
The sORF-Encoded Peptides, ATP Synthase Subunits, Facilitate WSSV Duplication in Shrimp

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis
2.2. Tissue Distribution and Expression Profile Analysis
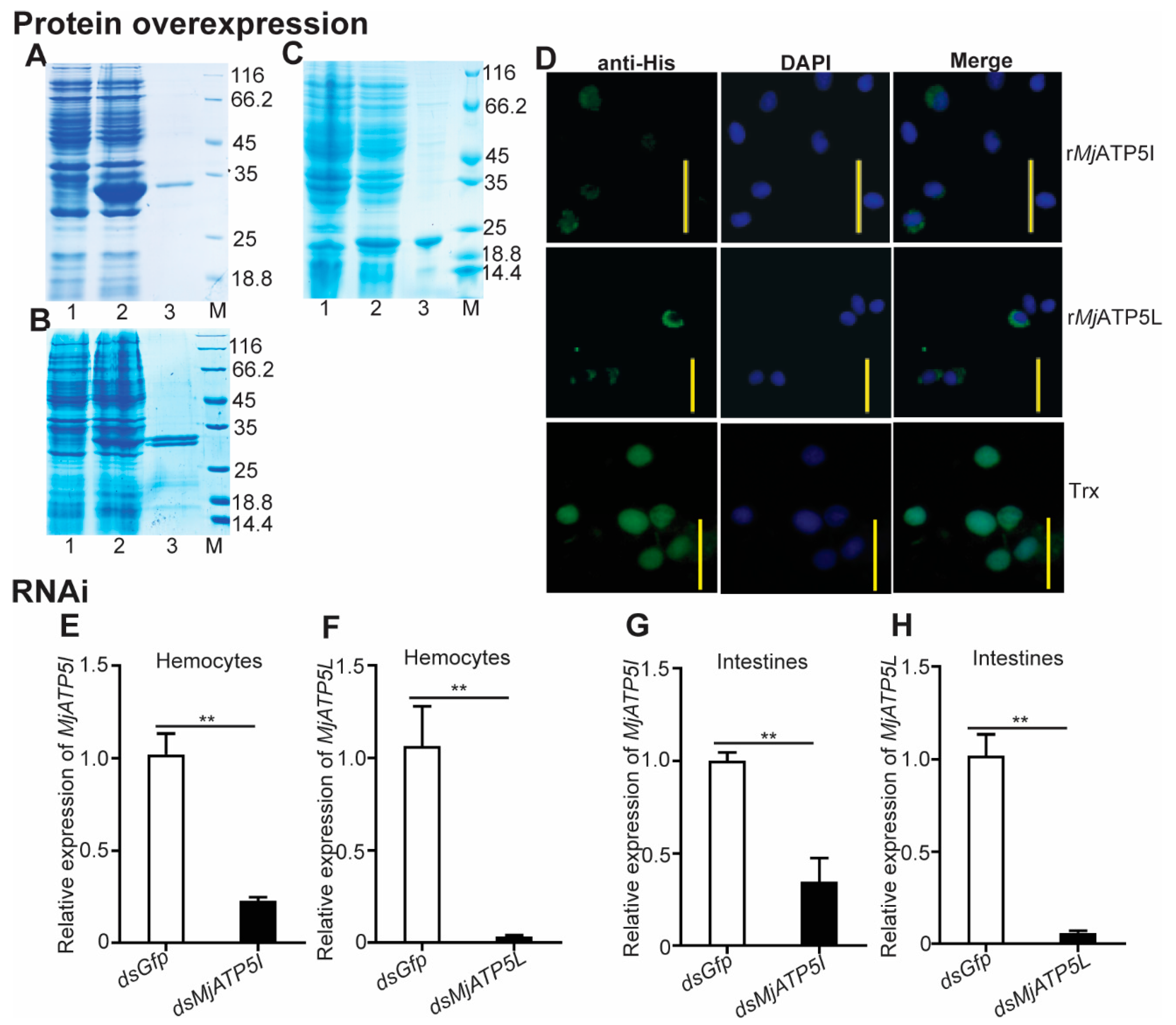
2.3. Recombinant Protein Expression and Purification
2.4. Protein Injection Assay
2.5. RNA Interference
2.6. Survival Rate Assay
2.7. ATP Content Detection Assay
2.8. The Expression Analysis of MjATP5I and MjATP5L after Relish Knockdown
3. Results
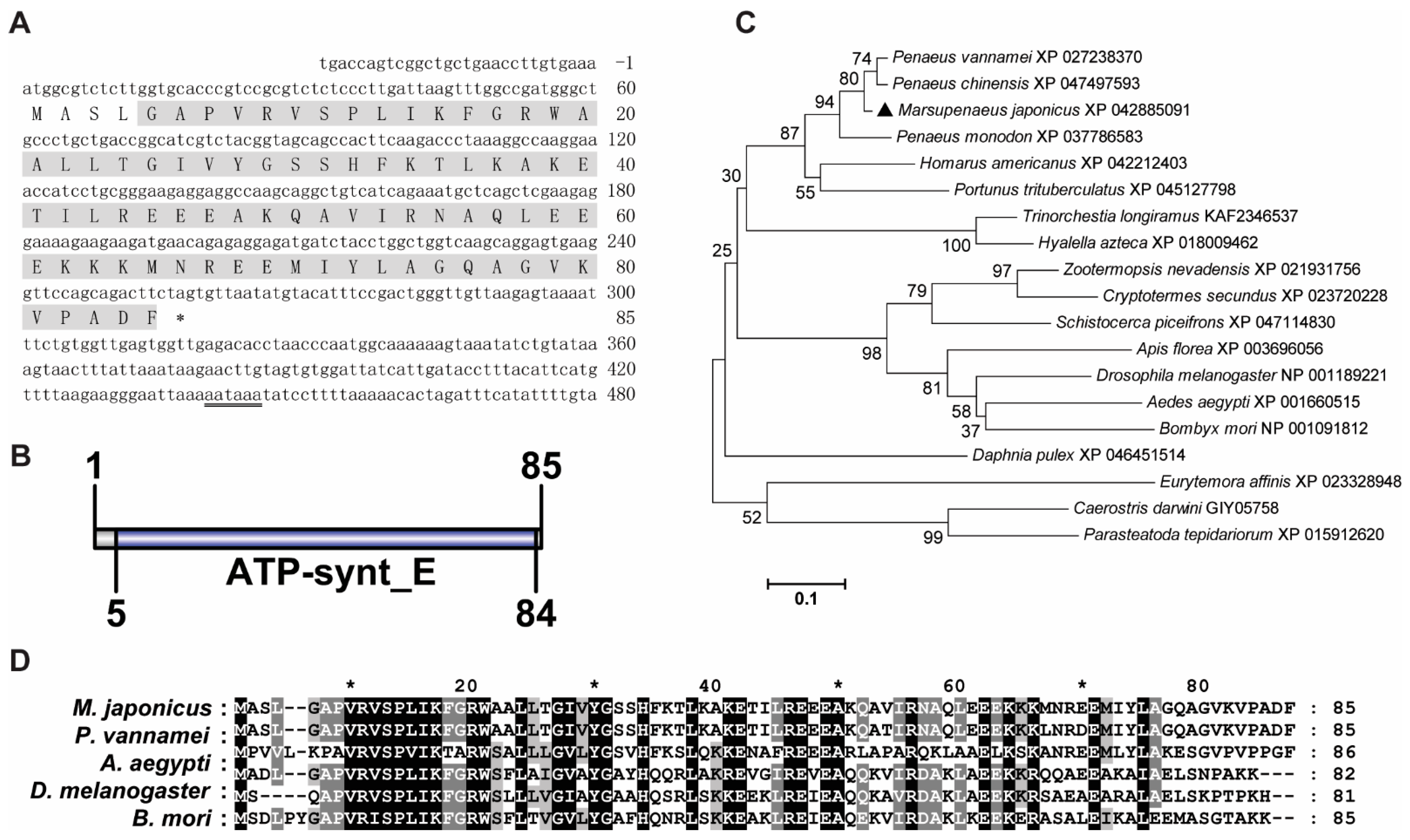
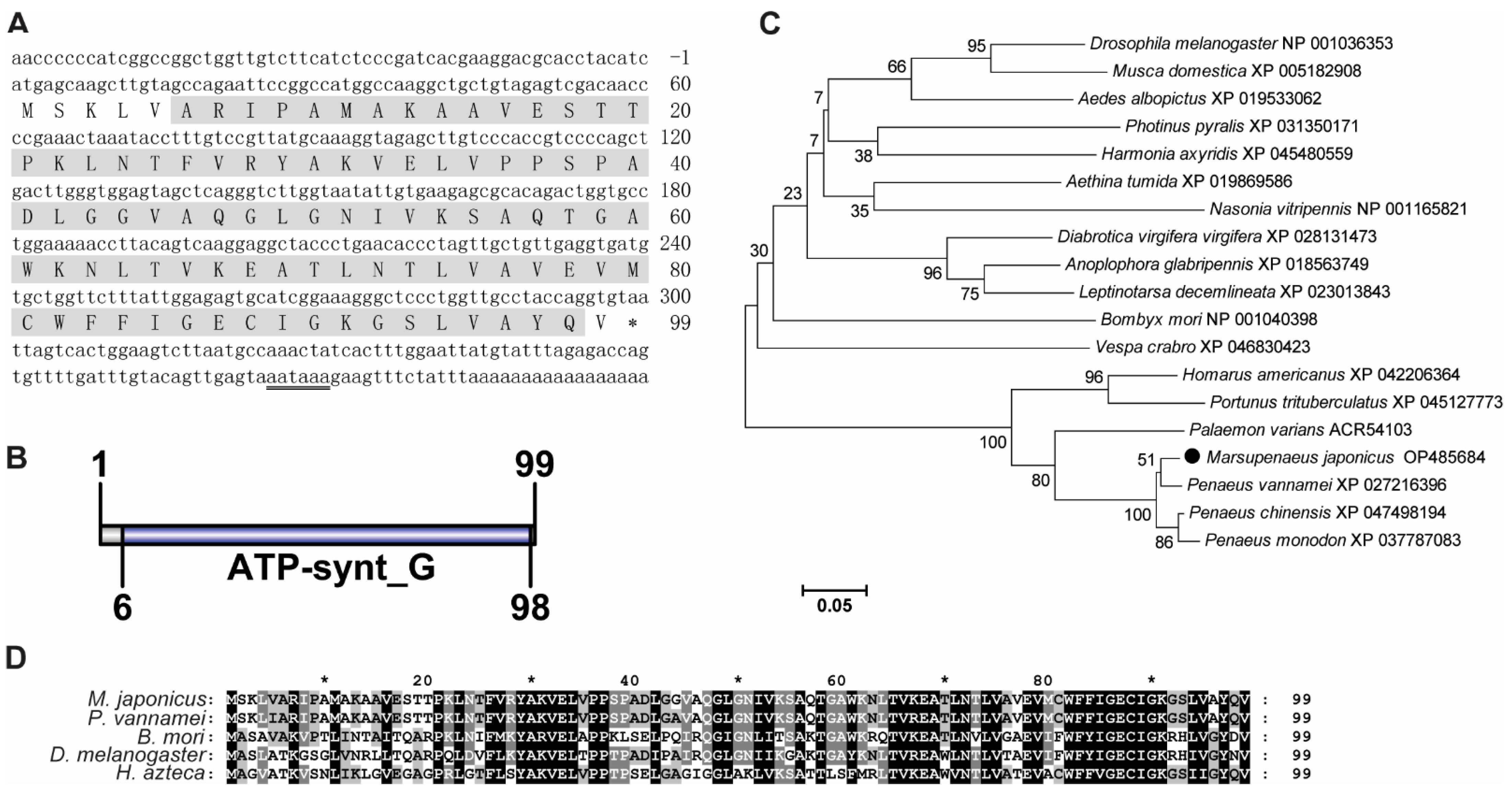
3.1. Sequence Analysis of MjATP5I and MjATP5L
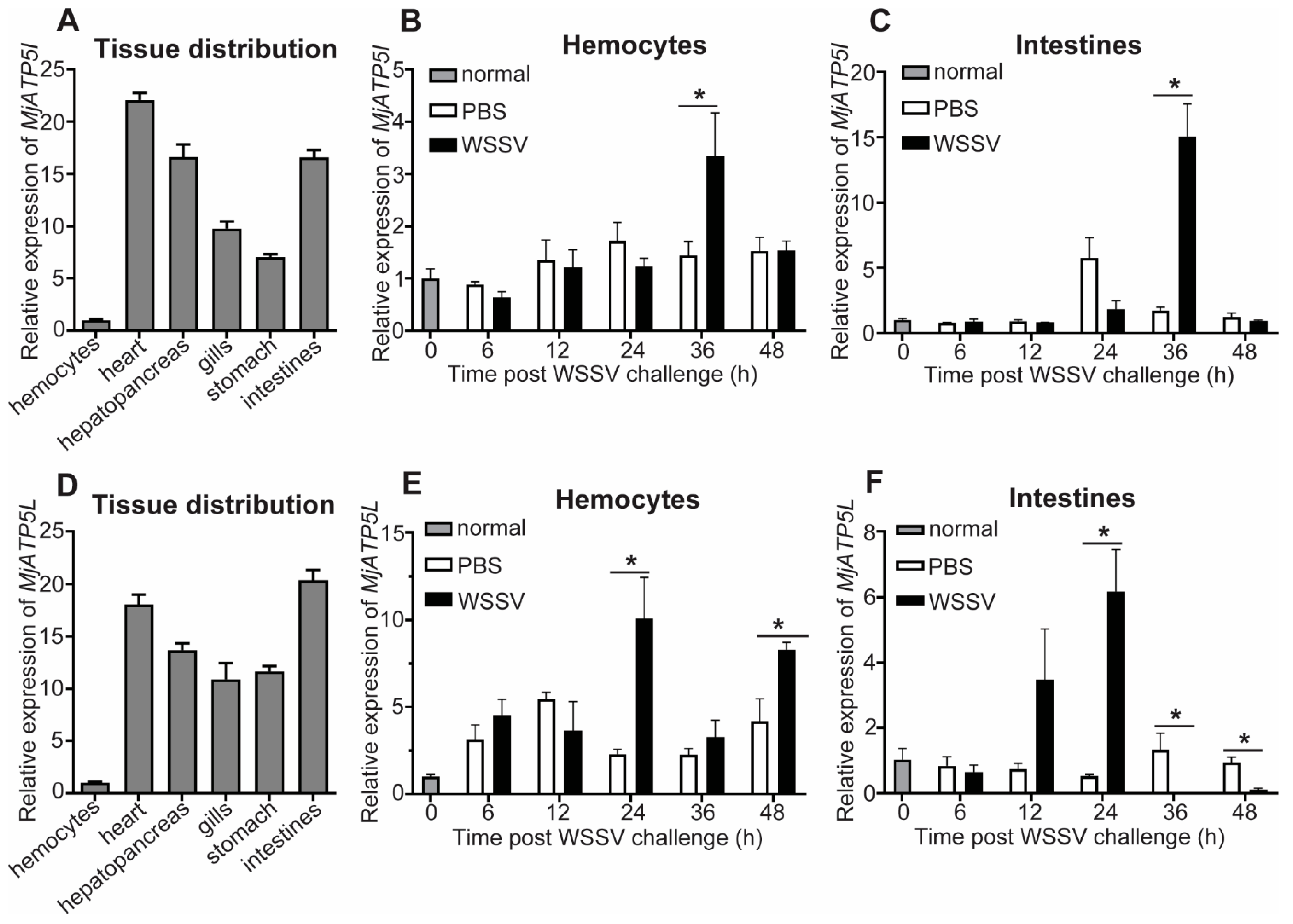
3.2. Tissue Distribution and Expression Profiles of MjATP5I and MjATP5L
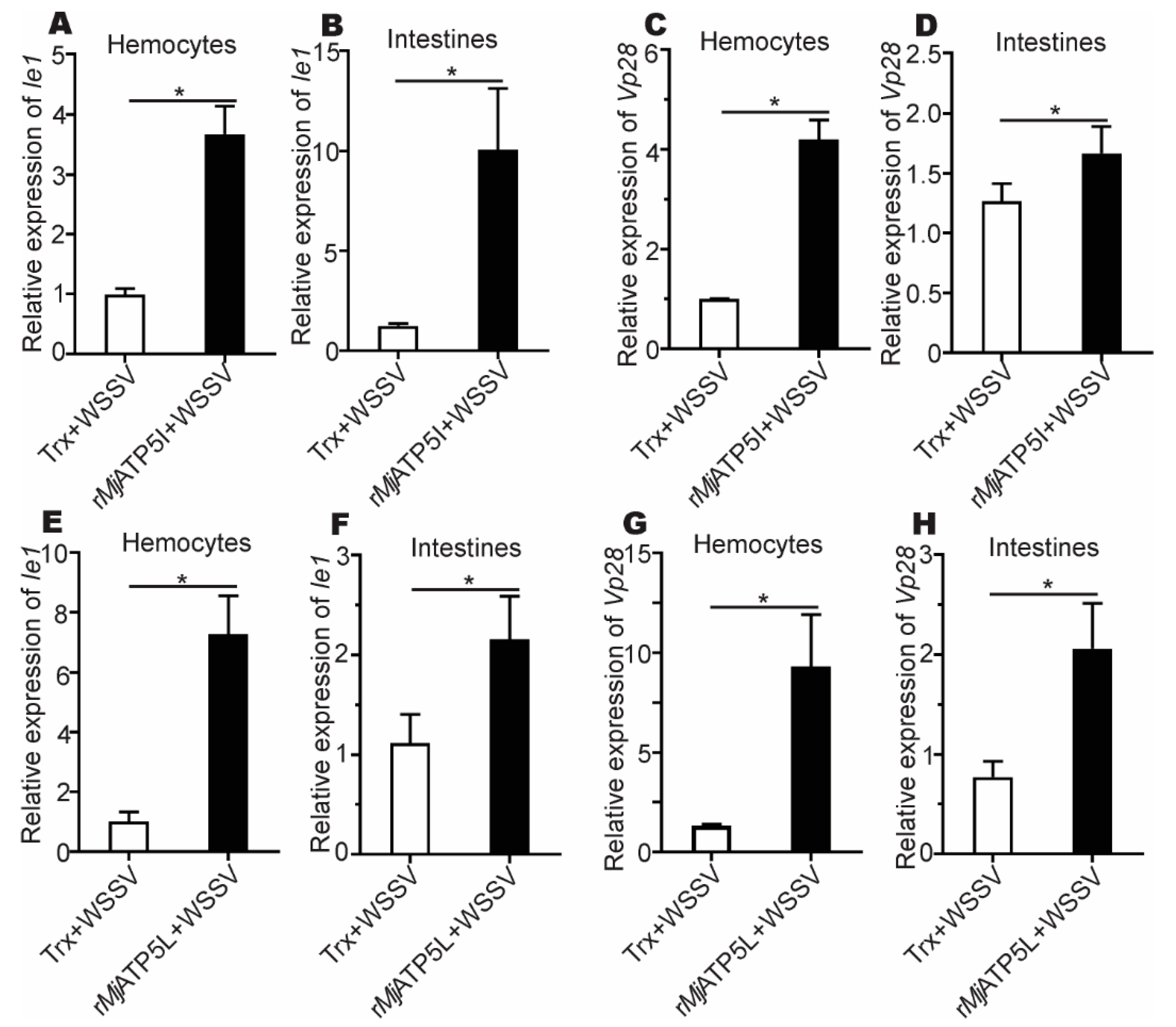
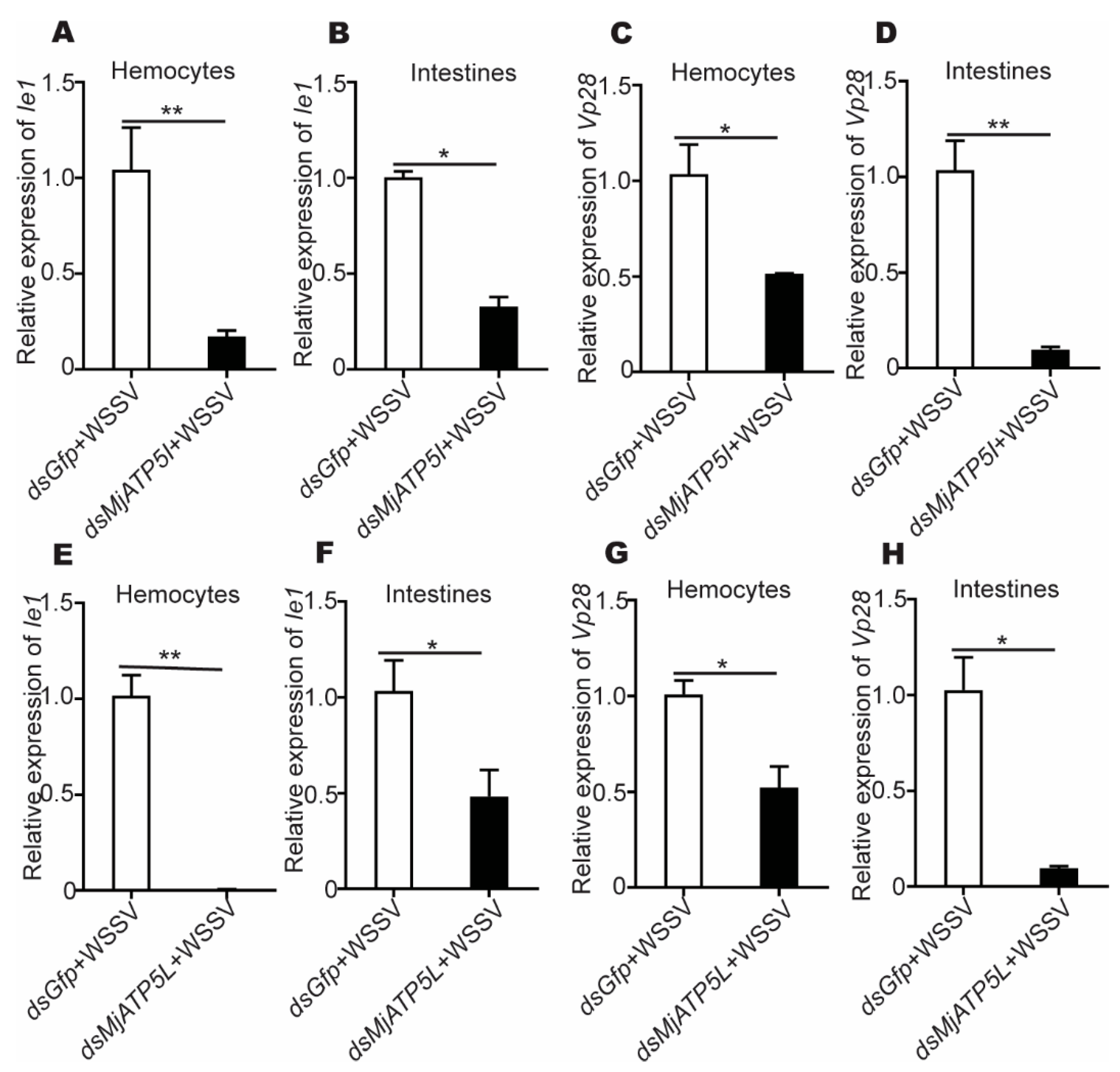
3.3. MjATP5I and MjATP5L Were Beneficial for WSSV Duplication
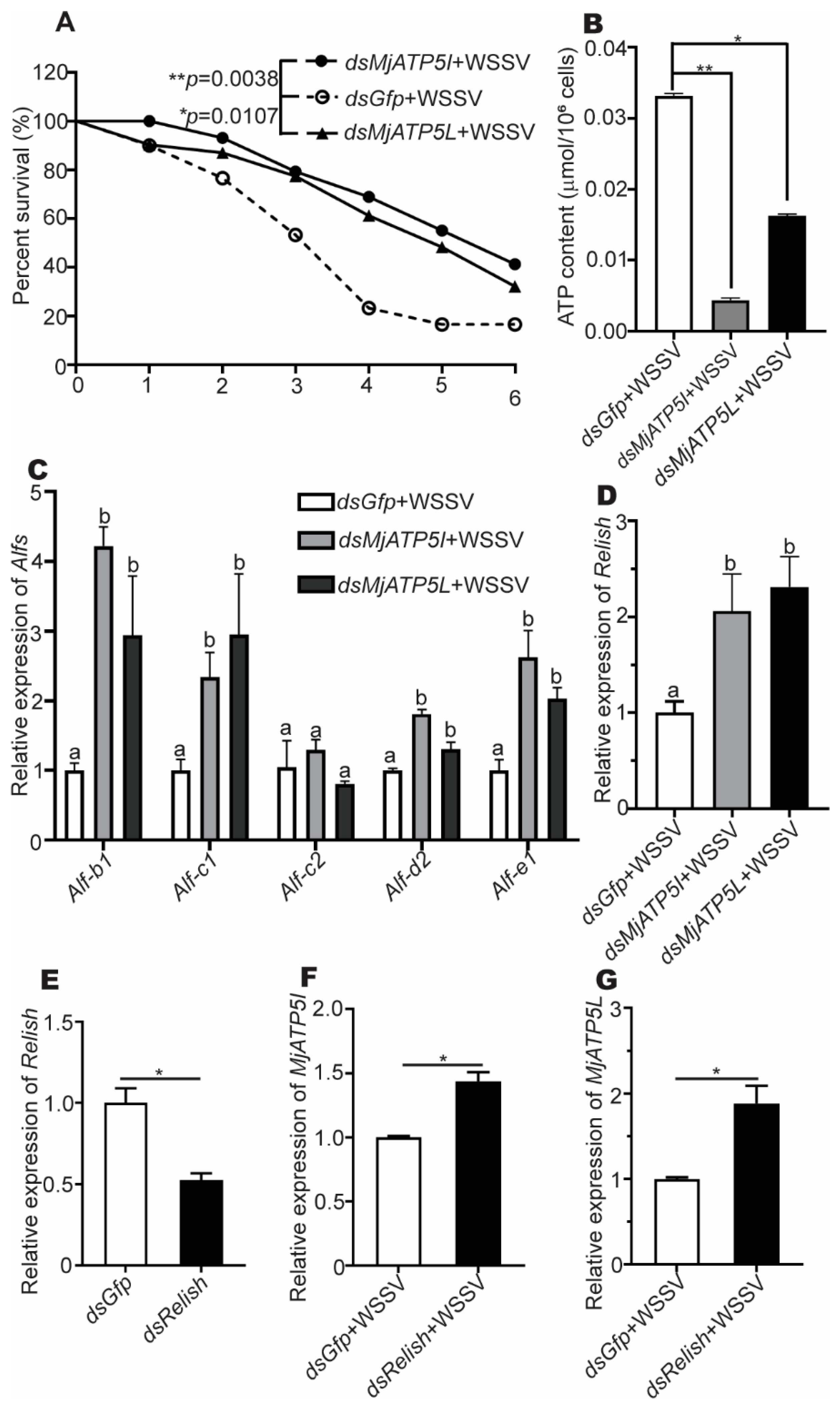
3.4. MjATP5I and MjATP5L Influenced the Shrimp Survival Rate and the Production of ATP
3.5. MjATP5I and MjATP5L Could Regulate the Expression of AMP Genes in Shrimp
3.6. MjATP5I/MjATP5L and Relish Negatively Regulated Each Other
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hong, P.P.; Yang, M.C.; Zhao, X.F.; Wang, J.X. FOXO regulates the expression of antimicrobial peptides and promotes phagocytosis of hemocytes in shrimp antibacterial immunity. PLoS Pathog. 2021, 17, e1009479. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, X.W.; Sun, J.J.; Wang, L.; Zhang, H.W.; Zhao, X.F.; Wang, J.X. Akirin interacts with Bap60 and 14-3-3 proteins to regulate the expression of antimicrobial peptides in the kuruma shrimp (Marsupenaeus japonicus). Dev. Comp. Immunol. 2016, 55, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.J.; Xu, S.; He, Z.H.; Shi, X.Z.; Zhao, X.F.; Wang, J.X. Activation of Toll pathway is different between kuruma shrimp and Drosophila. Front. Immunol. 2017, 8, 1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.S.; Lv, L.X.; Wang, J.X. Anti-lipopolysaccharide factor D from kuruma shrimp exhibits antiviral activity. Mar. Life Sci. Tech. 2022, 4, 52–61. [Google Scholar] [CrossRef]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Functional diversity of anti-lipopolysaccharide factor isoforms in shrimp and their characters related to antiviral activity. Mar. Drugs 2015, 13, 2602–2616. [Google Scholar] [CrossRef] [Green Version]
- Aspden, J.L.; Eyre-Walker, Y.C.; Phillips, R.J.; Amin, U.; Mumtaz, M.A.; Brocard, M.; Couso, J.P. Extensive translation of small Open Reading Frames revealed by Poly-Ribo-Seq. Elife 2014, 3, e03528. [Google Scholar] [CrossRef]
- Saghatelian, A.; Couso, J.P. Discovery and characterization of smORF-encoded bioactive polypeptides. Nat. Chem. Biol. 2015, 11, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, J.P.; Tobias-Santos, V.; Rodrigues, A.C.; Mury, F.B.; da Fonseca, R.N. small ORFs: A new class of essential genes for development. Genet. Mol. Biol. 2015, 38, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Couso, J.P. Finding smORFs: Getting closer. Genome Biol. 2015, 16, 189. [Google Scholar] [CrossRef]
- Srinivas, V.; Kumar, M.; Noronha, S.; Patankar, S. ORFpred: A Machine Learning Program to Identify Translatable Small Open Reading Frames in Intergenic Regions of the Plasmodium falciparum Genome. Curr. Bioinform. 2016, 11, 259–268. [Google Scholar] [CrossRef]
- Couso, J.P.; Patraquim, P. Classification and function of small open reading frames. Nat. Rev. Mol. Cell Biol. 2017, 18, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Malekos, E.; Carpenter, S. Short open reading frame genes in innate immunity: From discovery to characterization. Trends Immunol. 2022, 43, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Formentini, L.; Pereira, M.P.; Sanchez-Cenizo, L.; Santacatterina, F.; Lucas, J.J.; Navarro, C.; Martinez-Serrano, A.; Cuezva, J.M. In vivo inhibition of the mitochondrial H+-ATP synthase in neurons promotes metabolic preconditioning. EMBO J. 2014, 33, 762–778. [Google Scholar] [CrossRef] [Green Version]
- Havlickova, V.; Kaplanova, V.; Nuskova, H.; Drahota, Z.; Houstek, J. Knockdown of F1 epsilon subunit decreases mitochondrial content of ATP synthase and leads to accumulation of subunit c. Biochim. Biophys. Acta 2010, 1797, 1124–1129. [Google Scholar] [CrossRef] [Green Version]
- Bornhovd, C.; Vogel, F.; Neupert, W.; Reichert, A.S. Mitochondrial membrane potential is dependent on the oligomeric state of F1F0-ATP synthase supracomplexes. J. Biol. Chem. 2006, 281, 13990–13998. [Google Scholar] [CrossRef] [Green Version]
- Collinson, I.R.; Runswick, M.J.; Buchanan, S.K.; Fearnley, I.M.; Skehel, J.M.; van Raaij, M.J.; Griffiths, D.E.; Walker, J.E. Fo membrane domain of ATP synthase from bovine heart mitochondria: Purification, subunit composition, and reconstitution with F1-ATPase. Biochemistry 1994, 33, 7971–7978. [Google Scholar] [CrossRef]
- Leyva, J.A.; Bianchet, M.A.; Amzel, L.M. Understanding ATP synthesis: Structure and mechanism of the F1-ATPase (Review). Mol. Membr. Biol. 2009, 20, 27–33. [Google Scholar] [CrossRef]
- Cha, M.Y.; Cho, H.J.; Kim, C.; Jung, Y.O.; Kang, M.J.; Murray, M.E.; Hong, H.S.; Choi, Y.J.; Choi, H.; Kim, D.K.; et al. Mitochondrial ATP synthase activity is impaired by suppressed O-GlcNAcylation in Alzheimer’s disease. Hum. Mol. Genet. 2015, 24, 6492–6504. [Google Scholar] [CrossRef] [Green Version]
- Sardin, E.; Donadello, S.; di Rago, J.P.; Tetaud, E. Biochemical investigation of a human pathogenic mutation in the nuclear ATP5E gene using yeast as a model. Front. Genet. 2015, 6, 159. [Google Scholar] [CrossRef]
- Mayr, J.A.; Havlickova, V.; Zimmermann, F.; Magler, I.; Kaplanova, V.; Jesina, P.; Pecinova, A.; Nuskova, H.; Koch, J.; Sperl, W.; et al. Mitochondrial ATP synthase deficiency due to a mutation in the ATP5E gene for the F1 epsilon subunit. Hum. Mol. Genet. 2010, 19, 3430–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Cheng, J.J.; Yang, B.; Huang, J. The role of F1 ATP synthase beta subunit in WSSV infection in the shrimp, Litopenaeus vannamei. Virol. J. 2010, 7, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Zhu, L.; Tang, X.; Xing, J.; Sheng, X.; Chi, H.; Zhan, W. Differential white spot syndrome virus-binding proteins in two hemocyte subpopulations of Chinese shrimp (Fenneropenaeus chinensis). Dev. Comp. Immunol. 2021, 125, 104215. [Google Scholar] [CrossRef]
- Bustos, D.M.; Velours, J. The modification of the conserved GXXXG motif of the membrane-spanning segment of subunit g destabilizes the supramolecular species of yeast ATP synthase. J. Biol. Chem. 2005, 280, 29004–29010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, H.; Yu, Y.; Xu, Y. Antisense of ATP Synthase Subunit e Inhibits the Growth of Human Hepatocellular Carcinoma Cells. Oncol. Res. 2001, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Q.; Huang, Z.; Chen, X.; Yang, C.; Zhang, Y.; Zhao, Y.; Wang, F. ATP synthase subunit e is a shrimp growth-associated breeding marker. Genomics 2022, 114, 110410. [Google Scholar] [CrossRef]
- Dahiya, N.; Sarachana, T.; Kulkarni, S.; Wood, W.H., III; Zhang, Y.; Becker, K.G.; Wang, B.D.; Atreya, C.D. miR-570 interacts with mitochondrial ATPase subunit g (ATP5L) encoding mRNA in stored platelets. Platelets 2017, 28, 74–81. [Google Scholar] [CrossRef]
- Boyle, G.M.; Roucou, X.; Nagley, P.; Devenish, R.J.; Prescott, M. Identification of subunit g of yeast mitochondrial F1F0-ATP synthase, a protein required for maximal activity of cytochrome c oxidase. Eur. J. Biochem. 1999, 262, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Yang, C.; Li, Q.; Zhu, W.; Chen, X.; Peng, M.; Chen, X.; Lin, Y.; Wang, H.; Liu, H.; et al. Identification of a quantitative trait loci (QTL) associated with ammonia tolerance in the Pacific white shrimp (Litopenaeus vannamei). BMC Genom. 2020, 21, 857. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Lu, Y.; Zhuang, Y.; Liu, J. Mining antimicrobial peptides from small open reading frames in Ciona intestinalis. J. Pept. Sci. 2014, 20, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Zanet, J.; Chanut-Delalande, H.; Plaza, S.; Payre, F. Small Peptides as Newcomers in the Control of Drosophila Development. Curr. Top. Dev. Biol. 2016, 117, 199–219. [Google Scholar] [PubMed]
- Garone, C.; Pietra, A.; Nesci, S. From the Structural and (Dys)Function of ATP Synthase to Deficiency in Age-Related Diseases. Life 2022, 12, 401. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liang, H.; Liao, X.; Pan, J.; Chen, J.; Zhao, S.; Xu, Y.; Wu, Y.; Ni, J. A humanized chimeric antibody Hai178 targeted to the beta subunit of F1F0 ATP synthase. Tumour Biol. 2016, 37, 15903–15912. [Google Scholar] [CrossRef]
- Lin, X.; Kim, Y.A.; Lee, B.L.; Soderhall, K.; Soderhall, I. Identification and properties of a receptor for the invertebrate cytokine astakine, involved in hematopoiesis. Exp. Cell Res. 2009, 315, 1171–1180. [Google Scholar] [CrossRef]
- He, N.; Qin, Q.; Xu, X. Differential profile of genes expressed in hemocytes of White Spot Syndrome Virus-resistant shrimp (Penaeus japonicus) by combining suppression subtractive hybridization and differential hybridization. Antivir. Res. 2005, 66, 39–45. [Google Scholar] [CrossRef]
- Rosa, R.D.; Barracco, M.A. Shrimp interferon is rather a portion of the mitochondrial F0-ATP synthase than a true alpha-interferon. Mol. Immunol. 2008, 45, 3490–3493. [Google Scholar] [CrossRef]
- Wang, H.C.; Wang, H.C.; Leu, J.H.; Kou, G.H.; Wang, A.H.; Lo, C.F. Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Dev. Comp. Immunol. 2007, 31, 672–686. [Google Scholar] [CrossRef]
- Bourchookarn, A.; Havanapan, P.O.; Thongboonkerd, V.; Krittanai, C. Proteomic analysis of altered proteins in lymphoid organ of yellow head virus infected Penaeus monodon. Biochim. Biophys. Acta 2008, 1784, 504–511. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, M.L.; Wang, X.W.; Gao, X.X.; Cheng, J.J.; Li, C.; Huang, J. ATP synthesis is active on the cell surface of the shrimp Litopenaeus vannamei and is suppressed by WSSV infection. Virol. J. 2015, 12, 49. [Google Scholar] [CrossRef]
- Huo, L.J.; Yang, M.C.; Wang, J.X.; Shi, X.Z. Mitochondrial ATPase inhibitor factor 1, MjATPIF1, is beneficial for WSSV replication in kuruma shrimp (Marsupenaeus japonicus). Fish Shellfish Immunol. 2020, 98, 245–254. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Gene Target and GenBank Accession No. | Sequence (5′–3′) |
|---|---|---|
| MjATP5I-RTF | MjATP5I XP_042885091 | gccacttcaagaccctaa |
| MjATP5I-RTR | tctgctggaaccttcact | |
| MjATP5L-RTF | MjATP5L OP485684 | ccccgaaactaaatacct |
| MjATP5L-RTR | gaaccagcacatcacctc | |
| MjATP5I-RNAiF | taatacgactcactataggctcccttgattaagtttgg | |
| MjATP5I-RNAiR | taatacgactcactataggcattgggttaggtgtctc | |
| MjATP5L-RNAiF | taatacgactcactataggccgttatgcaaaggtagagc | |
| MjATP5L-RNAiR | taatacgactcactataggtggtaggcaaccagggag | |
| MjATP5I-RTF1 | gccacttcaagaccctaa | |
| MjATP5I-RTR1 | tctgctggaaccttcact | |
| MjATP5L-RTF1 | ccccgaaactaaatacct | |
| MjATP5L-RTR1 | gaaacttctttatttactca | |
| Gfp-RNAiF | Gfp MK371210.1 | gcgtaatacgactcactataggtggtcccaattctcgtggaac |
| Gfp-RNAiR | gcgtaatacgactcactataggcttgaagttgaccttgatgcc | |
| Relish-RNAiF | Relish MN607236 | taatacgactcactatagggtcggaggaaggttctcacggc |
| Relish-RNAiR | taatacgactcactatagggtctcctttcctgaccttgtctgtg | |
| Relish-RTF | acgcagaagaatgctgattg | |
| Relish-RTR | attcccccaccaacagc | |
| MjATP5I-ExF | tacgaattcatggcgtctcttggtgcaccc | |
| MjATP5I-ExR | tacctcgagctagaagtctgctggaacctt | |
| MjATP5L-ExF | tacgaattcatgagcaagcttgtagccaga | |
| MjATP5L-ExR | tacgcggccgcttacacctggtaggcaaccag | |
| Ie1-RTF | Ie1 | gactctacaaatctctttgcca |
| Ie1-RTR | 926915 | ctacctttgcaccaattgctag |
| Vp28-RTF | Vp28 | agctccaacacctcctccttca |
| Vp28-RTR | 927064 | ttactcggtctcagtgccaga |
| Alf-b1-RT-F | Alf-b1 KY627759 | cggtggtggccctggtggcactcttcg |
| Alf-b1-RTR | gactggctgcgtgtgctggcttcccctc | |
| Alf-c1-RTF | Alf-c1 KU213608 | cgcttcaagggtcggatgtg |
| Alf-c1-RTR | cgagcctcttcctccgtgatg | |
| Alf-c2-RTF | Alf-c2 KU160498 | tcctggtggtggcagtggct |
| Alf-c2-RTR | tgcgggtctcggcttctcct | |
| Alf-d2-RTF | Alf-d2 MT977630 | cgcaggcttatggaggac |
| Alf-d2-RT-R | aggtgacagtgccgagga | |
| Alf-e1-RTF | Alf-e1 KY627760 | tcctaaccacgcagtgctttgctaatg |
| Alf-e1-RTR | gcttttcggatttgccttcgatgtttg | |
| actin-RTF | β-actin GU645235 | cagccttccttcctgggtatgg |
| actin-RTR | gagggagcgagggcagtgatt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, L.-J.; Lu, P.-Y.; Li, D.-X.; Shi, X.-Z. The sORF-Encoded Peptides, ATP Synthase Subunits, Facilitate WSSV Duplication in Shrimp. Viruses 2022, 14, 2449. https://doi.org/10.3390/v14112449
Huo L-J, Lu P-Y, Li D-X, Shi X-Z. The sORF-Encoded Peptides, ATP Synthase Subunits, Facilitate WSSV Duplication in Shrimp. Viruses. 2022; 14(11):2449. https://doi.org/10.3390/v14112449
Chicago/Turabian StyleHuo, Li-Jie, Peng-Yuan Lu, Dian-Xiang Li, and Xiu-Zhen Shi. 2022. "The sORF-Encoded Peptides, ATP Synthase Subunits, Facilitate WSSV Duplication in Shrimp" Viruses 14, no. 11: 2449. https://doi.org/10.3390/v14112449
APA StyleHuo, L. -J., Lu, P. -Y., Li, D. -X., & Shi, X. -Z. (2022). The sORF-Encoded Peptides, ATP Synthase Subunits, Facilitate WSSV Duplication in Shrimp. Viruses, 14(11), 2449. https://doi.org/10.3390/v14112449