
Spatial Methods for Inferring Extremes in Dengue Outbreak Risk in Singapore

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. Max-Stable Processes
2.2.1. Max-Stable Models: Smith Model
2.2.2. Max-Stable Models: Schlather Model
2.2.3. Max-Stable Models: Brown–Resnick Model
2.2.4. Modeling Trend Surfaces
2.2.5. Model Assessment
2.2.6. Model Fitting
3. Results
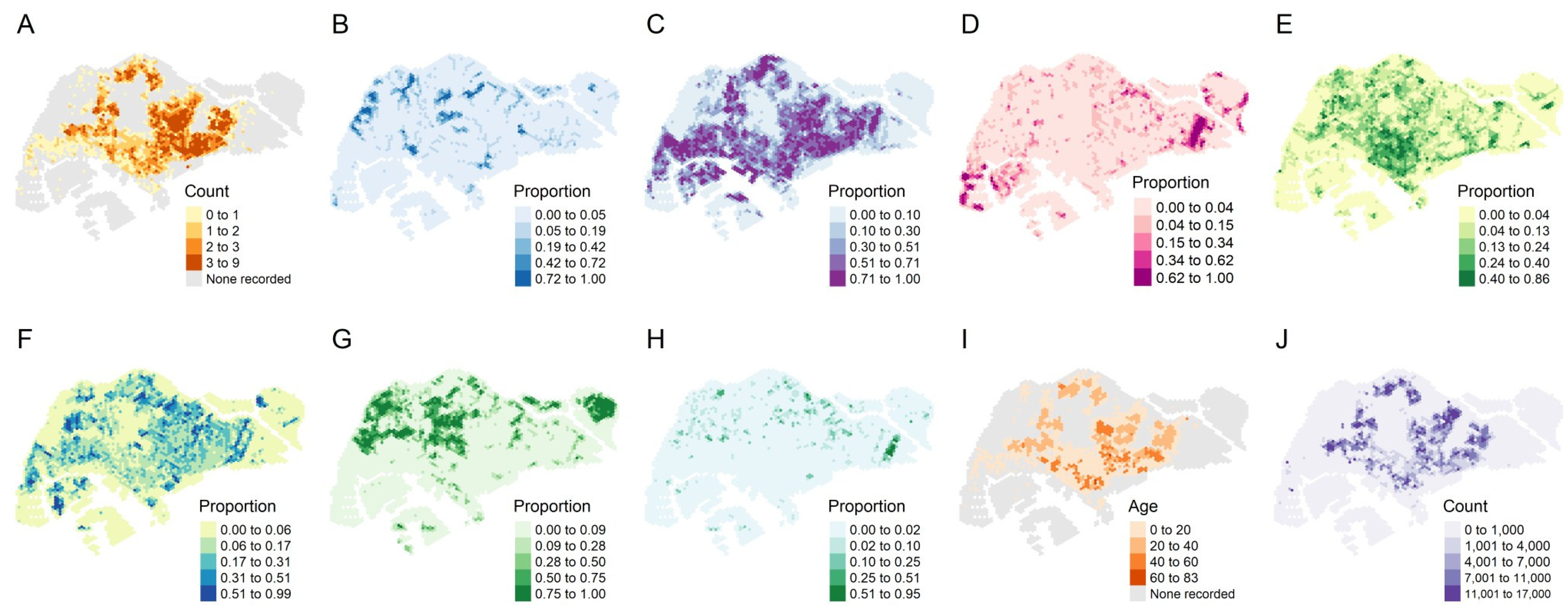
3.1. Descriptive Results
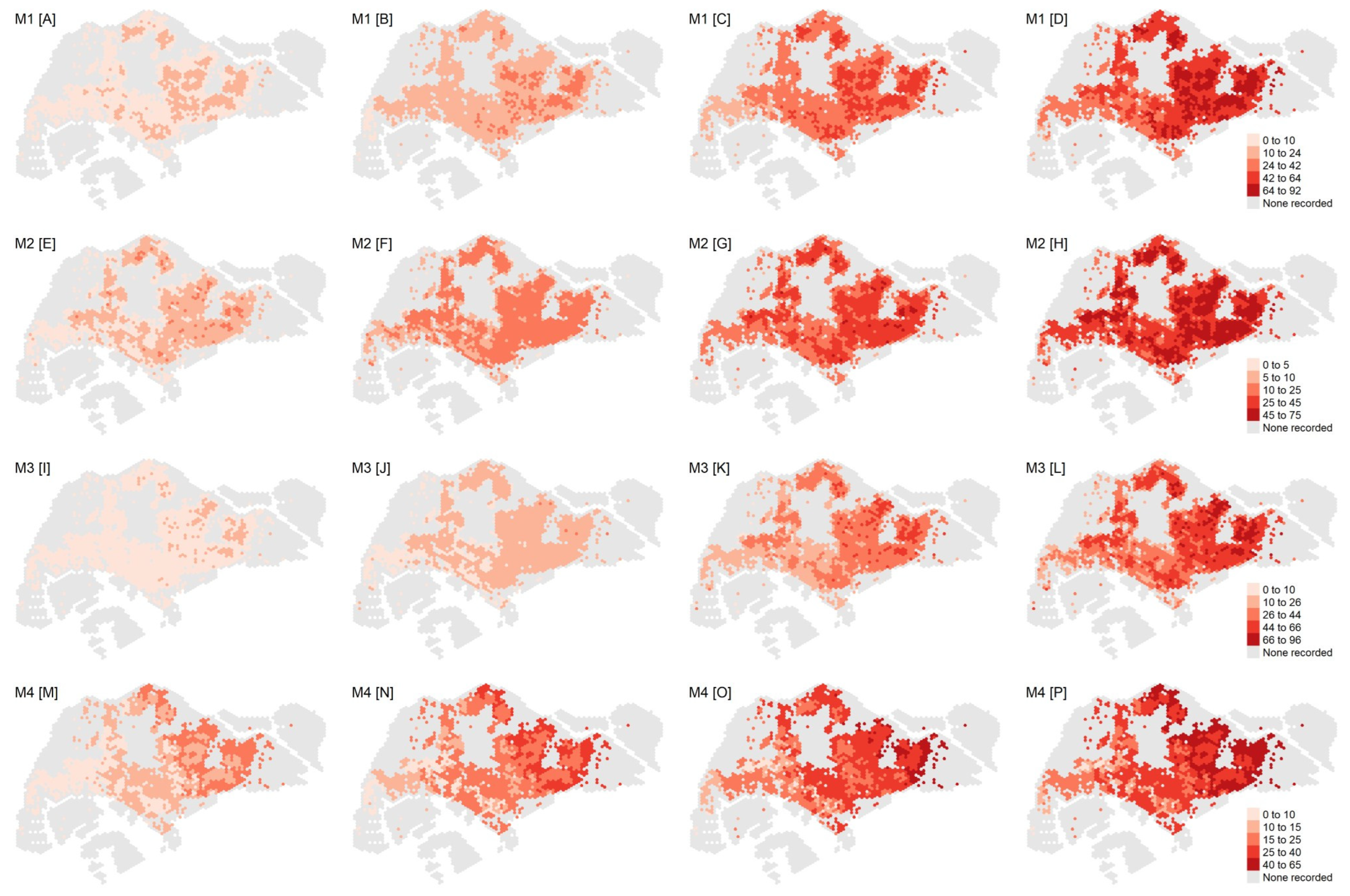
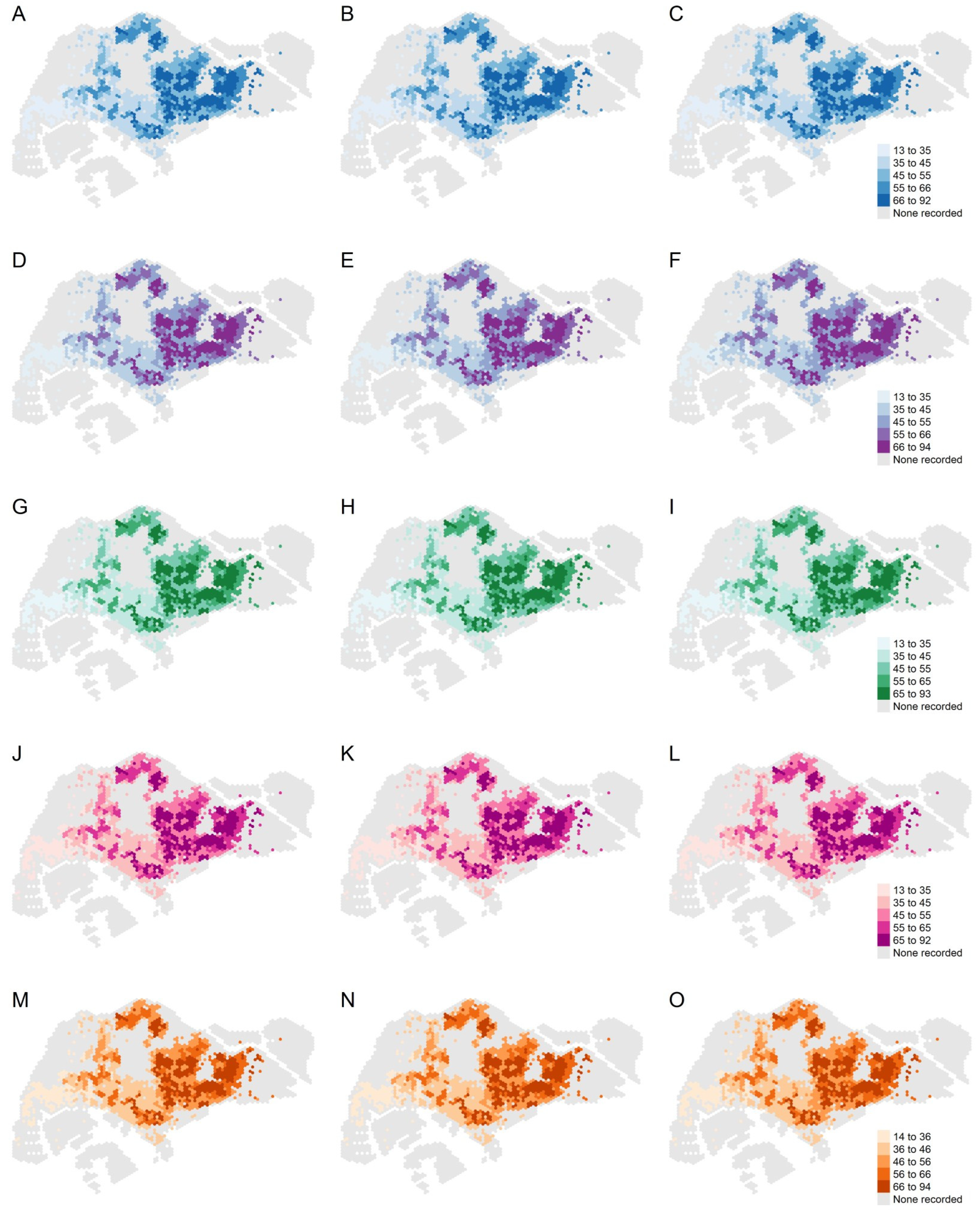
3.2. Model Assessment
3.3. Estimated Extreme Dengue Outbreak Risk across Space and Associated Spatial Drivers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, J.T.; Han, Y.; Sue Lee Dickens, B.; Ng, L.C.; Cook, A.R. Time varying methods to infer extremes in dengue transmission dynamics. PLoS Comput. Biol. 2020, 16, e1008279. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Lemaitre, M.; Wilson, M.L.; Viboud, C.; Yordanov, Y.; Wackernagel, H.; Carrat, F. Applications of Extreme Value Theory in Public Health. PLoS ONE 2016, 11, e0159312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, C.S.; Huynh, M.P.; Schroeder, M.; Anyatonwu, S.; Dahlgren, F.S.; Danyluk, G.; Fernandez, D.; Greene, S.K.; Kipshidze, N.; Liu, L.; et al. Applying infectious disease forecasting to public health: A path forward using influenza forecasting examples. BMC Public Health 2019, 19, 1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Liu, X.; Kok, S.-Y.; Rajarethinam, J.; Liang, S.; Yap, G.; Chong, C.-S.; Lee, K.-S.; Tan, S.S.Y.; Chin, C.K.Y.; et al. Three-Month Real-Time Dengue Forecast Models: An Early Warning System for Outbreak Alerts and Policy Decision Support in Singapore. Environ. Health Perspect. 2016, 124, 1369–1375. [Google Scholar] [CrossRef]
- Earnest, A.; Tan, S.B.; Wilder-Smith, A.; Machin, D. Comparing statistical models to predict dengue fever notifications. Comput. Math. Methods Med. 2012, 2012, 758674. [Google Scholar] [CrossRef]
- Hii, Y.L.; Zhu, H.; Ng, N.; Ng, L.C.; Rocklöv, J. Forecast of Dengue Incidence Using Temperature and Rainfall. PLoS Negl. Trop. Dis. 2012, 6, e1908. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.N. Epidemiological models are important tools for guiding COVID-19 interventions. BMC Med. 2020, 18, 152. [Google Scholar] [CrossRef]
- Chowell, G.; Sattenspiel, L.; Bansal, S.; Viboud, C. Mathematical models to characterize early epidemic growth: A review. Phys. Life Rev. 2016, 18, 66–97. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, P.; Taleb, N.N. Tail risk of contagious diseases. Nat. Phys. 2020, 16, 606–613. [Google Scholar] [CrossRef]
- Yamana, T.K.; Kandula, S.; Shaman, J. Superensemble forecasts of dengue outbreaks. J. R. Soc. Interface 2016, 13, 20160410. [Google Scholar] [CrossRef]
- Finkenstädt, B.F.; Grenfell, B.T. Time series modelling of childhood diseases: A dynamical systems approach. J. R. Stat. Soc. Ser. C Appl. Stat. 2000, 49, 187–205. [Google Scholar] [CrossRef]
- Van de Vyver, H. Spatial regression models for extreme precipitation in Belgium. Water Resour. Res. 2012, 48, 2011WR011707. [Google Scholar] [CrossRef]
- Scotto, M.G.; Barbosa, S.M.; Alonso, A.M. Extreme value and cluster analysis of European daily temperature series. J. Appl. Stat. 2011, 38, 2793–2804. [Google Scholar] [CrossRef] [Green Version]
- Kislov, A.; Matveeva, T. An Extreme Value Analysis of Wind Speed over the European and Siberian Parts of Arctic Region. Atmos. Clim. Sci. 2016, 06, 205–223. [Google Scholar] [CrossRef] [Green Version]
- Cattarino, L.; Rodriguez-Barraquer, I.; Imai, N.; Cummings, D.A.T.; Ferguson, N.M. Mapping global variation in dengue transmission intensity. Sci. Transl. Med. 2020, 12, eaax4144. [Google Scholar] [CrossRef]
- Chen, Y.; Ong, J.H.Y.; Rajarethinam, J.; Yap, G.; Ng, L.C.; Cook, A.R. Neighbourhood level real-time forecasting of dengue cases in tropical urban Singapore. BMC Med. 2018, 16, 129. [Google Scholar] [CrossRef] [Green Version]
- Department of Statistics, Singapore Population and Population Structure. 2020; [Internet]. Available online: https://www.singstat.gov.sg/find-data/search-by-theme/population/459population-and-population-structure/latest-data (accessed on 24 September 2020).
- Gubler, D.J. Dengue, Urbanization and Globalization: The Unholy Trinity of the 21(st) Century. Trop. Med. Health 2011, 39, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Rajarethinam, J.; Ang, L.-W.; Ong, J.; Ycasas, J.; Hapuarachchi, H.C.; Yap, G.; Chong, C.-S.; Lai, Y.-L.; Cutter, J.; Ho, D.; et al. Dengue in Singapore from 2004 to 2016: Cyclical Epidemic Patterns Dominated by Serotypes 1 and 2. Am. J. Trop. Med. Hyg. 2018, 99, 204–210. [Google Scholar] [CrossRef]
- Miller, J.C. Spread of infectious disease through clustered populations. J. R. Soc. Interface 2009, 6, 1121–1134. [Google Scholar] [CrossRef] [Green Version]
- Mammen, M.P.; Pimgate, C.; Koenraadt, C.J.M.; Rothman, A.L.; Aldstadt, J.; Nisalak, A.; Jarman, R.G.; Jones, J.W.; Srikiatkhachorn, A.; Ypil-Butac, C.A.; et al. Spatial and temporal clustering of dengue virus transmission in Thai villages. PLoS Med. 2008, 5, e205. [Google Scholar] [CrossRef]
- Yoon, I.-K.; Getis, A.; Aldstadt, J.; Rothman, A.L.; Tannitisupawong, D.; Koenraadt, C.J.M.; Fansiri, T.; Jones, J.W.; Morrison, A.C.; Jarman, R.G.; et al. Fine scale spatiotemporal clustering of dengue virus transmission in children and Aedes aegypti in rural Thai villages. PLoS Negl. Trop. Dis. 2012, 6, e1730. [Google Scholar] [CrossRef] [PubMed]
- Seidahmed, O.M.E.; Lu, D.; Chong, C.S.; Ng, L.C.; Eltahir, E.A.B. Patterns of Urban Housing Shape Dengue Distribution in Singapore at Neighborhood and Country Scales. GeoHealth 2018, 2, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Vythilingam, I.; Chong, C.-S.; Abdul Razak, M.A.; Tan, C.-H.; Liew, C.; Pok, K.-Y.; Ng, L.-C. Gravitraps for Management of Dengue Clusters in Singapore. Am. J. Trop. Med. Hyg. 2013, 88, 888–892. [Google Scholar] [CrossRef]
- Chen, J.; Lei, X.; Zhang, L.; Peng, B. Using Extreme Value Theory Approaches to Forecast the Probability of Outbreak of Highly Pathogenic Influenza in Zhejiang, China. PLoS ONE 2015, 10, e0118521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajarethinam, J.; Ong, J.; Lim, S.-H.; Tay, Y.-H.; Bounliphone, W.; Chong, C.-S.; Yap, G.; Ng, L.-C. Using Human Movement Data to Identify Potential Areas of Zika Transmission: Case Study of the Largest Zika Cluster in Singapore. Int. J. Environ. Res. Public. Health 2019, 16, 808. [Google Scholar] [CrossRef] [Green Version]
- Gaw, L.Y.-F..; Yee, A.T.K.; Richards, D.R. A High-Resolution Map of Singapore’s Terrestrial Ecosystems. Data 2019, 4, 116. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.C.; Padoan, S.A.; Ribatet, M. Statistical Modeling of Spatial Extremes. Stat. Sci. 2012, 27, 161–186. Available online: https://projecteuclid.org/journals/statistical-science/volume-27/issue-2/Statistical-Modeling-of-Spatial-Extremes/10.1214/11-STS376.full (accessed on 1 August 2022). [CrossRef] [Green Version]
- Ribatet, M. Spatial Extremes: Max-stable processes at work. J. Société Fr. Stat. 2013, 154, 156–177. [Google Scholar]
- Ribatet, M. A User’s Guide to the SpatialExtremes Package [Internet]. 2009. Available online: https://cran.r-project.org/web/packages/SpatialExtremes/vignettes/SpatialExtremesGuide.pdf (accessed on 8 January 2022).
- Ribatet, M. SpatialExtremes: Modelling Spatial Extremes [Internet]. Available online: https://cran.r-project.org/web/packages/SpatialExtremes/index.html (accessed on 22 September 2020).
- Sun, H.; Dickens, B.L.; Richards, D.; Ong, J.; Rajarethinam, J.; Hassim, M.E.E.; Lim, J.T.; Carrasco, L.R.; Aik, J.; Yap, G.; et al. Spatio-temporal analysis of the main dengue vector populations in Singapore. Parasit. Vectors 2021, 14, 41. [Google Scholar] [CrossRef]
- Weng, F.; Pu, R. Mapping and assessing of urban impervious areas using multiple endmember spectral mixture analysis: A case study in the city of Tampa, Florida. Geocarto Int. 2013, 28, 594–615. [Google Scholar] [CrossRef] [Green Version]
- Ong, J.; Soh, S.; Ho, S.H.; Seah, A.; Dickens, B.S.; Tan, K.W.; Koo, J.R.; Cook, A.R.; Richards, D.R.; Gaw, L.Y.-F.; et al. Fine-scale estimation of effective reproduction numbers for dengue surveillance. PLOS Comput. Biol. 2022, 18, e1009791. [Google Scholar] [CrossRef] [PubMed]
- Wimberly, M.C.; Davis, J.K.; Evans, M.V.; Hess, A.; Newberry, P.M.; Solano-Asamoah, N.; Murdock, C.C. Land cover affects microclimate and temperature suitability for arbovirus transmission in an urban landscape. PLoS Negl. Trop. Dis. 2020, 14, e0008614. [Google Scholar] [CrossRef] [PubMed]
- Kek, R.; Hapuarachchi, H.C.; Chung, C.-Y.; Humaidi, M.B.; Razak, M.A.B.A.; Chiang, S.; Lee, C.; Tan, C.-H.; Yap, G.; Chong, C.-S.; et al. Feeding host range of Aedes albopictus (Diptera: Culicidae) demonstrates its opportunistic host-seeking behavior in rural Singapore. J. Med. Entomol. 2014, 51, 880–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, F.; Collins, J.J. Evidence that coronavirus superspreading is fat-tailed. Proc. Natl. Acad. Sci. USA 2020, 117, 29416–29418. [Google Scholar] [CrossRef]
- Lin, C.-H.; Wen, T.-H. How Spatial Epidemiology Helps Understand Infectious Human Disease Transmission. Trop. Med. Infect. Dis. 2022, 7, 164. [Google Scholar] [CrossRef]
- Hagenaars, T.J.; Donnelly, C.A.; Ferguson, N.M. Spatial heterogeneity and the persistence of infectious diseases. J. Theor. Biol. 2004, 229, 349–359. [Google Scholar] [CrossRef]
- Lim, J.T.; Dickens, B.S.L.; Cook, A.R. Modelling the epidemic extremities of dengue transmissions in Thailand. Epidemics 2020, 33, 100402. [Google Scholar] [CrossRef]
- Lessler, J.; Salje, H.; Grabowski, M.K.; Cummings, D.A.T. Measuring Spatial Dependence for Infectious Disease Epidemiology. PLoS ONE 2016, 11, e0155249. [Google Scholar] [CrossRef]
- Pollington, T.M.; Tildesley, M.J.; Hollingsworth, T.D.; Chapman, L.A.C. The spatiotemporal tau statistic: A review. arXiv 2019, arXiv:1911.11476. Available online: http://arxiv.org/abs/1911.11476 (accessed on 1 August 2022).
- Team RIP. R-INLA Project [Internet]. Available online: https://www.r-inla.org/home (accessed on 23 September 2021).
- Urban Redevelopment Authority, Singapore Long-Term Planning [Internet]. Available online: https://www.ura.gov.sg/Corporate/Planning/Our-Planning-Process/Long-Term-Planning (accessed on 1 August 2022).




{kind=link}
{kind=link}
{kind=link}
| Model | Restriction |
|---|---|
| Smith | |
| Schlather | where |
| Brown–Resnick | where |
| Model 1 | Trend Surfaces 2 | Relative Deviance 3 |
|---|---|---|
| Brown–Resnick (M1) | 1 | |
| Schlather (M2) | 1.007 | |
| Schlather (M3) | 1.022 | |
| Brown–Resnick (M4) | 1.427 |
| Year (20XX 1) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 07 | 08 | 09 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | |
| Min. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 25th perc. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 50th perc. | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0.5 | 0 | 1 | 2 |
| 75th perc. | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 2 | 2 | 2 | 1 | 1 | 2 | 4 |
| Max. | 27 | 16 | 13 | 24 | 19 | 20 | 31 | 107 | 26 | 26 | 31 | 21 | 39 | 42 |
| Spatial Factor | Change in Return Level 1 | Average Return Level % Change |
|---|---|---|
| 3-year increase in Median age of public apartments | 1.8 | 3.8% |
| 10% increase in Impervious surfaces | 1.6 | 3.3% |
| Vegetation 2 | 0.7 | 1.4% |
| Freshwater surfaces | 0.3 | 0.7% |
| Population size | 0.1 | 0.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soh, S.; Ho, S.H.; Seah, A.; Ong, J.; Richards, D.R.; Gaw, L.Y.-F.; Dickens, B.S.; Tan, K.W.; Koo, J.R.; Cook, A.R.; et al. Spatial Methods for Inferring Extremes in Dengue Outbreak Risk in Singapore. Viruses 2022, 14, 2450. https://doi.org/10.3390/v14112450
Soh S, Ho SH, Seah A, Ong J, Richards DR, Gaw LY-F, Dickens BS, Tan KW, Koo JR, Cook AR, et al. Spatial Methods for Inferring Extremes in Dengue Outbreak Risk in Singapore. Viruses. 2022; 14(11):2450. https://doi.org/10.3390/v14112450
Chicago/Turabian StyleSoh, Stacy, Soon Hoe Ho, Annabel Seah, Janet Ong, Daniel R. Richards, Leon Yan-Feng Gaw, Borame Sue Dickens, Ken Wei Tan, Joel Ruihan Koo, Alex R. Cook, and et al. 2022. "Spatial Methods for Inferring Extremes in Dengue Outbreak Risk in Singapore" Viruses 14, no. 11: 2450. https://doi.org/10.3390/v14112450
APA StyleSoh, S., Ho, S. H., Seah, A., Ong, J., Richards, D. R., Gaw, L. Y. -F., Dickens, B. S., Tan, K. W., Koo, J. R., Cook, A. R., & Lim, J. T. (2022). Spatial Methods for Inferring Extremes in Dengue Outbreak Risk in Singapore. Viruses, 14(11), 2450. https://doi.org/10.3390/v14112450