
Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections

 ,
,  and
and
Abstract
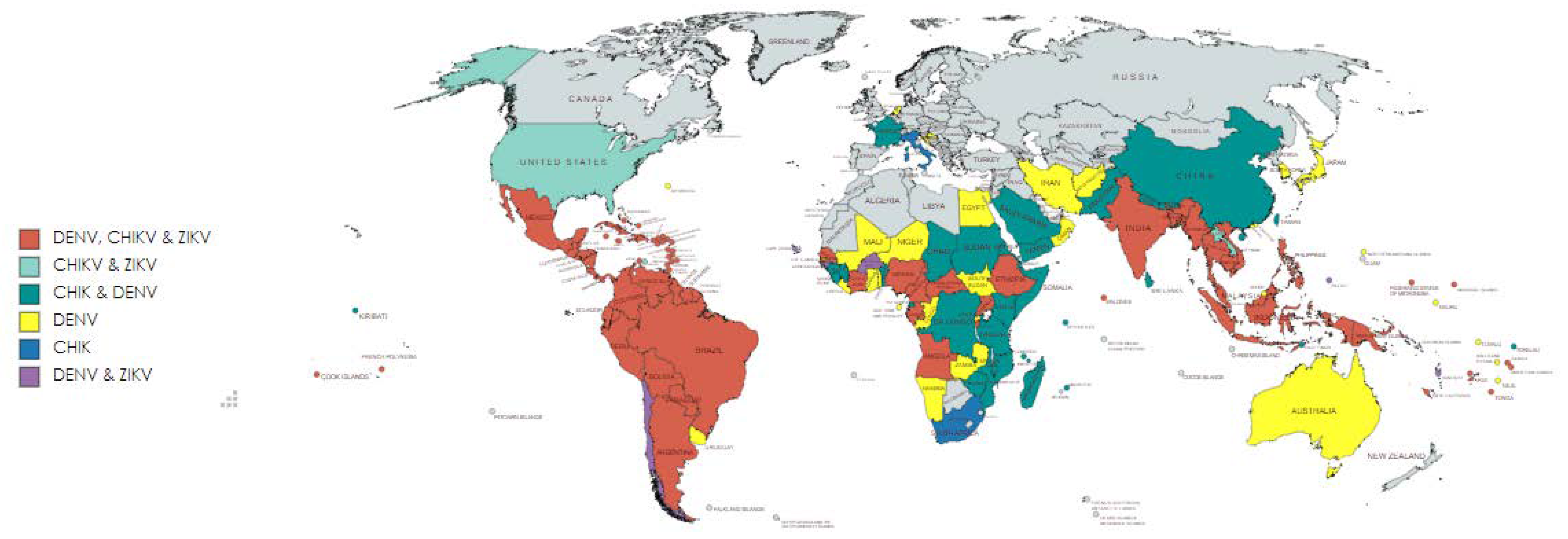
:1. Introduction
2. Epitope Identification of CHIKV, DENV and ZIKV; Progression and Knowledge Gap
3. Complex Role of T Cells in CHIKV, DENV and ZIKV
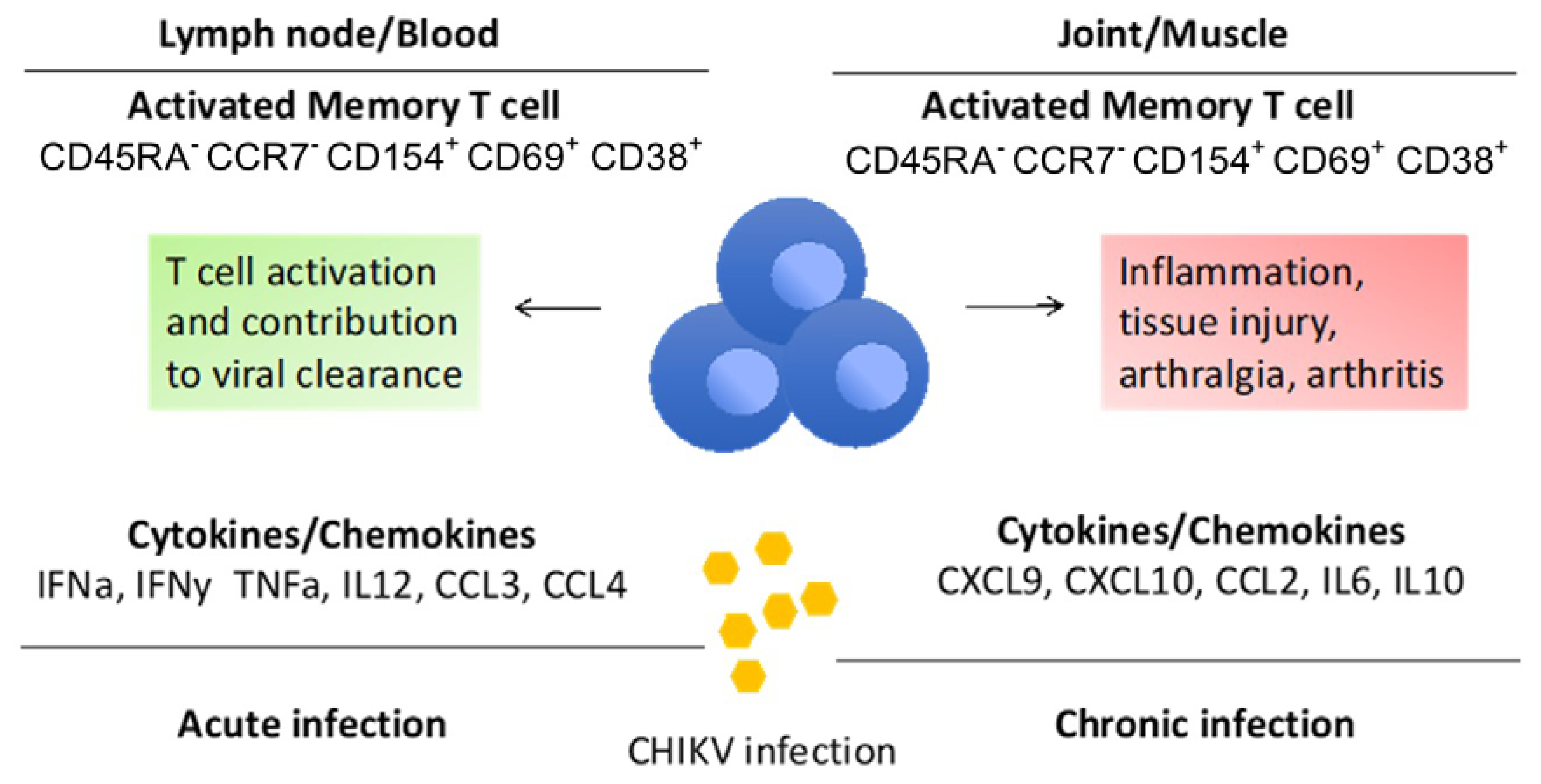
3.1. Understanding the Role of T Cells in CHIKV: Nescience and Opportunity for Research
3.2. T Cell Response in DENV Infection
3.3. Role of T Cells Response in ZIKV Infection
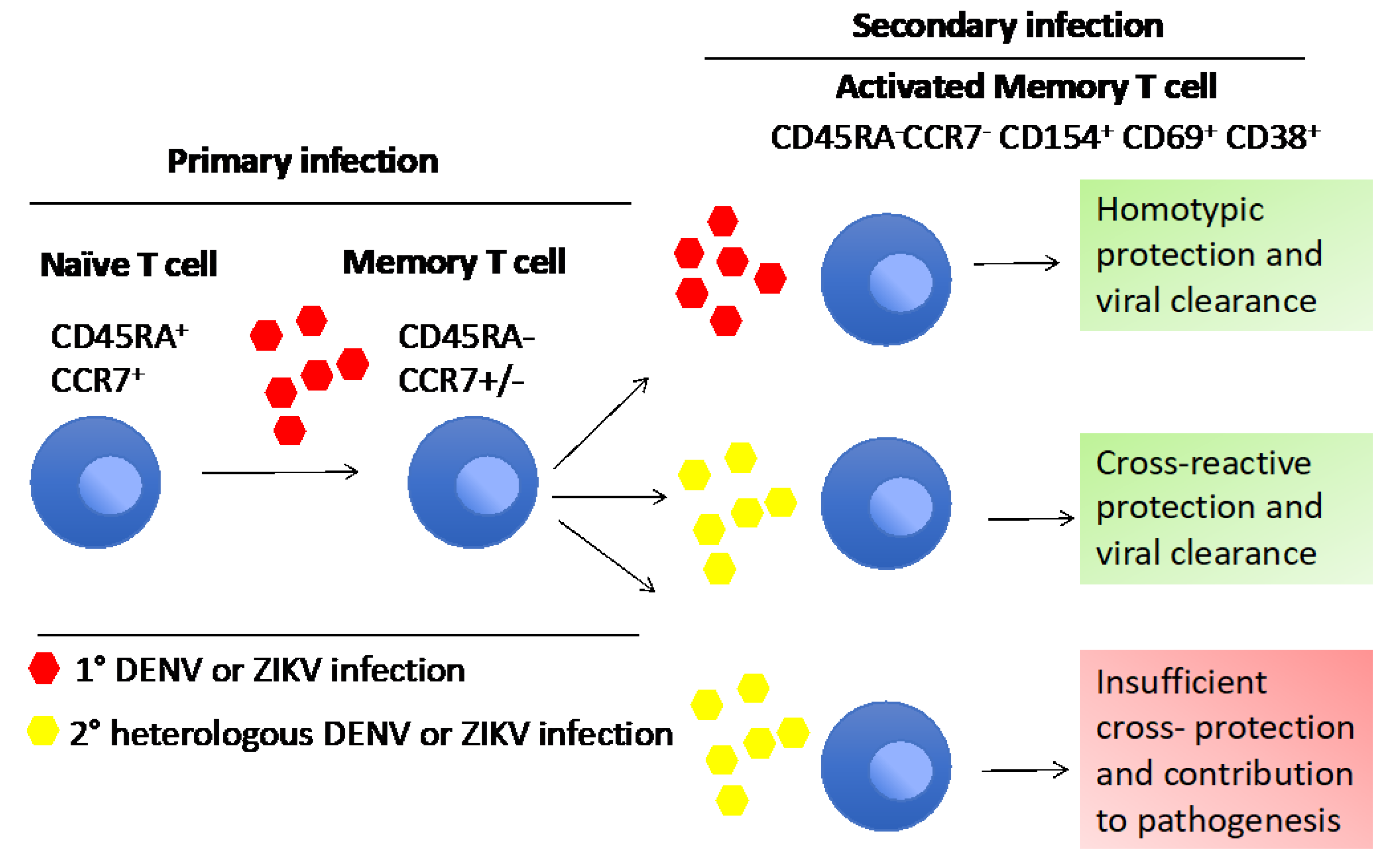
4. The Impact of Sequence Homology and Cross-Reactivity between DENV, ZIKV and CHIKV Shaping Their T Cell Responses
5. Current Understanding of T Cell Responses on Future Vaccine Development and Immune Therapy in Arboviral Infections
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilder-Smith, A.; Gubler, D.J.; Weaver, S.C.; Monath, T.P.; Heymann, D.L.; Scott, T.W. Epidemic Arboviral Diseases: Priorities for Research and Public Health. Lancet Infect. Dis. 2017, 17, e101–e106. [Google Scholar] [CrossRef] [Green Version]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 15412. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of Dengue Virus: Implications for Flavivirus Organization, Maturation, and Fusion. Cell 2002, 108, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Sirohi, D.; Kuhn, R.J. Zika Virus Structure, Maturation, and Receptors. J. Infect. Dis. 2017, 216 (Suppl. 10), S935–S944. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, V.K.; Duan, B.; Reid, S.P. Chikungunya Virus: Pathophysiology, Mechanism, and Modeling. Viruses 2017, 9, 368. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.M.O.; Tauro, L.B.; Kikuti, M.; Anjos, R.O.; Santos, V.C.; Gonçalves, T.S.F.; Paploski, I.A.D.; Moreira, P.S.S.; Nascimento, L.C.J.; Campos, G.S.; et al. Concomitant Transmission of Dengue, Chikungunya, and Zika Viruses in Brazil: Clinical and Epidemiological Findings from Surveillance for Acute Febrile Illness. Clin. Infect. Dis. 2019, 69, 1353–1359. [Google Scholar] [CrossRef]
- Rodríguez-Barraquer, I.; Salje, H.; Cummings, D.A. Dengue Pre-Vaccination Screening and Positive Predictive Values. Lancet Infect. Dis. 2019, 19, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N. Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef]
- Ferguson, N.M.; Rodríguez-Barraquer, I.; Dorigatti, I.; Mier-Y-Teran-Romero, L.; Laydon, D.J.; Cummings, D.A.T. Benefits and Risks of the Sanofi-Pasteur Dengue Vaccine: Modeling Optimal Deployment. Science 2016, 353, 1033–1036. [Google Scholar] [CrossRef] [Green Version]
- Martinez, D.R.; Metz, S.W.; Baric, R.S. Dengue Vaccines: The Promise and Pitfalls of Antibody-Mediated Protection. Cell Host Microbe 2021, 29, 13–22. [Google Scholar] [CrossRef]
- Muthumani, K.; Griffin, B.D.; Agarwal, S.; Kudchodkar, S.B.; Reuschel, E.L.; Choi, H.; Kraynyak, K.A.; Duperret, E.K.; Keaton, A.A.; Chung, C.; et al. In Vivo Protection against ZIKV Infection and Pathogenesis through Passive Antibody Transfer and Active Immunisation with a PrMEnv DNA Vaccine. NPJ Vaccines 2016, 1, 16021. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Ko, S.; Morabito, K.M.; Yang, E.S.; Pelc, R.S.; Demaso, C.R.; Castilho, L.R.; Abbink, P.; Boyd, M.; Nityanandam, R.; et al. Rapid Development of a DNA Vaccine for Zika Virus. Science 2016, 354, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelman, R.; Tacket, C.O.; Wasserman, S.S.; Bodison, S.A.; Perry, J.G.; Mangiafico, J.A. Phase II Safety and Immunogenicity Study of Live Chikungunya Virus Vaccine TSI-GSD-218. Am. J. Trop. Med. Hyg. 2000, 62, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Erasmus, J.H.; Auguste, A.J.; Kaelber, J.T.; Luo, H.; Shannan, L.; Fenton, K.; Leal, G.; Kim, D.Y.; Chiu, W.; Wang, T.; et al. A Chikungunya Fever Vaccine Utilizing an Insect-Specific Virus Platform. Nat. Med. 2017, 23, 192–199. [Google Scholar] [CrossRef]
- Tian, Y.; Grifoni, A.; Sette, A.; Weiskopf, D. Human T Cell Response to Dengue Virus Infection. Front. Immunol. 2019, 10, 2125. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, G.S.; Joshi, A.; Akhtar, N.; Kaushik, V. Immunoinformatics Designed T Cell Multi Epitope Dengue Peptide Vaccine Derived from Non Structural Proteome. Microb. Pathog. 2021, 150, 104728. [Google Scholar] [CrossRef]
- Grifoni, A.; Pham, J.; Sidney, J.; O’Rourke, P.H.; Paul, S.; Peters, B.; Martini, S.R.; de Silva, A.D.; Ricciardi, M.J.; Magnani, D.M.; et al. Prior Dengue Virus Exposure Shapes T Cell Immunity to Zika Virus in Humans. J. Virol. 2017, 91, e01469-17. [Google Scholar] [CrossRef] [Green Version]
- Delgado, F.G.; Torres, K.I.; Castellanos, J.E.; Romero-Sánchez, C.; Simon-Lorière, E.; Sakuntabhai, A.; Roth, C. Improved Immune Responses against Zika Virus after Sequential Dengue and Zika Virus Infection in Humans. Viruses 2018, 10, 480. [Google Scholar] [CrossRef] [Green Version]
- Hanajiri, R.Y.O.; SANI, G.M.; HANLEY, P.J.; SILVEIRA, C.G.; KALLAS, E.G.; KELLER, M.D.; BOLLARD, C.M. Generation of Zika Virus–Specific T Cells from Seropositive and Virus-Naïve Donors for Potential Use as an Autologous or “off-the-Shelf” Immunotherapeutic. Cytotherapy 2019, 21, 840–855. [Google Scholar] [CrossRef]
- Lim, M.Q.; Kumaran, E.A.P.; Tan, H.C.; Lye, D.C.; Leo, Y.S.; Ooi, E.E.; MacAry, P.A.; Bertoletti, A.; Rivino, L. Cross-Reactivity and Anti-Viral Function of Dengue Capsid and NS3- Specific Memory t Cells toward Zika Virus. Front. Immunol. 2018, 9, 2225. [Google Scholar] [CrossRef] [Green Version]
- Paquin-Proulx, D.; Leal, F.E.; Terrassani Silveira, C.G.; Maestri, A.; Brockmeyer, C.; Kitchen, S.M.; Cabido, V.D.; Kallas, E.G.; Nixon, D.F. T-Cell Responses in Individuals Infected with Zika Virus and in Those Vaccinated Against Dengue Virus. Pathog. Immun. 2017, 2, 274. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, M.J.; Magnani, D.M.; Grifoni, A.; Kwon, Y.C.; Gutman, M.J.; Grubaugh, N.D.; Gangavarapu, K.; Sharkey, M.; Silveira, C.G.T.; Bailey, V.K.; et al. Ontogeny of the B- and T-Cell Response in a Primary Zika Virus Infection of a Dengue-Naïve Individual during the 2016 Outbreak in Miami, FL. PLoS Negl. Trop. Dis. 2017, 11, e0006000. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Voic, H.; Dhanda, S.K.; Kidd, C.K.; Brien, J.D.; Buus, S.; Stryhn, A.; Durbin, A.P.; Whitehead, S.; Diehl, S.A.; et al. T Cell Responses Induced by Attenuated Flavivirus Vaccination Are Specific and Show Limited Cross-Reactivity with Other Flavivirus Species. J. Virol. 2020, 94, e00089-20. [Google Scholar] [CrossRef]
- Schouest, B.; Grifoni, A.; Pham, J.; Mateus, J.; Sydney, J.; Brien, J.D.; De Silva, A.D.; Balmaseda, A.; Harris, E.; Sette, A.; et al. Pre-Existing T Cell Memory against Zika Virus. J. Virol. 2021, 95, e00132-21. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.J.; Watber, P.; Santos, C.N.O.; Ribeiro, D.R.; Alves, J.C.; Fonseca, A.B.L.; Bispo, A.J.B.; Porto, R.L.S.; Bokea, K.; de Jesus, A.M.R.; et al. Strong CD4 T Cell Responses to Zika Virus Antigens in a Cohort of Dengue Virus Immune Mothers of Congenital Zika Virus Syndrome Infants. Front. Immunol. 2020, 11, 185. [Google Scholar] [CrossRef] [Green Version]
- Koblischke, M.; Stiasny, K.; Aberle, S.W.; Malafa, S.; Tschouchnikas, G.; Schwaiger, J.; Kundi, M.; Heinz, F.X.; Aberle, J.H. Structural Influence on the Dominance of Virus-Specific CD4 T Cell Epitopes in Zika Virus Infection. Front. Immunol. 2018, 9, 1196. [Google Scholar] [CrossRef]
- Khan, B.A.; Saifullah, A.L.; Lail, A.; Khan, S. Sub-Genomic Analysis of Chikungunya Virus E2 Mutations in Pakistani Isolates Potentially Modulating b-Cell & t-Cell Immune Response. Pakistan J. Med. Sci. 2020, 37, 93. [Google Scholar] [CrossRef]
- Bukhari, S.N.H.; Jain, A.; Haq, E.; Khder, M.A.; Neware, R.; Bhola, J.; Lari Najafi, M. Machine Learning-Based Ensemble Model for Zika Virus T-Cell Epitope Prediction. J. Healthc. Eng. 2021, 2021, 9591670. [Google Scholar] [CrossRef]
- Yactayo, S.; Staples, J.E.; Millot, V.; Cibrelus, L.; Ramon-Pardo, P. Epidemiology of Chikungunya in the Americas. J. Infect. Dis. 2016, 214 (Suppl. 5), S441–S445. [Google Scholar] [CrossRef] [Green Version]
- Thiberville, S.D.; Moyen, N.; Dupuis-Maguiraga, L.; Nougairede, A.; Gould, E.A.; Roques, P.; de Lamballerie, X. Chikungunya Fever: Epidemiology, Clinical Syndrome, Pathogenesis and Therapy. Antiviral Res. 2013, 99, 345–370. [Google Scholar] [CrossRef]
- Borgherini, G.; Poubeau, P.; Staikowsky, F.; Lory, M.; Moullec, N.L.; Becquart, J.P.; Wengling, C.; Michault, A.; Paganin, F. Outbreak of Chikungunya on Reunion Island: Early Clinical and Laboratory Features in 157 Adult Patients. Clin. Infect. Dis. 2007, 44, 1401–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wauquier, N.; Becquart, P.; Nkoghe, D.; Padilla, C.; Ndjoyi-Mbiguino, A.; Leroy, E.M. The Acute Phase of Chikungunya Virus Infection in Humans Is Associated with Strong Innate Immunity and T CD8 Cell Activation. J. Infect. Dis. 2011, 204, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Foo, S.S.; Sims, N.A.; Herrero, L.J.; Walsh, N.C.; Mahalingam, S. Arthritogenic Alphaviruses: New Insights into Arthritis and Bone Pathology. Trends Microbiol. 2015, 23, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.M.; Chan, Y.H.; Ng, L.F.P. Role of T Cells in Chikungunya Virus Infection and Utilizing Their Potential in Anti-Viral Immunity. Front. Immunol. 2020, 11, 287. [Google Scholar] [CrossRef] [Green Version]
- de Sousa Dias, C.N.; Gois, B.M.; Lima, V.S.; Guerra-Gomes, I.C.; Araújo, J.M.G.; de Assis Silva Gomes, J.; Araújo, D.A.M.; Medeiros, I.A.; de Lourdes Assunção Araújo de Azevedo, F.; Veras, R.C.; et al. Human CD8 T-Cell Activation in Acute and Chronic Chikungunya Infection. Immunology 2018, 155, 499–504. [Google Scholar] [CrossRef]
- Hoarau, J.-J.; Jaffar Bandjee, M.-C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent Chronic Inflammation and Infection by Chikungunya Arthritogenic Alphavirus in Spite of a Robust Host Immune Response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.N.; Ahmed, R. High Antigen Levels Are the Cause of T Cell Exhaustion during Chronic Viral Infection. Proc. Natl. Acad. Sci. USA 2009, 106, 8623–8628. [Google Scholar] [CrossRef] [Green Version]
- Petitdemange, C.; Wauquier, N.; Vieillard, V. Control of Immunopathology during Chikungunya Virus Infection. J. Allergy Clin. Immunol. 2015, 135, 846–855. [Google Scholar] [CrossRef]
- Teo, T.H.; Chan, Y.H.; Lee, W.W.L.; Lum, F.M.; Amrun, S.N.; Her, Z.; Rajarethinam, R.; Merits, A.; Rötzschke, O.; Rénia, L.; et al. Fingolimod Treatment Abrogates Chikungunya Virus-Induced Arthralgia. Sci. Transl. Med. 2017, 9, 375. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Harrison, K.; Preciado-Llanes, L.; Lopez, F.R.; Bittaye, M.; Kim, Y.C.; Flaxman, A.; Bellamy, D.; Makinson, R.; Sheridan, J.; et al. A Single Dose of ChAdOx1 Chik Vaccine Induces Neutralizing Antibodies against Four Chikungunya Virus Lineages in a Phase 1 Clinical Trial. Nat. Commun. 2021, 12, 4636. [Google Scholar] [CrossRef]
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya Virus Arthritis in Adult Wild-Type Mice. J. Virol. 2010, 84, 8021–8032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labadie, K.; Le Grand, R.; Roques, P.; Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; De Lamballerie, X.; Suhrbier, A.; et al. Chikungunya Disease in Nonhuman Primates Involves Long-Term Viral Persistence in Macrophages Find the Latest Version: Chikungunya Disease in Nonhuman Primates Involves Long-Term Viral Persistence in Macrophages. J. Clin. Investig. 2010, 120, 894–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, T.E.; Oko, L.; Montgomery, S.A.; Whitmore, A.C.; Lotstein, A.R.; Gunn, B.M.; Elmore, S.A.; Heise, M.T. A Mouse Model of Chikungunya Virus-Induced Musculoskeletal Inflammatory Disease: Evidence of Arthritis, Tenosynovitis, Myositis, and Persistence. Am. J. Pathol. 2011, 178, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Noret, M.; Herrero, L.; Rulli, N.; Rolph, M.; Smith, P.N.; Li, R.W.; Roques, P.; Gras, G.; Mahalingam, S. Interleukin 6, RANKL, and Osteoprotegerin Expression by Chikungunya Virus-Infected Human Osteoblasts. J. Infect. Dis. 2012, 206, 455–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Born, W.; Cady, C.; Jones-Carson, J.; Mukasa, A.; Lahn, M.; O’Brien, R. Immunoregulatory Functions of Γδ T Cells. Adv. Immunol. 1998, 71, 77–144. [Google Scholar] [CrossRef]
- Carding, S.R.; Egan, P.J. Γδ T Cells: Functional Plasticity and Heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Long, K.M.; Ferris, M.T.; Whitmore, A.C.; Montgomery, S.A.; Thurlow, L.R.; McGee, C.E.; Rodriguez, C.A.; Lim, J.K.; Heise, M.T. Γδ T Cells Play a Protective Role in Chikungunya Virus-Induced Disease. J. Virol. 2016, 90, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.P.; Ganu, M.; Jayawant, P.; Thanapati, S.; Ganu, A.; Tripathy, A.S. Regulatory T Cells and IL-10 as Modulators of Chikungunya Disease Outcome: A Preliminary Study. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2475–2481. [Google Scholar] [CrossRef]
- Rivino, L.; Kumaran, E.A.P.; Jovanovic, V.; Nadua, K.; Teo, E.W.; Pang, S.W.; Teo, G.H.; Gan, V.C.H.; Lye, D.C.; Leo, Y.S.; et al. Differential Targeting of Viral Components by CD4+ versus CD8+ T Lymphocytes in Dengue Virus Infection. J. Virol. 2013, 87, 2693–2706. [Google Scholar] [CrossRef] [Green Version]
- Cerny, D.; Haniffa, M.; Shin, A.; Bigliardi, P.; Tan, B.K.; Lee, B.; Poidinger, M.; Tan, E.Y.; Ginhoux, F.; Fink, K. Selective Susceptibility of Human Skin Antigen Presenting Cells to Productive Dengue Virus Infection. PLoS Pathog. 2014, 10, e1004548. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.J.L.; Grouard-Vogel, G.; Sun, W.; Mascola, J.R.; Brachtel, E.; Putvatana, R.; Louder, M.K.; Filgueira, L.; Marovich, M.A.; Wong, H.K.; et al. Human Skin Langerhans Cells Are Targets of Dengue Virus Infection. Nat. Med. 2000, 6, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Le Duyen, H.T.; Cerny, D.; Trung, D.T.; Pang, J.; Velumani, S.; Toh, Y.X.; Qui, P.T.; Van Hao, N.; Simmons, C.; Haniffa, M.; et al. Skin Dendritic Cell and T Cell Activation Associated with Dengue Shock Syndrome. Sci. Rep. 2017, 7, 14224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivino, L.; Kumaran, E.A.; Thein, T.L.; Too, C.T.; Gan, V.C.H.; Hanson, B.J.; Wilder-Smith, A.; Bertoletti, A.; Gascoigne, N.R.J.; Lye, D.C.; et al. Virus-Specific T Lymphocytes Home to the Skin during Natural Dengue Infection. Sci. Transl. Med. 2015, 7, 278ra35. [Google Scholar] [CrossRef] [PubMed]
- Rivino, L.; Lim, M.Q. CD4+ and CD8+ T-Cell Immunity to Dengue—Lessons for the Study of Zika Virus. Immunology 2017, 150, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, S.N.; Gebhardt, T.; Carbone, F.R.; Heath, W.R. Memory T Cell Subsets, Migration Patterns, and Tissue Residence. Annu. Rev. Immunol. 2013, 31, 137–161. [Google Scholar] [CrossRef]
- Rattanamahaphoom, J.; Leaungwutiwong, P.; Limkittikul, K.; Kosoltanapiwat, N.; Srikaitkhachorn, A. Activation of Dengue Virus-Specific T Cells Modulates Vascular Endothelial Growth Factor Receptor 2 Expression. Asian Pacific J. Allergy Immunol. 2018, 35, 171–178. [Google Scholar] [CrossRef]
- Mongkolsapaya, J.; Wanwisa, D.; Xu, X.-N.; Vasanawathana, S.; Angthawornchaikul, N.T.; Chairunsri, A.; Sawasdivorn, O.; Duangchinda, I.T.; Dong, T.; Rowland-Jones, S.; et al. Original Antigenic Sin and Apoptosis in the Pathogenesis of Dengue Hemorrhagic Fever. Nat. Med. 2003, 9, 921–927. [Google Scholar] [CrossRef]
- Halstead, S.B.; Rojanasuphot, S.; Sangkawibha, N. Original Antigenic Sin in Dengue. Am. J. Trop. Med. Hyg. 1983, 32, 154–156. [Google Scholar] [CrossRef]
- Kurane, I.; Meager, A.; Ennis, F.A. Dengue Virus-Specific Human t Cell Clones. Serotype Crossreactive Proliferation, Interferon γ Production, and Cytotoxic Activity. J. Exp. Med. 1989, 170, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Friberg, H.; Bashyam, H.; Toyosaki-Maeda, T.; Potts, J.A.; Greenough, T.; Kalayanarooj, S.; Gibbons, R.V.; Nisalak, A.; Srikiatkhachorn, A.; Green, S.; et al. Cross-Reactivity and Expansion of Dengue-Specific T Cells during Acute Primary and Secondary Infections in Humans. Sci. Rep. 2011, 1, 51. [Google Scholar] [CrossRef]
- Wrammert, J.; Onlamoon, N.; Akondy, R.S.; Perng, G.C.; Polsrila, K.; Chandele, A.; Kwissa, M.; Pulendran, B.; Wilson, P.C.; Wittawatmongkol, O.; et al. Rapid and Massive Virus-Specific Plasmablast Responses during Acute Dengue Virus Infection in Humans. J. Virol. 2012, 86, 2911–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyamvada, L. B Cell Responses during Secondary Dengue Virus Infection Are. J. Virol. 2016, 90, 5574–5585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Hadinoto, V.; Appanna, R.; Joensson, K.; Toh, Y.X.; Balakrishnan, T.; Ong, S.H.; Warter, L.; Leo, Y.S.; Wang, C.-I.; et al. Plasmablasts Generated during Repeated Dengue Infection Are Virus Glycoprotein–Specific and Bind to Multiple Virus Serotypes. J. Immunol. 2012, 189, 5877–5885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskopf, D.; Angelo, M.A.; De Azeredo, E.L.; Sidney, J.; Greenbaum, J.A.; Fernando, A.N.; Broadwater, A.; Kolla, R.V.; De Silva, A.D.; De Silva, A.M.; et al. Comprehensive Analysis of Dengue Virus-Specific Responses Supports an HLA-Linked Protective Role for CD8+ T Cells. Proc. Natl. Acad. Sci. USA 2013, 110, E2046–E2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskopf, D.; Angelo, M.A.; Grifoni, A.; O’Rourke, P.H.; Sidney, J.; Paul, S.; De Silva, A.D.; Phillips, E.; Mallal, S.; Premawansa, S.; et al. HLA-DRB1 Alleles Are Associated with Different Magnitudes of Dengue Virus-Specific CD4+ T-Cell Responses. J. Infect. Dis. 2016, 214, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- Yauch, L.E.; Zellweger, R.M.; Kotturi, M.F.; Qutubuddin, A.; Sidney, J.; Peters, B.; Prestwood, T.R.; Sette, A.; Shresta, S. A Protective Role for Dengue Virus-Specific CD8 + T Cells. J. Immunol. 2009, 182, 4865–4873. [Google Scholar] [CrossRef] [Green Version]
- Zellweger, R.M.; Eddy, W.E.; Tang, W.W.; Miller, R.; Shresta, S. CD8 + T Cells Prevent Antigen-Induced Antibody-Dependent Enhancement of Dengue Disease in Mice. J. Immunol. 2014, 193, 4117–4124. [Google Scholar] [CrossRef] [Green Version]
- Zellweger, R.M.; Tang, W.W.; Eddy, W.E.; King, K.; Sanchez, M.C.; Shresta, S. CD8 + T Cells Can Mediate Short-Term Protection against Heterotypic Dengue Virus Reinfection in Mice. J. Virol. 2015, 89, 6494–6505. [Google Scholar] [CrossRef] [Green Version]
- Elong Ngono, A.; Shresta, S. Immune Response to Dengue and Zika. Annu. Rev. Immunol. 2018, 36, 279–308. [Google Scholar] [CrossRef] [Green Version]
- de Alwis, R.; Bangs, D.J.; Angelo, M.A.; Cerpas, C.; Fernando, A.; Sidney, J.; Peters, B.; Gresh, L.; Balmaseda, A.; de Silva, A.D.; et al. Immunodominant Dengue Virus-Specific CD8 + T Cell Responses Are Associated with a Memory PD-1 + Phenotype. J. Virol. 2016, 90, 4771–4779. [Google Scholar] [CrossRef] [Green Version]
- Odorizzi, P.M.; Pauken, K.E.; Paley, M.A.; Sharpe, A.; John Wherry, E. Genetic Absence of PD-1 Promotes Accumulation of Terminally Differentiated Exhausted CD8+ T Cells. J. Exp. Med. 2015, 212, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Chandele, A.; Sewatanon, J.; Gunisetty, S.; Singla, M.; Onlamoon, N.; Akondy, R.S.; Kissick, H.T.; Nayak, K.; Reddy, E.S.; Kalam, H.; et al. Characterization of Human CD8 T Cell Responses in Dengue Virus-Infected Patients from India. J. Virol. 2016, 90, 11259–11278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Babor, M.; Lane, J.; Seumois, G.; Liang, S.; Suraj Goonawardhana, N.D.; De Silva, A.D.; Phillips, E.J.; Mallal, S.A.; da Silva Antunes, R.; et al. Dengue-Specific CD8+ T Cell Subsets Display Specialized Transcriptomic and TCR Profiles. J. Clin. Investig. 2019, 129, 1727–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Wang, H.; Yu, Q.; Liu, J.J.; Chen, S.; Zhao, Z. Protective Role of Follicular CXCR5+CD8+ T Cells against Dengue Virus 2 Infection. Int. J. Infect. Dis. 2019, 83, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves Pereira, M.H.; Figueiredo, M.M.; Queiroz, C.P.; Magalhães, T.V.B.; Mafra, A.; Diniz, L.M.O.; da Costa, Ú.L.; Gollob, K.J.; do Valle Antonelli, L.R.; da Costa Santiago, H. T Cells Producing Multiple Combinations of IFNγ, TNF and IL10 Are Associated with Mild Forms of Dengue Infection. Immunology 2020, 160, 90–102. [Google Scholar] [CrossRef]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding Roles for CD4 + T Cells in Immunity to Viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef]
- Rivino, L. Understanding the Human T Cell Response to Dengue Virus; Springer: Singapore, 2018; Volume 1062, pp. 265–276. [Google Scholar] [CrossRef]
- Gagnon, S.J.; Ennis, F.A.; Rothman, A.L. Bystander Target Cell Lysis and Cytokine Production by Dengue Virus-Specific Human CD4+ Cytotoxic T-Lymphocyte Clones. J. Virol. 1999, 73, 3623–3629. [Google Scholar] [CrossRef] [Green Version]
- St. John, A.L.; Rathore, A.P.S. Adaptive Immune Responses to Primary and Secondary Dengue Virus Infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef]
- Simon-Lorière, E.; Duong, V.; Tawfik, A.; Ung, S.; Ly, S.; Casadémont, I.; Prot, M.; Courtejoie, N.; Bleakley, K.; Buchy, P.; et al. Increased Adaptive Immune Responses and Proper Feedback Regulation Protect against Clinical Dengue. Sci. Transl. Med. 2017, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Weiskopf, D.; Bangs, D.J.; Sidney, J.; Kolla, R.V.; De Silva, A.D.; De Silva, A.M.; Crotty, S.; Peters, B.; Sette, A. Dengue Virus Infection Elicits Highly Polarized CX3CR1+ Cytotoxic CD4+ T Cells Associated with Protective Immunity. Proc. Natl. Acad. Sci. USA 2015, 112, E4256–E4263. [Google Scholar] [CrossRef] [Green Version]
- Lühn, K.; Simmons, C.P.; Moran, E.; Dung, N.T.P.; Chau, T.N.B.; Quyen, N.T.H.; Thao, L.T.T.; Van Ngoc, T.; Nguyen, M.D.; Wills, B.; et al. Increased Frequencies of CD4+CD25high Regulatory T Cells in Acute Dengue Infection. J. Exp. Med. 2007, 204, 979–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaratne, H.E.; Wijeratne, D.; Fernando, S.; Kamaladasa, A.; Gomes, L.; Wijewickrama, A.; Ogg, G.S.; Malavige, G.N. Regulatory T-Cells in Acute Dengue Viral Infection. Immunology 2018, 154, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saron, W.A.A.; Rathore, A.P.S.; Ting, L.; Ooi, E.E.; Low, J.; Abraham, S.N.; St. John, A.L. Flavivirus Serocomplex Cross-Reactive Immunity Is Protective by Activating Heterologous Memory CD4 T Cells. Sci. Adv. 2018, 4, eaar4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haltaufderhyde, K.; Srikiatkhachorn, A.; Green, S.; Macareo, L.; Park, S.; Kalayanarooj, S.; Rothman, A.L.; Mathew, A. Activation of Peripheral T Follicular Helper Cells during Acute Dengue Virus Infection. J. Infect. Dis. 2018, 218, 1675–1685. [Google Scholar] [CrossRef]
- Elgueta, R.; Benson, M.J.; De Vries, V.C.; Noelle, R.J.; Noelle, R.J. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef] [Green Version]
- Noelle, R.J.; Roy, M.; Shepherd, D.M.; Stamenkovic, I.; Ledbetter, J.A.; Aruffo, A. A 39-KDa Protein on Activated Helper T Cells Binds CD40 and Transduces the Signal for Cognate Activation of B Cells. Proc. Natl. Acad. Sci. USA 1992, 89, 6550–6554. [Google Scholar] [CrossRef] [Green Version]
- Chase, A.J.; Medina, F.A.; Muñoz-Jordán, J.L. Impairment of CD4+ T Cell Polarization by Dengue Virus-Infected Dendritic Cells. J. Infect. Dis. 2011, 203, 1763–1774. [Google Scholar] [CrossRef]
- Sprokholt, J.K.; Kaptein, T.M.; van Hamme, J.L.; Overmars, R.J.; Gringhuis, S.I.; Geijtenbeek, T.B.H. RIG-I-like Receptor Activation by Dengue Virus Drives Follicular T Helper Cell Formation and Antibody Production. PLoS Pathog. 2017, 13, e1006738. [Google Scholar] [CrossRef] [Green Version]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Musso, D.; Roche, C.; Robin, E.; Nhan, T.; Teissier, A.; Cao-Lormeau, V.M. Potential Sexual Transmission of Zika Virus. EMBO Rep. 2015, 21, 359–361. [Google Scholar] [CrossRef]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonça, M.C.L.; de Oliveira, L.; et al. Detection and Sequencing of Zika Virus from Amniotic Fluid of Fetuses with Microcephaly in Brazil: A Case Study. Lancet Infect. Dis. 2016, 16, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Oliveira Melo, A.S.; Malinger, G.; Ximenes, R.; Szejnfeld, P.O.; Alves Sampaio, S.; Bispo De Filippis, A.M. Zika Virus Intrauterine Infection Causes Fetal Brain Abnormality and Microcephaly: Tip of the Iceberg? Ultrasound Obstet. Gynecol. 2016, 47, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastãre, S.; Valour, F.; Baudouin, L.; Mallet, H.P.; Musso, D.; Ghawche, F. Zika Virus Infection Complicated by Guillain-Barré Syndrome €”case Report, French Polynesia, December 2013. Eurosurveillance 2014, 19, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Dudley, D.M.; Aliota, M.T.; Mohr, E.L.; Weiler, A.M.; Lehrer-Brey, G.; Weisgrau, K.L.; Mohns, M.S.; Breitbach, M.E.; Rasheed, M.N.; Newman, C.M.; et al. A Rhesus Macaque Model of Asian-Lineage Zika Virus Infection. Nat. Commun. 2016, 7, 12204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, A.; Ponia, S.S.; Tripathi, S.; Balasubramaniam, V.; Miorin, L.; Sourisseau, M.; Schwarz, M.C.; Sánchez-Seco, M.P.; Evans, M.J.; Best, S.M.; et al. Zika Virus Targets Human STAT2 to Inhibit Type i Interferon Signaling. Cell Host Microbe 2016, 19, 882–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, V.; Yuen, K.; Chan, J.F.; Chan, C. Selective Activation of Type II Interferon Signaling by Zika Virus NS5 Protein. J. Virol. 2017, 91, e00163-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, J.R.; Quicke, K.M.; Maddur, M.S.; O’Neal, J.T.; McDonald, C.E.; Fedorova, N.B.; Puri, V.; Shabman, R.S.; Pulendran, B.; Suthar, M.S. Zika Virus Antagonizes Type I Interferon Responses during Infection of Human Dendritic Cells. PLoS Pathog. 2017, 13, e1006164. [Google Scholar] [CrossRef]
- Dowall, S.D.; Graham, V.A.; Rayner, E.; Hunter, L.; Atkinson, B.; Pearson, G.; Dennis, M.; Hewson, R. Lineage-Dependent Differences in the Disease Progression of Zika Virus Infection in Type-I Interferon Receptor Knockout (A129) Mice. PLoS Negl. Trop. Dis. 2017, 11, e0005704. [Google Scholar] [CrossRef]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe 2016, 19, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Grifoni, A.; Costa-Ramos, P.; Pham, J.; Tian, Y.; Rosales, S.L.; Seumois, G.; Sidney, J.; de Silva, A.D.; Premkumar, L.; Collins, M.H.; et al. Cutting Edge: Transcriptional Profiling Reveals Multifunctional and Cytotoxic Antiviral Responses of Zika Virus–Specific CD8 + T Cells. J. Immunol. 2018, 201, 3487–3491. [Google Scholar] [CrossRef] [Green Version]
- Tappe, D.; Pérez-Girón, J.V.; Zammarchi, L.; Rissland, J.; Ferreira, D.F.; Jaenisch, T.; Gómez-Medina, S.; Günther, S.; Bartoloni, A.; Muñoz-Fontela, C.; et al. Cytokine Kinetics of Zika Virus-Infected Patients from Acute to Reconvalescent Phase. Med. Microbiol. Immunol. 2016, 205, 269–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elong Ngono, A.; Vizcarra, E.A.; Tang, W.W.; Sheets, N.; Joo, Y.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Mapping and Role of the CD8+ T Cell Response During Primary Zika Virus Infection in Mice. Cell Host Microbe 2017, 21, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Li, S.; Zhang, Y.; Han, X.; Jia, B.; Liu, H.; Liu, D.; Tan, S.; Wang, Q.; Bi, Y.; et al. CD8+ T Cell Immune Response in Immunocompetent Mice during Zika Virus Infection. J. Virol. 2017, 91, e00900-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardy, R.D.; Rajah, M.M.; Condotta, S.A.; Taylor, N.G.; Sagan, S.M.; Richer, M.J. Analysis of the T Cell Response to Zika Virus and Identification of a Novel CD8+T Cell Epitope in Immunocompetent Mice. PLoS Pathog. 2017, 13, e1006184. [Google Scholar] [CrossRef] [Green Version]
- Larocca, R.A.; Abbink, P.; Peron, J.P.S.; Zanotto, P.M.D.A.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’ang’a, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine Protection against Zika Virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef]
- Chahal, J.S.; Fang, T.; Woodham, A.W.; Khan, O.F.; Ling, J.; Anderson, D.G.; Ploegh, H.L. An RNA Nanoparticle Vaccine against Zika Virus Elicits Antibody and CD8+ T Cell Responses in a Mouse Model. Sci. Rep. 2017, 7, 252. [Google Scholar] [CrossRef]
- Wen, J.; Tang, W.W.; Sheets, N.; Ellison, J.; Sette, A.; Kim, K.; Shresta, S. Identification of Zika Virus Epitopes Reveals Immunodominant and Protective Roles for Dengue Virus Cross-Reactive CD8+ T Cells. Nat. Microbiol. 2017, 2, 17036. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Saucedo-Cuevas, L.; Regla-Nava, J.A.; Chai, G.; Sheets, N.; Tang, W.; Terskikh, A.V.; Shresta, S.; Gleeson, J.G. Zika Virus Infects Neural Progenitors in the Adult Mouse Brain and Alters Proliferation. Cell Stem Cell 2016, 19, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Jurado, K.A.; Yockey, L.J.; Wong, P.W.; Lee, S.; Huttner, A.J.; Iwasaki, A. Antiviral CD8 T Cells Induce Zika-Virus-Associated Paralysis in Mice. Nat. Microbiol. 2018, 3, 141–147. [Google Scholar] [CrossRef]
- Mathew, A.; Rothman, A.L. Understanding the Contribution of Cellular Immunity to Dengue Disease Pathogenesis. Immunol. Rev. 2008, 225, 300–313. [Google Scholar] [CrossRef]
- McElroy, A.K.; Akondy, R.S.; Davis, C.W.; Ellebedy, A.H.; Mehta, A.K.; Kraft, C.S.; Lyon, G.M.; Ribner, B.S.; Varkey, J.; Sidney, J.; et al. Human Ebola Virus Infection Results in Substantial Immune Activation. Proc. Natl. Acad. Sci. USA 2015, 112, 4719–4724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimini, E.; Castilletti, C.; Sacchi, A.; Casetti, R.; Bordoni, V.; Romanelli, A.; Turchi, F.; Martini, F.; Tumino, N.; Nicastri, E.; et al. Human Zika Infection Induces a Reduction of IFN-γ Producing CD4 T-Cells and a Parallel Expansion of Effector Vδ2 T-Cells. Sci. Rep. 2017, 7, 6313. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.G.O.; Kitoko, J.Z.; Ferreira, F.M.; Suzart, V.G.; Papa, M.P.; Coelho, S.V.A.; Cavazzoni, C.B.; Paula-Neto, H.A.; Olsen, P.C.; Iwasaki, A.; et al. Critical Role of CD4+ T Cells and IFNγ Signaling in Antibody-Mediated Resistance to Zika Virus Infection. Nat. Commun. 2018, 9, 3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T. Role of Γδ T Cells in West Nile Virus-Induced Encephalitis: Friend or Foe? J. Neuroimmunol. 2011, 240–241, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poccia, F.; Agrati, C.; Castilletti, C.; Bordi, L.; Gioia, C.; Horejsh, D.; Ippolito, G.; Chan, P.K.S.; Hui, D.S.C.; Sung, J.J.Y.; et al. Anti–Severe Acute Respiratory Syndrome Coronavirus Immune Responses: The Role Played by Vγ9Vδ2 T Cells. J. Infect. Dis. 2006, 193, 1244–1249. [Google Scholar] [CrossRef]
- Azeredo, E.L.; Zagne, S.M.O.; Alvarenga, A.R.; Nogueira, R.M.R.; Kubelka, C.F.; De Oliveira-Pinto, L.M. Activated Peripheral Lymphocytes with Increased Expression of Cell Adhesion Molecules and Cytotoxic Markers Are Associated with Dengue Fever Disease. Mem. Inst. Oswaldo Cruz 2006, 101, 437–449. [Google Scholar] [CrossRef] [Green Version]
- Moulin, E.; Selby, K.; Cherpillod, P.; Kaiser, L.; Boillat-Blanco, N. Simultaneous Outbreaks of Dengue, Chikungunya and Zika Virus Infections: Diagnosis Challenge in a Returning Traveller with Nonspecific Febrile Illness. New Microbes New Infect. 2016, 11, 6–7. [Google Scholar] [CrossRef] [Green Version]
- Andrade, D.V.; Harris, E. Recent Advances in Understanding the Adaptive Immune Response to Zika Virus and the Effect of Previous Flavivirus Exposure. Virus Res. 2018, 254, 27–33. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue Virus Sero-Cross-Reactivity Drives Antibody-Dependent Enhancement of Infection with Zika Virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- Stettler, K.; Beltramello, M.; Espinosa, D.A.; Graham, V.; Cassotta, A.; Bianchi, S.; Vanzetta, F.; Minola, A.; Jaconi, S.; Mele, F.; et al. Specificity, Cross-Reactivity, and Function of Antibodies Elicited by Zika Virus Infection. Science 2016, 353, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Swanstrom, J.A.; Plante, J.A.; Plante, K.S.; Young, E.F.; Mcgowan, E.; Gallichotte, E.N.; Widman, D.G.; Heise, M.T. Isolated from Dengue Patients Are Protective against Zika Virus. Am. Soc. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human Antibody Responses after Dengue Virus Infection Are Highly Cross-Reactive to Zika Virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyamvada, L.; Hudson, W.; Ahmed, R.; Wrammert, J. Humoral Cross-Reactivity between Zika and Dengue Viruses: Implications for Protection and Pathology. Emerg. Microbes Infect. 2017, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanha, P.M.S.; Nascimento, E.J.M.; Braga, C.; Cordeiro, M.T.; De Carvalho, O.V.; De Mendonça, L.R.; Azevedo, E.A.N.; França, R.F.O.; Dhalia, R.; Marques, E.T.A. Dengue Virus-Specific Antibodies Enhance Brazilian Zika Virus Infection. J. Infect. Dis. 2017, 215, 781–785. [Google Scholar] [CrossRef] [Green Version]
- Paul, L.M.; Carlin, E.R.; Jenkins, M.M.; Tan, A.L.; Barcellona, C.M.; Nicholson, C.O.; Michael, S.F.; Isern, S. Dengue Virus Antibodies Enhance Zika Virus Infection. Clin. Transl. Immunol. 2016, 5, e117. [Google Scholar] [CrossRef]
- Halstead, S.B. Biologic Evidence Required for Zika Disease Enhancement by Dengue Antibodies. Emerg. Infect. Dis. 2017, 23, 569–573. [Google Scholar] [CrossRef]
- Reynolds, C.J.; Suleyman, O.M.; Ortega-Prieto, A.M.; Skelton, J.K.; Bonnesoeur, P.; Blohm, A.; Carregaro, V.; Silva, J.S.; James, E.A.; Maillère, B.; et al. T Cell Immunity to Zika Virus Targets Immunodominant Epitopes That Show Cross-Reactivity with Other Flaviviruses. Sci. Rep. 2018, 8, 672. [Google Scholar] [CrossRef]
- Collins, M.; De Silva, A. Host Response: Cross-Fit T Cells Battle Zika Virus. Nat. Microbiol. 2017, 2, 17082. [Google Scholar] [CrossRef]
- Badolato-Corrêa, J.; Sánchez-Arcila, J.C.; Alves de Souza, T.M.; Santos Barbosa, L.; Conrado Guerra Nunes, P.; da Rocha Queiroz Lima, M.; Gandini, M.; Bispo de Filippis, A.M.; Venâncio da Cunha, R.; Leal de Azeredo, E.; et al. Human T Cell Responses to Dengue and Zika Virus Infection Compared to Dengue/Zika Coinfection. Immun. Inflamm. Dis. 2018, 6, 194–206. [Google Scholar] [CrossRef]
- Herrera, B.B.; Tsai, W.-Y.; Chang, C.; Hamel, D.J.; Wang, W.-K.; Lu, Y.; Mboup, S.; Kanki, P.J. Sustained Specific and Cross-Reactive T Cell Responses to Zika and Dengue Virus NS3 in West Africa. J. Virol. 2018, 92, e01992-17. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Guzmán, E.X.; Pantoja, P.; Serrano-Collazo, C.; Hassert, M.A.; Ortiz-Rosa, A.; Rodríguez, I.V.; Giavedoni, L.; Hodara, V.; Parodi, L.; Cruz, L.; et al. Time Elapsed between Zika and Dengue Virus Infections Affects Antibody and T Cell Responses. Nat. Commun. 2019, 10, 4316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, F.; Llau, A.; Gutierrez, G.; Bezerra, H.; Coelho, G.; Ault, S.; Barbiratto, S.B.; de Resende, M.C.; Cerezo, L.; Kleber, G.L.; et al. The Decline of Dengue in the Americas in 2017: Discussion of Multiple Hypotheses. Trop. Med. Int. Health 2019, 24, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, G.S.; Kikuti, M.; Tauro, L.B.; Nascimento, L.C.J.; Cardoso, C.W.; Campos, G.S.; Ko, A.I.; Weaver, S.C.; Reis, M.G.; Kitron, U.; et al. Does Immunity after Zika Virus Infection Cross-Protect against Dengue? Lancet Glob. Health 2018, 6, e140–e141. [Google Scholar] [CrossRef] [Green Version]
- Katzelnick, L.C.; Harris, E. The Use of Longitudinal Cohorts for Studies of Dengue Viral Pathogenesis and Protection. Curr. Opin. Virol. 2018, 29, 51–61. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Narvaez, C.; Arguello, S.; Mercado, B.L.; Collado, D.; Ampie, O.; Elizondo, D.; Miranda, T.; Carillo, F.B.; Mercado, J.C.; et al. Zika Virus Infection Enhances Future Risk of Severe Dengue Disease. Science 2020, 369, 1123–1128. [Google Scholar] [CrossRef]
- Serrano-Collazo, C.; Pérez-Guzmán, E.X.; Pantoja, P.; Hassert, M.A.; Rodríguez, I.V.; Giavedoniid, L.; Hodara, V.; Parodi, L.; Cruz, L.; Arana, T.; et al. Effective Control of Early Zika Virus Replication by Dengue Immunity Is Associated to the Length of Time between the 2 Infections but Not Mediated by Antibodies. PLoS Negl. Trop. Dis. 2020, 14, e0008285. [Google Scholar] [CrossRef]
- Masel, J.; McCracken, M.K.; Gleeson, T.; Morrison, B.; Rutherford, G.; Imrie, A.; Jarman, R.G.; Koren, M.; Pollett, S. Does Prior Dengue Virus Exposure Worsen Clinical Outcomes of Zika Virus Infection? A Systematic Review, Pooled Analysis and Lessons Learned. PLoS Negl. Trop. Dis. 2019, 13, e0007060. [Google Scholar] [CrossRef]
- Culshaw, A.; Ladell, K.; Gras, S.; McLaren, J.E.; Miners, K.L.; Farenc, C.; Van Den Heuvel, H.; Gostick, E.; Dejnirattisai, W.; Wangteeraprasert, A.; et al. Germline Bias Dictates Cross-Serotype Reactivity in a Common Dengue-Virus-Specific CD8 + T Cell Response. Nat. Immunol. 2017, 18, 1228–1237. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-Dependent Enhancement of Severe Dengue Disease in Humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, R.V.; Kalanarooj, S.; Jarman, R.G.; Nisalak, A.; Vaughn, D.W.; Endy, T.P.; Mammen, M.P.; Srikiatkhachorn, A. Analysis of Repeat Hospital Admissions for Dengue to Estimate the Frequency of Third or Fourth Dengue Infections Resulting in Admissions and Dengue Hemorrhagic Fever, and Serotype Sequences. Am. J. Trop. Med. Hyg. 2007, 77, 910–913. [Google Scholar] [CrossRef] [Green Version]
- Arredondo-García, J.L.; Hadinegoro, S.R.; Reynales, H.; Chua, M.N.; Rivera Medina, D.M.; Chotpitayasunondh, T.; Tran, N.H.; Deseda, C.C.; Wirawan, D.N.; Cortés Supelano, M.; et al. Four-Year Safety Follow-up of the Tetravalent Dengue Vaccine Efficacy Randomized Controlled Trials in Asia and Latin America. Clin. Microbiol. Infect. 2018, 24, 755–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Meng, Y.; Halloran, M.E.; Longini, I.M. Dependency of Vaccine Efficacy on Preexposure and Age: A Closer Look at a Tetravalent Dengue Vaccine. Clin. Infect. Dis. 2018, 66, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Sette, A. T-Cell Immunity to Infection with Dengue Virus in Humans. Front. Immunol. 2014, 5, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, B.; Barrere, B.; Malinowski, C.; Saville, M.; Teyssou, R.; Lang, J. From Research to Phase III: Preclinical, Industrial and Clinical Development of the Sanofi Pasteur Tetravalent Dengue Vaccine. Vaccine 2011, 29, 7229–7241. [Google Scholar] [CrossRef]
- Graham, N.; Eisenhauer, P.; Diehl, S.A.; Pierce, K.K.; Whitehead, S.S.; Durbin, A.P.; Kirkpatrick, B.D.; Sette, A.; Weiskopf, D.; Boyson, J.E.; et al. Rapid Induction and Maintenance of Virus-Specific CD8+ TEMRA and CD4+ TEM Cells Following Protective Vaccination Against Dengue Virus Challenge in Humans. Front. Immunol. 2020, 11, 479. [Google Scholar] [CrossRef] [Green Version]
- Angelo, M.A.; Grifoni, A.; O’Rourke, P.H.; Sidney, J.; Paul, S.; Peters, B.; de Silva, A.D.; Phillips, E.; Mallal, S.; Deihl, S.A.; et al. Human CD4+ T Cell Responses to an Attenuated Tetravalent Dengue Vaccine Parallel Those Induced by Natural Infection in Magnitude, HLA Restriction, and Antigen Specificity. J. Virol. 2017, 91, e02147-16. [Google Scholar] [CrossRef] [Green Version]
- Weiskopf, D.; Angelo, M.A.; Bangs, D.J.; Sidney, J.; Paul, S.; Peters, B.; de Silva, A.D.; Lindow, J.C.; Diehl, S.A.; Whitehead, S.; et al. The Human CD8 + T Cell Responses Induced by a Live Attenuated Tetravalent Dengue Vaccine Are Directed against Highly Conserved Epitopes. J. Virol. 2015, 89, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.V.J.; Lopes, T.R.R.; de Oliveira-Filho, E.F.; Oliveira, R.A.S.; Durães-Carvalho, R.; Gil, L.H.V.G. Current Status, Challenges and Perspectives in the Development of Vaccines against Yellow Fever, Dengue, Zika and Chikungunya Viruses. Acta Trop. 2018, 182, 257–263. [Google Scholar] [CrossRef]
- Richner, J.M.; Diamond, M.S. Zika Virus Vaccines: Immune Response, Current Status, and Future Challenges. Curr. Opin. Immunol. 2018, 53, 130–136. [Google Scholar] [CrossRef]
- Schwameis, M.; Buchtele, N.; Wadowski, P.P.; Schoergenhofer, C.; Jilma, B. Chikungunya Vaccines in Development. Hum. Vaccines Immunother. 2016, 12, 716–731. [Google Scholar] [CrossRef]
- Pierson, T.C.; Graham, B.S. Zika Virus: Immunity and Vaccine Development. Cell 2016, 167, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Lima, N.S.; Rolland, M.; Modjarrad, K.; Trautmann, L. T Cell Immunity and Zika Virus Vaccine Development. Trends Immunol. 2017, 38, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Ngono, A.E.; Shresta, S. Cross-Reactive T Cell Immunity to Dengue and Zika Viruses: New Insights into Vaccine Development. Front. Immunol. 2019, 10, 1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, K.S.; Lant, S.; Goodwin, L.; Grifoni, A.; Weiskopf, D.; Turtle, L. Two Is Better Than One: Evidence for T-Cell Cross-Protection Between Dengue and Zika and Implications on Vaccine Design. Front. Immunol. 2020, 11, 517. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | T Cell Type/Phenotype | Disease Condition | Markers | Gene Regulation | Function | References |
|---|---|---|---|---|---|---|
| CHIKV | CD8+ T cells | Acute | Upregulation | T cell activation, destroy virus infected cells | [32,34,35,36] | |
| Acute | IL-17, IL-10 R | Downregulation | develop joint symptoms in acute phase and maintain these symptoms in the chronic phase | [35] | ||
| Chronic | CD95L | Upregulation | involved in CHIKV chronicity mechanism | [35] | ||
| CD4+ T cells | Acute | CD95L | Upregulation | apoptosis of CD4 T cells | [32] | |
| Chronic | , | Upregulation | inflammation and joint swelling | [39] | ||
| T cells | Acute | Downregulation | reduced foot swelling in mice | [45,46,109] | ||
| CD4-Treg | recovery | IL-10 | Up regulation | resolution of the disease | [48] | |
| DENV | CD8+ T cells | Acute | CCR5, CXCR3, CXCR6 | Upregulation | supporting T cells to migrate in to peripheral inflamed tissues | [53,54] |
| CLA | Upregulation | influx of T cells to skin providing immediate onsite protection | [117] | |||
| CD69, HLA-DP, DQ, DR, CD38, cytotoxic granule TIA-1, VLA-4, ICAM-1, LFA-1CD44, CD11a | Upregulation | T cell activation, elimination the virus, induce inflammation | [117] | |||
| HLA-DR+CD38+ CD8+T cells | Acute | PD-1, Lag3, KLRG1, CTLA-4, CD160 | Upregulation | T cell activation, Proliferation, cytotoxicity and migration | [72] | |
| HLA-DR−CD38+ CD8+ T cells | Acute | AKT3, ACTN1 | Downregulation | TCR signaling, amplification and synapse | [72] | |
| CD8+Tem (CD45RA−CCR7−) and Temra (CD45RA+CCR7−) | Acute | , TNFα, CTLA4, ICOS, LIGHT, IRF4, IRF8, SLAMF7, KIR2DL3 | Upregulation | T cell activation, proliferation and polyfunctional properties, narrow transcriptional responses | [73] | |
| CXCR5+CD8+ T cells (TFH) | Chronic | PD-1 | Upregulation | T cell proliferation and exert effector functions | [74] | |
| CD4+ T cells | Upregulation | cytotoxic effects | [77,79] | |||
| CD4+Temra CD45RA+CCR7− | Upregulation | cytotoxic, protective role in DENV with HLA DR allele | [81] | |||
| CD4+CD25+FoxP3+ Treg | acute | CTLA-4 | Upregulation | producing immunosuppressive cytokines | [82,83] | |
| TFH | critical | CD40L | Upregulation | strong activator of B cells | [86,87] | |
| TFHCXCR5+CD45RA−CD4+Tcells | Critical | PD-1, CD38 | Upregulation | predictor for neutralizing antibody titer and disease severity | [85] | |
| ZIKV | CD8+IFN-γ+ T cells | acute | granzyme B, CRTAM, X-C motif chemokine ligand 1 and 2, CC3, CC4 | Upregulation | cytotoxicity, T cell activation and regulation, proinflammation, T cell homing | [101] |
| CD8+IFN-γ+ T cells | Acute | , TNF-α, Granzyme B | Upregulation | Th1, Th2, Th17 and Th9 responses | [17,102] | |
| 2 T cells | Acute | Granzyme B | Upregulation | antiviral and stimulatory activities | [115,116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mapalagamage, M.; Weiskopf, D.; Sette, A.; De Silva, A.D. Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections. Viruses 2022, 14, 242. https://doi.org/10.3390/v14020242
Mapalagamage M, Weiskopf D, Sette A, De Silva AD. Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections. Viruses. 2022; 14(2):242. https://doi.org/10.3390/v14020242
Chicago/Turabian StyleMapalagamage, Maheshi, Daniela Weiskopf, Alessandro Sette, and Aruna Dharshan De Silva. 2022. "Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections" Viruses 14, no. 2: 242. https://doi.org/10.3390/v14020242
APA StyleMapalagamage, M., Weiskopf, D., Sette, A., & De Silva, A. D. (2022). Current Understanding of the Role of T Cells in Chikungunya, Dengue and Zika Infections. Viruses, 14(2), 242. https://doi.org/10.3390/v14020242