 Variants of Concern: Novel Multiplex Real-Time RT-PCR Assay for Rapid Detection and Surveillance. Microbiol. Spectr. 2022, 10, e0251321. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.; Coppens, J.; Loens, K.; Le Mercier, M.; Xavier, B.B.; Lammens, C.; Vandamme, S.; Jansens, H.; Goossens, H.; Matheeussen, V. Monitoring the SARS-CoV-2 pandemic: Screening algorithm with single nucleotide polymorphism detection for the rapid identification of established and emerging variants. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2022, 28, 124–129. [Google Scholar] [CrossRef]
- Ong, D.S.Y.; Koeleman, J.G.M.; Vaessen, N.; Breijer, S.; Paltansing, S.; de Man, P. Rapid screening method for the detection of SARS-CoV-2 variants of concern. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2021, 141, 104903. [Google Scholar] [CrossRef] [PubMed]
- Banada, P.; Green, R.; Banik, S.; Chopoorian, A.; Streck, D.; Jones, R.; Chakravorty, S.; Alland, D. A Simple Reverse Transcriptase PCR Melting-Temperature Assay to Rapidly Screen for Widely Circulating SARS-CoV-2 Variants. J. Clin. Microbiol. 2021, 59, e0084521. [Google Scholar] [CrossRef]
- Yan, T.; Xu, Y.; Zheng, R.; Zeng, X.; Chen, Z.; Lin, S.; Xia, Z.; Liao, Y.; Zhang, Y.; Li, Q. Accessible and Adaptable Multiplexed Real-Time PCR Approaches to Identify SARS-CoV-2 Variants of Concern. Microbiol. Spectr. 2022, 10, e0322222. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.E.; Wang, Z.; Ostman, E.; Zheng, H.; Yao, H.; Cantarel, B.; Kanchwala, M.; Xing, C.; Chen, L.; Irwin, P.; et al. Multiplex Fragment Analysis for Flexible Detection of All SARS-CoV-2 Variants of Concern. Clin. Chem. 2022, 68, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Othman, S.; Lau, Y.L.; Radu, S.; Chee, H.Y. Loop-mediated isothermal amplification (LAMP): A versatile technique for detection of micro-organisms. J. Appl. Microbiol. 2018, 124, 626–643. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Ye, C.; Xiao, H.; Yin, J.; Liang, Y.; Ruan, Z.; Luo, D.; Gao, D.; Tan, Q.; Li, Y.; et al. Optimization of loop-mediated isothermal amplification (LAMP) assay for robust visualization in SARS-CoV-2 and emerging variants diagnosis. Chem. Eng. Sci. 2022, 251, 117430. [Google Scholar] [CrossRef]
- Alves, P.A.; de Oliveira, E.G.; Franco-Luiz, A.P.M.; Almeida, L.T.; Gonçalves, A.B.; Borges, I.A.; Rocha, F.S.; Rocha, R.P.; Bezerra, M.F.; Miranda, P.; et al. Optimization and Clinical Validation of Colorimetric Reverse Transcription Loop-Mediated Isothermal Amplification, a Fast, Highly Sensitive and Specific COVID-19 Molecular Diagnostic Tool That Is Robust to Detect SARS-CoV-2 Variants of Concern. Front. Microbiol. 2021, 12, 713713. [Google Scholar] [CrossRef]
- Jones, L.; Naikare, H.-K.; Mosley, Y.-Y.-C.; Tripp, R.-A. Isothermal amplification using sequence-specific fluorescence detection of SARS coronavirus 2 and variants in nasal swabs. Biotechniques 2022, 72, 263–272. [Google Scholar] [CrossRef]
- Lim, J.; Stavins, R.; Kindratenko, V.; Baek, J.; Wang, L.; White, K.; Kumar, J.; Valera, E.; King, W.P.; Bashir, R. Microfluidic point-of-care device for detection of early strains and B.1.1.7 variant of SARS-CoV-2 virus. Lab. Chip. 2022, 22, 1297–1309. [Google Scholar] [CrossRef]
- Talap, J.; Shen, M.; Yu, L.; Zeng, S.; Cai, S. RT-LAMP assay combining multi-fluorescent probes for SARS-CoV-2 RNA detection and variant differentiation. Talanta 2022, 248, 123644. [Google Scholar] [CrossRef]
- Yang, J.; Hu, X.; Wang, W.; Yang, Y.; Zhang, X.; Fang, W.; Zhang, L.; Li, S.; Gu, B. RT-LAMP assay for rapid detection of the R203M mutation in SARS-CoV-2 Delta variant. Emerg. Microbes Infect. 2022, 11, 978–987. [Google Scholar] [CrossRef]
- Chertow, D.S. Next-generation diagnostics with CRISPR. Science 2018, 360, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Andryukov, B.G. Six decades of lateral flow immunoassay: From determining metabolic markers to diagnosing COVID-19. AIMS Microbiol. 2020, 6, 280–304. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Gulati, S.; Ansari, A.H.; Phutela, R.; Acharya, S.; Azhar, M.; Murthy, J.; Kathpalia, P.; Kanakan, A.; Maurya, R.; et al. FnCas9-based CRISPR diagnostic for rapid and accurate detection of major SARS-CoV-2 variants on a paper strip. eLife 2021, 10, e67130. [Google Scholar] [CrossRef]
- Ali, Z.; Sánchez, E.; Tehseen, M.; Mahas, A.; Marsic, T.; Aman, R.; Sivakrishna Rao, G.; Alhamlan, F.S.; Alsanea, M.S.; Al-Qahtani, A.A.; et al. Bio-SCAN: A CRISPR/dCas9-Based Lateral Flow Assay for Rapid, Specific, and Sensitive Detection of SARS-CoV-2. ACS Synth. Biol. 2022, 11, 406–419. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, F.; Zhu, K.; Lin, W.; Ma, W. dsmCRISPR: Dual synthetic mismatches CRISPR/Cas12a-based detection of SARS-CoV-2 D614G mutation. Virus Res. 2021, 304, 198530. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Lin, H.; Zou, L.; Zhao, J.; Li, B.; Wang, H.; Lu, J.; Sun, J.; Yang, X.; Deng, X.; et al. CRISPR-Cas12a-Based Detection for the Major SARS-CoV-2 Variants of Concern. Microbiol. Spectr. 2021, 9, e0101721. [Google Scholar] [CrossRef]
- Ning, B.; Youngquist, B.M.; Li, D.D.; Lyon, C.J.; Zelazny, A.; Maness, N.J.; Tian, D.; Hu, T.Y. Rapid detection of multiple SARS-CoV-2 variants of concern by PAM-targeting mutations. Cell Rep. Methods 2022, 2, 100173. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Lin, C.; Mo, G.; Xi, B.; Li, A.A.; Huang, D.; Wan, Y.; Chen, F.; Liang, Y.; Zuo, Q.; et al. Rapid and accurate detection of SARS-CoV-2 mutations using a Cas12a-based sensing platform. Biosens. Bioelectron. 2022, 198, 113857. [Google Scholar] [CrossRef]
- Yang, J.; Barua, N.; Rahman, M.N.; Lo, N.; Tsang, T.F.; Yang, X.; Chan, P.K.S.; Zhang, L.; Ip, M. Chimeric crRNA improves CRISPR-Cas12a specificity in the N501Y mutation detection of Alpha, Beta, Gamma, and Mu variants of SARS-CoV-2. PLoS ONE 2021, 16, e0261778. [Google Scholar] [CrossRef]
- Liang, Y.; Zou, L.; Lin, H.; Li, B.; Zhao, J.; Wang, H.; Sun, J.; Chen, J.; Mo, Y.; Yang, X.; et al. Detection of Major SARS-CoV-2 Variants of Concern in Clinical Samples via CRISPR-Cas12a-Mediated Mutation-Specific Assay. ACS Synth. Biol. 2022, 11, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- De Puig, H.; Lee, R.A.; Najjar, D.; Tan, X.; Soeknsen, L.R.; Angenent-Mari, N.M.; Donghia, N.M.; Weckman, N.E.; Ory, A.; Ng, C.F.; et al. Minimally instrumented SHERLOCK (miSHERLOCK) for CRISPR-based point-of-care diagnosis of SARS-CoV-2 and emerging variants. Sci. Adv. 2021, 7, eabh2944. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Macaluso, N.C.; Pizzano, B.L.M.; Cash, M.N.; Spacek, J.; Karasek, J.; Miller, M.R.; Lednicky, J.A.; Dinglasan, R.R.; Salemi, M.; et al. A thermostable Cas12b from Brevibacillus leverages one-pot discrimination of SARS-CoV-2 variants of concern. eBioMedicine 2022, 77, 103926. [Google Scholar] [CrossRef] [PubMed]
- Arizti-Sanz, J.; Bradley, A.D.; Zhang, Y.B.; Boehm, C.K.; Freije, C.A.; Grunberg, M.E.; Kosoko-Thoroddsen, T.F.; Welch, N.L.; Pillai, P.P.; Mantena, S.; et al. Equipment-free detection of SARS-CoV-2 and Variants of Concern using Cas13. medRxiv Prepr. Serv. Health Sci. 2021. [Google Scholar] [CrossRef]
- Lin, H.; Liang, Y.; Zou, L.; Li, B.; Zhao, J.; Wang, H.; Sun, J.; Deng, X.; Tang, S. Combination of Isothermal Recombinase-Aided Amplification and CRISPR-Cas12a-Mediated Assay for Rapid Detection of Major Severe Acute Respiratory Syndrome Coronavirus 2 Variants of Concern. Front. Microbiol. 2022, 13, 945133. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Han, Y.; Dong, X.; Yang, L.; Li, F.; Zhang, Y.; Hu, Q.; Xia, X.; Li, H.; Sun, Y. Highly Sensitive Detection Method for HV69-70del in SARS-CoV-2 Alpha and Omicron Variants Based on CRISPR/Cas13a. Front. Bioeng. Biotechnol. 2022, 10, 831332. [Google Scholar] [CrossRef]
- Pekin, D.; Skhiri, Y.; Baret, J.C.; Le Corre, D.; Mazutis, L.; Salem, C.B.; Millot, F.; El Harrak, A.; Hutchison, J.B.; Larson, J.W.; et al. Quantitative and sensitive detection of rare mutations using droplet-based microfluidics. Lab. Chip. 2011, 11, 2156–2166. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, L.; Elsinga, G.; de Graaf, M.; Molenkamp, R.; Koopmans, M.P.G.; Medema, G. Droplet digital RT-PCR to detect SARS-CoV-2 signature mutations of variants of concern in wastewater. Sci. Total Environ. 2021, 799, 149456. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.; Hughes, B.; Duong, D.; Chan-Herur, V.; Wigginton, K.R.; White, B.J.; Boehm, A.B. Detection of SARS-CoV-2 Variants Mu, Beta, Gamma, Lambda, Delta, Alpha, and Omicron in Wastewater Settled Solids Using Mutation-Specific Assays Is Associated with Regional Detection of Variants in Clinical Samples. Appl. Environ. Microbiol. 2022, 88, e0004522. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.G.; Hajian, P.; Bakhash, S.M.; Xie, H.; Mantzke, D.; Zhu, H.; Perchetti, G.A.; Huang, M.L.; Pepper, G.; Jerome, K.R.; et al. Rapid and accurate identification of SARS-CoV-2 Omicron variants using droplet digital PCR (RT-ddPCR). J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2022, 154, 105218. [Google Scholar] [CrossRef] [PubMed]
- Lou, E.G.; Sapoval, N.; McCall, C.; Bauhs, L.; Carlson-Stadler, R.; Kalvapalle, P.; Lai, Y.; Palmer, K.; Penn, R.; Rich, W.; et al. Direct comparison of RT-ddPCR and targeted amplicon sequencing for SARS-CoV-2 mutation monitoring in wastewater. Sci. Total Environ. 2022, 833, 155059. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, H.; Ichinohe, R.; Koshikawa, T. High-resolution melting analysis after nested PCR for the detection of SARS-CoV-2 spike protein G339D and D796Y variations. Biochem. Biophys. Res. Commun. 2022, 606, 128–134. [Google Scholar] [CrossRef]
- Cherkaoui, D.; Huang, D.; Miller, B.S.; Turbe, V.; McKendry, R.A. Harnessing recombinase polymerase amplification for rapid multi-gene detection of SARS-CoV-2 in resource-limited settings. Biosens. Bioelectron. 2021, 189, 113328. [Google Scholar] [CrossRef] [PubMed]
- Cherkaoui, D.; Heaney, J.; Huang, D.; Byott, M.; Miller, B.S.; Nastouli, E.; McKendry, R.A. Clinical Validation of a Rapid Variant-Proof RT-RPA Assay for the Detection of SARS-CoV-2. Diagnostics 2022, 12, 1263. [Google Scholar] [CrossRef]
- Yang, K.; Schuder, D.N.; Ngor, A.K.; Chaput, J.C. REVEALR-Based Genotyping of SARS-CoV-2 Variants of Concern in Clinical Samples. J. Am. Chem. Soc. 2022, 144, 11685–11692. [Google Scholar] [CrossRef]
- Yoon, T.; Shin, J.; Choi, H.J.; Park, K.S. Split T7 promoter-based isothermal transcription amplification for one-step fluorescence detection of SARS-CoV-2 and emerging variants. Biosens. Bioelectron. 2022, 208, 114221. [Google Scholar] [CrossRef]
- Dorlass, E.G.; Lourenço, K.L.; Magalhães, R.D.M.; Sato, H.; Fiorini, A.; Peixoto, R.; Coelho, H.P.; Telezynski, B.L.; Scagion, G.P.; Ometto, T.; et al. Survey of SARS-CoV-2 genetic diversity in two major Brazilian cities using a fast and affordable Sanger sequencing strategy. Genomics 2021, 113, 4109–4115. [Google Scholar] [CrossRef]
- Bloemen, M.; Rector, A.; Swinnen, J.; Ranst, M.V.; Maes, P.; Vanmechelen, B.; Wollants, E. Fast Detection of SARS-CoV-2 Variants including Omicron Using One-step RT-PCR and Sanger Sequencing. J. Virol. Methods 2022, 304, 114512. [Google Scholar] [CrossRef]
- Lee, S.H. A Routine Sanger Sequencing Target Specific Mutation Assay for SARS-CoV-2 Variants of Concern and Interest. Viruses 2021, 13, 2386. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, M.; Imaizumi, Y.; Fujikawa, T.; Ishige, T.; Nishimura, M.; Miyabe, A.; Murata, S.; Kawasaki, K.; Taniguchi, T.; Igari, H.; et al. Tracking SARS-CoV-2 variants by entire S-gene analysis using long-range RT-PCR and Sanger sequencing. Clin. Chim. Acta 2022, 530, 94–98. [Google Scholar] [CrossRef]
- Van Poelvoorde, L.A.E.; Delcourt, T.; Coucke, W.; Herman, P.; De Keersmaecker, S.C.J.; Saelens, X.; Roosens, N.H.C.; Vanneste, K. Strategy and Performance Evaluation of Low-Frequency Variant Calling for SARS-CoV-2 Using Targeted Deep Illumina Sequencing. Front. Microbiol. 2021, 12, 747458. [Google Scholar] [CrossRef]
- Castañeda-Mogollón, D.; Kamaliddin, C.; Fine, L.; Oberding, L.K.; Pillai, D.R. SARS-CoV-2 variant detection with ADSSpike. Diagn. Microbiol. Infect. Dis. 2022, 102, 115606. [Google Scholar] [CrossRef]
- Dachert, C.; Muenchhoff, M.; Graf, A.; Autenrieth, H.; Bender, S.; Mairhofer, H.; Wratil, P.R.; Thieme, S.; Krebs, S.; Grzimek-Koschewa, N.; et al. Rapid and sensitive identification of omicron by variant-specific PCR and nanopore sequencing: Paradigm for diagnostics of emerging SARS-CoV-2 variants. Med. Microbiol. Immunol. 2022, 211, 71–77. [Google Scholar] [CrossRef]
- Bull, R.A.; Adikari, T.N.; Ferguson, J.M.; Hammond, J.M.; Stevanovski, I.; Beukers, A.G.; Naing, Z.; Yeang, M.; Verich, A.; Gamaarachchi, H.; et al. Analytical validity of nanopore sequencing for rapid SARS-CoV-2 genome analysis. Nat. Commun. 2020, 11, 6272. [Google Scholar] [CrossRef]
- Abnizova, I.; Boekhorst, R.t.; Orlov, Y.L. Computational Errors and Biases in Short Read Next Generation Sequencing. J. Proteom. Bioinform. 2017, 10. [Google Scholar] [CrossRef]
- Chappleboim, A.; Joseph-Strauss, D.; Rahat, A.; Sharkia, I.; Adam, M.; Kitsberg, D.; Fialkoff, G.; Lotem, M.; Gershon, O.; Schmidtner, A.K.; et al. Early sample tagging and pooling enables simultaneous SARS-CoV-2 detection and variant sequencing. Sci. Transl. Med. 2021, 13, eabj2266. [Google Scholar] [CrossRef]
- Chiari, W.; Damayanti, R.; Harapan, H.; Puspita, K.; Saiful, S.; Rahmi, R.; Rizki, D.R.; Iqhrammullah, M. Trend of Polymer Research Related to COVID-19 Pandemic: Bibliometric Analysis. Polymers 2022, 14, 3297. [Google Scholar] [CrossRef]
- Di Domenico, M.; De Rosa, A.; Boccellino, M. Detection of SARS-COV-2 Proteins Using an ELISA Test. Diagnostics 2021, 11, 698. [Google Scholar] [CrossRef]
- Hingrat, Q.L.; Visseaux, B.; Laouenan, C.; Tubiana, S.; Bouadma, L.; Yazdanpanah, Y.; Duval, X.; Burdet, C.; Ichou, H.; Damond, F.; et al. Detection of SARS-CoV-2 N-antigen in blood during acute COVID-19 provides a sensitive new marker and new testing alternatives. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 27, 789.e1–789.e5. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, M.; Jung, Y.; Lee, S.K.; Lee, C.S.; Kim, J.; Kim, J.; Kim, N.H.; Kim, B.T.; Kim, H.G. A novel rapid detection for SARS-CoV-2 spike 1 antigens using human angiotensin converting enzyme 2 (ACE2). Biosens. Bioelectron. 2021, 171, 112715. [Google Scholar] [CrossRef]
- Dinnes, J.; Deeks, J.J.; Berhane, S.; Taylor, M.; Adriano, A.; Davenport, C.; Dittrich, S.; Emperador, D.; Takwoingi, Y.; Cunningham, J.; et al. Rapid, point-of-care antigen and molecular-based tests for diagnosis of SARS-CoV-2 infection. Cochrane Database Syst. Rev. 2021, 3, CD013705. [Google Scholar] [PubMed]
- Igloi, Z.; Velzing, J.; van Beek, J.; van de Vijver, D.; Aron, G.; Ensing, R.; Benschop, K.; Han, W.; Boelsums, T.; Koopmans, M.; et al. Clinical Evaluation of Roche SD Biosensor Rapid Antigen Test for SARS-CoV-2 in Municipal Health Service Testing Site, the Netherlands. Emerg. Infect. Dis. 2021, 27, 1323–1329. [Google Scholar] [CrossRef]
- Hsiao, N.-y.; Ngo Nsoga, M.T.; Kronig, I.; Perez Rodriguez, F.J.; Sattonnet-Roche, P.; Da Silva, D.; Helbling, J.; Sacks, J.A.; de Vos, M.; Boehm, E.; et al. Diagnostic accuracy of Panbio rapid antigen tests on oropharyngeal swabs for detection of SARS-CoV-2. PLoS ONE 2021, 16, e0253321. [Google Scholar]
- Medoro, A.; Davinelli, S.; Voccola, S.; Cardinale, G.; Passarella, D.; Marziliano, N.; Intrieri, M. Assessment of the Diagnostic Performance of a Novel SARS-CoV-2 Antigen Sealing Tube Test Strip (Colloidal Gold) as Point-of-Care Surveillance Test. Diagnostics 2022, 12, 1279. [Google Scholar] [CrossRef]
- Peronace, C.; Tallerico, R.; Colosimo, M.; Sacco, V.; Talarico, R.; De Fazio, M.; Pasceri, F.; Talotta, I.; Panduri, G.; Kim, J.H.; et al. Validation of GeneFinder COVID-19 Ag Plus Rapid Test and Its Potential Utility to Slowing Infection Waves: A Single-Center Laboratory Evaluation Study. Diagnostics 2022, 12, 1126. [Google Scholar] [CrossRef]
- Jungnick, S.; Hobmaier, B.; Mautner, L.; Hoyos, M.; Haase, M.; Baiker, A.; Lahne, H.; Eberle, U.; Wimmer, C.; Hepner, S.; et al. Detection of the new SARS-CoV-2 variants of concern B.1.1.7 and B.1.351 in five SARS-CoV-2 rapid antigen tests (RATs), Germany, March 2021. Eur. Surveill. 2021, 26, 2100413. [Google Scholar] [CrossRef] [PubMed]
- Osterman, A.; Iglhaut, M.; Lehner, A.; Späth, P.; Stern, M.; Autenrieth, H.; Muenchhoff, M.; Graf, A.; Krebs, S.; Blum, H.; et al. Comparison of four commercial, automated antigen tests to detect SARS-CoV-2 variants of concern. Med. Microbiol. Immunol. 2021, 210, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Jungnick, S.; Hobmaier, B.; Mautner, L.; Hoyos, M.; Haase, M.; Baiker, A.; Lahne, H.; Eberle, U.; Wimmer, C.; Hepner, S.; et al. In Vitro Rapid Antigen Test Performance with the SARS-CoV-2 Variants of Concern B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma), and B.1.617.2 (Delta). Microorganisms 2021, 9, 1967. [Google Scholar] [CrossRef]
- Barrera-Avalos, C.; Luraschi, R.; Vallejos-Vidal, E.; Mella-Torres, A.; Hernández, F.; Figueroa, M.; Rioseco, C.; Valdés, D.; Imarai, M.; Acuña-Castillo, C.; et al. The Rapid Antigen Detection Test for SARS-CoV-2 Underestimates the Identification of COVID-19 Positive Cases and Compromises the Diagnosis of the SARS-CoV-2 (K417N/T, E484K, and N501Y) Variants. Front. Public Health 2021, 9, 780801. [Google Scholar] [CrossRef]
- Szekely, J.; Mongkolprasert, J.; Jeayodae, N.; Senorit, C.; Chaimuti, P.; Swangphon, P.; Nanakorn, N.; Nualnoi, T.; Wongwitwichot, P.; Pengsakul, T. Development, Analytical, and Clinical Evaluation of Rapid Immunochromatographic Antigen Test for SARS-CoV-2 Variants Detection. Diagnostics 2022, 12, 381. [Google Scholar] [CrossRef]
- Weishampel, Z.A.; Young, J.; Fischl, M.; Fischer, R.J.; Donkor, I.O.; Riopelle, J.C.; Schulz, J.E.; Port, J.R.; Saturday, T.A.; van Doremalen, N.; et al. OraSure InteliSwab Rapid Antigen Test Performance with the SARS-CoV-2 Variants of Concern-Alpha, Beta, Gamma, Delta, and Omicron. Viruses 2022, 14, 543. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Bassit, L.; Lin, J.; Verma, K.; Bowers, H.B.; Pachura, K.; Greenleaf, M.; Sullivan, J.; Lai, E.; Creager, R.S.; et al. Assessment of the Abbott BinaxNOW SARS-CoV-2 rapid antigen test against viral variants of concern. iScience 2022, 25, 103968. [Google Scholar] [CrossRef]
- Liu, L.; Meyers, K.; Purpura, L.J.; Nguyen, N.; Mohri, H.; Chang, J.Y.; Annavajhala, M.K.; Lopez, L., 3rd; Lee, S.W.; Shah, J.; et al. Development and performance of a point-of-care rapid antigen test for detection of SARS-COV-2 variants. J. Clin. Virol. Plus 2022, 2, 100080. [Google Scholar] [CrossRef] [PubMed]
- Soni, A.; Herbert, C.; Filippaios, A.; Broach, J.; Colubri, A.; Fahey, N.; Woods, K.; Nanavati, J.; Wright, C.; Orwig, T.; et al. Comparison of Rapid Antigen Tests’ Performance between Delta (B.1.61.7; AY.X) and Omicron (B.1.1.529; BA1) Variants of SARS-CoV-2: Secondary Analysis from a Serial Home Self-Testing Study. medRxiv Prepr. Serv. Health Sci. 2022. [Google Scholar] [CrossRef]
- Zhan, Z.; Li, J.; Cheng, Z.J. Rapid Antigen Test Combine with Nucleic Acid Detection: A Better Strategy for COVID-19 Screening at Points of Entry. J. Epidemiol. Glob. Health 2022, 12, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Iwasaki, S.; Oguri, S.; Tanaka, K.; Suzuki, R.; Hayasaka, K.; Fujisawa, S.; Watanabe, C.; Konno, S.; Yokota, I.; et al. SARS-CoV-2 Omicron detection by antigen tests using saliva. J. Clin. Virol. Plus 2022, 2, 100109. [Google Scholar] [CrossRef]
- Puligedda, R.D.; Al-Saleem, F.H.; Wirblich, C.; Kattala, C.D.; Jović, M.; Geiszler, L.; Devabhaktuni, H.; Feuerstein, G.Z.; Schnell, M.J.; Sack, M.; et al. A Strategy to Detect Emerging Non-Delta SARS-CoV-2 Variants with a Monoclonal Antibody Specific for the N501 Spike Residue. Diagnostics 2021, 11, 2092. [Google Scholar] [CrossRef]
- Adashek, J.J.; Kurzrock, R. Balancing clinical evidence in the context of a pandemic. Nat. Biotechnol. 2021, 39, 270–274. [Google Scholar] [CrossRef]
- Lee, J.H.; Jung, Y.; Lee, S.K.; Kim, J.; Lee, C.S.; Kim, S.; Lee, J.S.; Kim, N.H.; Kim, H.G. Rapid Biosensor of SARS-CoV-2 Using Specific Monoclonal Antibodies Recognizing Conserved Nucleocapsid Protein Epitopes. Viruses 2022, 14, 255. [Google Scholar] [CrossRef]
- Barlev-Gross, M.; Weiss, S.; Paran, N.; Yahalom-Ronen, Y.; Israeli, O.; Nemet, I.; Kliker, L.; Zuckerman, N.; Glinert, I.; Noy-Porat, T.; et al. Sensitive Immunodetection of Severe Acute Respiratory Syndrome Coronavirus 2 Variants of Concern 501Y.V2 and 501Y.V1. J. Infect. Dis. 2021, 224, 616–619. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e9. [Google Scholar] [CrossRef]
- Ozono, S.; Zhang, Y.; Ode, H.; Sano, K.; Tan, T.S.; Imai, K.; Miyoshi, K.; Kishigami, S.; Ueno, T.; Iwatani, Y.; et al. SARS-CoV-2 D614G spike mutation increases entry efficiency with enhanced ACE2-binding affinity. Nat. Commun. 2021, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Nie, J.; Wu, J.; Zhang, L.; Ding, R.; Wang, H.; Zhang, Y.; Li, T.; Liu, S.; Zhang, M.; et al. SARS-CoV-2 501Y.V2 variants lack higher infectivity but do have immune escape. Cell 2021, 184, 2362–2371.e9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, Y.; Lee, S.K.; Kim, J.; Lee, C.S.; Kim, N.H.; Kim, H.G. Versatile role of ACE2-based biosensors for detection of SARS-CoV-2 variants and neutralizing antibodies. Biosens. Bioelectron. 2022, 203, 114034. [Google Scholar] [CrossRef]
- Zhao, F.; Lu, J.; Lu, B.; Qin, T.; Wang, X.; Hou, X.; Meng, F.; Xu, X.; Li, T.; Zhou, H.; et al. A Novel Strategy for the Detection of SARS-CoV-2 Variants Based on Multiplex PCR-Mass Spectrometry Minisequencing Technology. Microbiol. Spectr. 2021, 9, e0126721. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.M.; Banu, R.; Gonzalez-Reiche, A.S.; van de Guchte, A.; Khan, Z.; Shrestha, P.; Cao, L.; Chen, F.; Shi, H.; Hanna, A.; et al. Robust clinical detection of SARS-CoV-2 variants by RT-PCR/MALDI-TOF multitarget approach. J. Med. Virol. 2022, 94, 1606–1616. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Wang, J.; He, S.; Su, X.; Huang, W.; Chen, M.; Zhuo, Z.; Zhu, X.; Fang, M.; Li, T.; et al. An encodable multiplex microsphere-phase amplification sensing platform detects SARS-CoV-2 mutations. Biosens. Bioelectron. 2022, 203, 114032. [Google Scholar] [CrossRef]
- Welch, N.L.; Zhu, M.; Hua, C.; Weller, J.; Mirhashemi, M.E.; Nguyen, T.G.; Mantena, S.; Bauer, M.R.; Shaw, B.M.; Ackerman, C.M.; et al. Multiplexed CRISPR-based microfluidic platform for clinical testing of respiratory viruses and identification of SARS-CoV-2 variants. Nat. Med. 2022, 28, 1083–1094. [Google Scholar] [CrossRef]


{kind=link}
| SARS-CoV-2 Variants | Mutation-Specific Targets | References |
|---|---|---|
| Alpha | ΔH69/ΔV70 + N501Y | [14] |
| Alpha; Beta; Gamma | N501Y + delHV69/70; N501Y + K417N; N501Y + V1176F | [15] |
| Alpha; Beta | S.1.1.7; B.241–243 | [16] |
| Alpha; Beta; Gamma | L452R + D614G; L5F + L18F + Δ69–70 + D80 + Δ144 + E484K + N501Y + D614G + P681H; D80 + Δ241-3 + K417N + N501Y + D614G; L18F + T20N + P26S + D138Y + K417T + E484K + N501Y + D614G + V1176F | [17] |
| Gamma; Delta | four-nucleotide insertion of ORF8; SΔ157–158 | [18] |
| Alpha; Beta; Gamma | S69–70del + SN501Y + ND3L; SN501Y | [19] |
| Alpha; Beta; Gamma; Delta | N501Y + HV69/70del + E484; N501Y + E484K + K417N; N501Y + E484K + K417T + V1176F; N501 + E484 + (L452R or P681R or T478K) | [20] |
| Alpha; Beta; Gamma | HV69/70 + N501Y; N501Y + E484K; | [21] |
| Alpha; Beta; Gamma; Delta | P314L + RG203_204KR + N501Y; P314L + N501Y + E484K + K417N + A701V; P314L + RG203_204KR + N501Y + E484K + V1176F; P314L + L452R | [22] |
| Alpha; Delta | C1709A; C56G | [23] |
| Omicron BA.1 | SGTF +ΔH69/ΔV70 + K417N | [24] |
| Omicron BA.1; Omicron BA.2 | Q493R + Q498R + G496S; Q493R + Q498R | [25] |
| Alpha; Beta; Gamma; Delta; Omicron | N501Y +ΔH69/ΔV70; N501Y + E484K + K417N; N501Y + E484K + K417T; L452R + P681R + T478K; N501Y +ΔH69/ΔV70 + T478K | [26] |
| Alpha; Beta + Gamma; Delta; Omicron | N501Y + ΔH69/V70 + E gene; 242–244 WT + K417N + E484K + K417T; L452R + P681 WT + E484Q; N501Y + ΔH69/V70 + E gene + 242-244 WT + K417N | [27] |
| Samples | Test Kits | SARS-CoV-2 Variants | Detection Performance | References |
|---|---|---|---|---|
| Infectious Vero E6 cell culture supernatants | SARS-CoV-2 Rapid Antigen Test (Roche) | Alpha | LOD in RNA copies/mL: 8.9 × 105 (DMEM) and 1.9 × 106 (saliva) | [88] |
| Beta | LOD in RNA copies/mL: 8.5 × 105 (DMEM) and 1.1 × 106 (saliva) | |||
| CLINITEST Rapid COVID-19 Antigen Test (Siemens) | Alpha | LOD in RNA copies/mL: 8.9 × 105 (DMEM) and 1.9 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 8.5 × 105 (DMEM) and 1.1 × 106 (saliva) | |||
| Panbio COVID-19 Ag RAPID TEST DEVICE (Abbott) | Alpha | LOD in RNA copies/mL: 8.9 × 105 (DMEM) and 1.9 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 8.5 × 105 (DMEM) and 1.1 × 106 (saliva) | |||
| NADAL COVID-19 Ag rapid test (nal von minden) | Alpha | LOD in RNA copies/mL: 8.9 × 105 (DMEM) and 1.9 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 8.5 × 105 (DMEM) and 6.4 × 106 (saliva) | |||
| BIOCREDIT COVID-19 Ag rapid test kit (RapiGEN) | Alpha | LOD in RNA copies/mL: 3.4 × 107 (DMEM) and 6.6 × 107 (saliva) | ||
| Beta | LOD in RNA copies/mL: 2.6 × 107 (DMEM) and 3.9 × 107 (saliva) | |||
| A total of 410 respiratory samples | Lumipulse G SARS-CoV-2 Ag (CLEIA) | Alpha | Viral loads: 1 × 105 Geq/mL | [89] |
| Beta | Viral loads: 1 × 105 Geq/mL | |||
| Elecsys SARS-CoV-2 Ag (ECLIA) | Alpha | Viral loads: 5 × 105 Geq/mL | ||
| Beta | Viral loads: 2 × 106 Geq/mL | |||
| LIAISON SARS-CoV-2 Ag (CLIA) | Alpha | Viral loads: 8 × 106 Geq/mL | ||
| Beta | Viral loads: 8 × 106 Geq/mL | |||
| SARS-CoV-2 Ag ELISA (ELISA) | Alpha | Viral loads: 8 × 106 Geq/mL | ||
| Beta | Viral loads: 8 × 106 Geq/mL | |||
| Cell culture medium and pooled saliva | SARS-CoV-2 Rapid Antigen Test | Alpha | LOD in RNA copies/mL: 1 × 106 (DMEM) and 1 × 106 (saliva) | [90] |
| Beta | LOD in RNA copies/mL: 2 × 106 (DMEM) and 2 × 106 (saliva) | |||
| Gamma | LOD in RNA copies/mL: 1 × 106 (DMEM) and 1 × 106 (saliva) | |||
| Delta | LOD in RNA copies/mL: 2 × 106 (DMEM) and 2 × 106 (saliva) | |||
| CLINITEST Rapid COVID-19 Antigen Self-Test | Alpha | LOD in RNA copies/mL: 1 × 106 (DMEM) and 2 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 1 × 105 (DMEM) and 1 × 105 (saliva) | |||
| Gamma | LOD in RNA copies/mL: 1 × 106 (DMEM) and 1 × 106 (saliva) | |||
| Delta | LOD in RNA copies/mL: 1 × 106 (DMEM) and 1 × 106 (saliva) | |||
| Rapid SARS-CoV-2 Antigen Test Card | Alpha | LOD in RNA copies/mL: 1 × 105 (DMEM) and 1 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 1 × 105 (DMEM) and 2 × 106 (saliva) | |||
| Gamma | LOD in RNA copies/mL: 2 × 106 (DMEM) and 2 × 106 (saliva) | |||
| Delta | LOD in RNA copies/mL: 1 × 106 (DMEM) and 1 × 106 (saliva) | |||
| Panbio COVID-19 Ag RAPID TEST DEVICE | Alpha | LOD in RNA copies/mL: 2 × 106 (DMEM) and 2 × 106 (saliva) | ||
| Beta | LOD in RNA copies/mL: 2 × 106 (DMEM) and 1 × 105 (saliva) | |||
| Gamma | LOD in RNA copies/mL: 1×106 (DMEM) and 2×106 (saliva) | |||
| Delta | LOD in RNA copies/mL: 2 × 106 (DMEM) and 2 × 106 (saliva) | |||
| 55 nasopharyngeal swab samples | SD BIOSENSOR | Beta, Gamma | Sensitivity (a positive group diagnosed with the SARS-CoV-2 variants by RT-qPCR): 42.8% with RT-qPCR amplification range 20 ≤ Cq < 25 | [91] |
| 319 nasopharyngeal specimens | the KestrelTM COVID-19 Ag Rapid Test | Alpha | LOD: 0.156 ng/mL | [92] |
| Beta | LOD: 0.156 ng/mL | |||
| Gamma | LOD: 0.156 ng/mL | |||
| Delta | LOD: 0.156 ng/mL | |||
| Epsilon | LOD: 0.156 ng/mL | |||
| Kappa | LOD: 0.156 ng/mL | |||
| Omicron | LOD: 0.39 ng/mL | |||
| VeroE6 cells and four six-week-old Syrian golden hamsters | OraSure InteliSwab™ Rapid Antigen Test | Alpha | LOD: 0.313 ng/mL, genome Copies/mL: 6.06 × 105 | [93] |
| Beta | LOD: 0.469 ng/mL, genome Copies/mL: 3.77 × 105 | |||
| Gamma | LOD: 0.313 ng/mL, genome Copies/mL: 4.30 × 105 | |||
| Delta | LOD: 0.469 ng/mL, genome Copies/mL: 9.13 × 105 | |||
| Omicron | LOD: 0.469 ng/mL, genome Copies/mL: 4.51 × 105 | |||
| SARS-CoV-2 heat-inactivated positive samples | The Abbott BinaxNOW SARS-CoV-2 rapid antigen test | Alpha | Lowest viral load (highest RT-qPCR Ct value): 28.9 | [94] |
| Beta | Lowest viral load (highest RT-qPCR Ct value): 25.92 | |||
| Gamma | Lowest viral load (highest RT-qPCR Ct value): 26.14 | |||
| Delta | Lowest viral load (highest RT-qPCR Ct value): 26.7 | |||
| Eta | Lowest viral load (highest RT-qPCR Ct value): 26.28 | |||
| Lambda | Lowest viral load (highest RT-qPCR Ct value): 23.81 | |||
| Mu | Lowest viral load (highest RT-qPCR Ct value): 24.25 | |||
| Omicron | Lowest viral load (highest RT-qPCR Ct value): 24.6 | |||
| 10 authentic SARS-CoV-2 variants, and 148 symptomatic clinical samples | CoV-SCAN | Alpha | LOD: 6.25 TCID50/swab | [95] |
| Beta | LOD: 12.5 TCID50/swab | |||
| Gamma | LOD: 12.5 TCID50/swab | |||
| Delta | LOD: 6.25 TCID50/swab | |||
| Omicron | LOD: 3.2 TCID50/swab; Sensitivity (a positive group diagnosed with the SARS-CoV-2 variants by RT-qPCR): 93.8% |
| Methods | Advantages | Disadvantages |
|---|---|---|
| Mutation-specific SARS-CoV-2 PCR/Multiplex PCR | Short turnaround times; 98.6% sensitivity; reduce the number of reagents required for sample analysis and hands-on time; simultaneous detection of multiple mutation sites | The genetic information provided is limited; the mutant nature of the SARS-CoV-2 variant needs to be known |
| LAMP | Sensitivity of 90% or more; simple operation; fast amplification speed; high efficiency; no need for complex thermal cycler; can be used at the grass-roots level, laboratories with poor experimental conditions, and POC | False-positive; cross-contamination; poor stability |
| CRISPR-Cas-based detection technology | 100.0% specificity and accuracy; rapid screening; low cost; suitable for POC diagnosis | Only for point mutations; relatively low sensitivity to samples with low viral load 53.97% |
| RT-ddPCR | Absolutely quantitative; sensitive and accurate | Expensive |
| RAT | Short turnaround times; easy to use; cost-effective; no complex instrumentation and/or expertise required for results interpretation; suitable for home inspection analysis | False-negative; sensitivity is not high compared to nucleic acid detection-based; particularly sensitive to sample quality |
| ELISA | High negative predictive value; high throughput suitable for POC detection | Low sensitivity due to large differences in affinity between variants; no clinically implemented testing |
| LFA | Specificity; long-term stability; suitable for home testing and self-testing | Low sensitivity due to the identification of only specific regions of the antigen |
| Viral genome sequencing | High-throughput sequencing; super sensitive; provide a detailed map of new mutations; accurately identify mutation types | Resource-intensive; long turnaround times of hours to weeks; expensive; demanding in technic and equipment |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, W.; Ji, W.; Zhang, Y.; Xie, Y.; Chen, S.; Jin, Y.; Duan, G. An Update on Detection Technologies for SARS-CoV-2 Variants of Concern. Viruses 2022, 14, 2324. https://doi.org/10.3390/v14112324
Jiang W, Ji W, Zhang Y, Xie Y, Chen S, Jin Y, Duan G. An Update on Detection Technologies for SARS-CoV-2 Variants of Concern. Viruses. 2022; 14(11):2324. https://doi.org/10.3390/v14112324
Chicago/Turabian StyleJiang, Wenjie, Wangquan Ji, Yu Zhang, Yaqi Xie, Shuaiyin Chen, Yuefei Jin, and Guangcai Duan. 2022. "An Update on Detection Technologies for SARS-CoV-2 Variants of Concern" Viruses 14, no. 11: 2324. https://doi.org/10.3390/v14112324
APA StyleJiang, W., Ji, W., Zhang, Y., Xie, Y., Chen, S., Jin, Y., & Duan, G. (2022). An Update on Detection Technologies for SARS-CoV-2 Variants of Concern. Viruses, 14(11), 2324. https://doi.org/10.3390/v14112324