
West Nile and Usutu Virus Introduction via Migratory Birds: A Retrospective Analysis in Italy

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
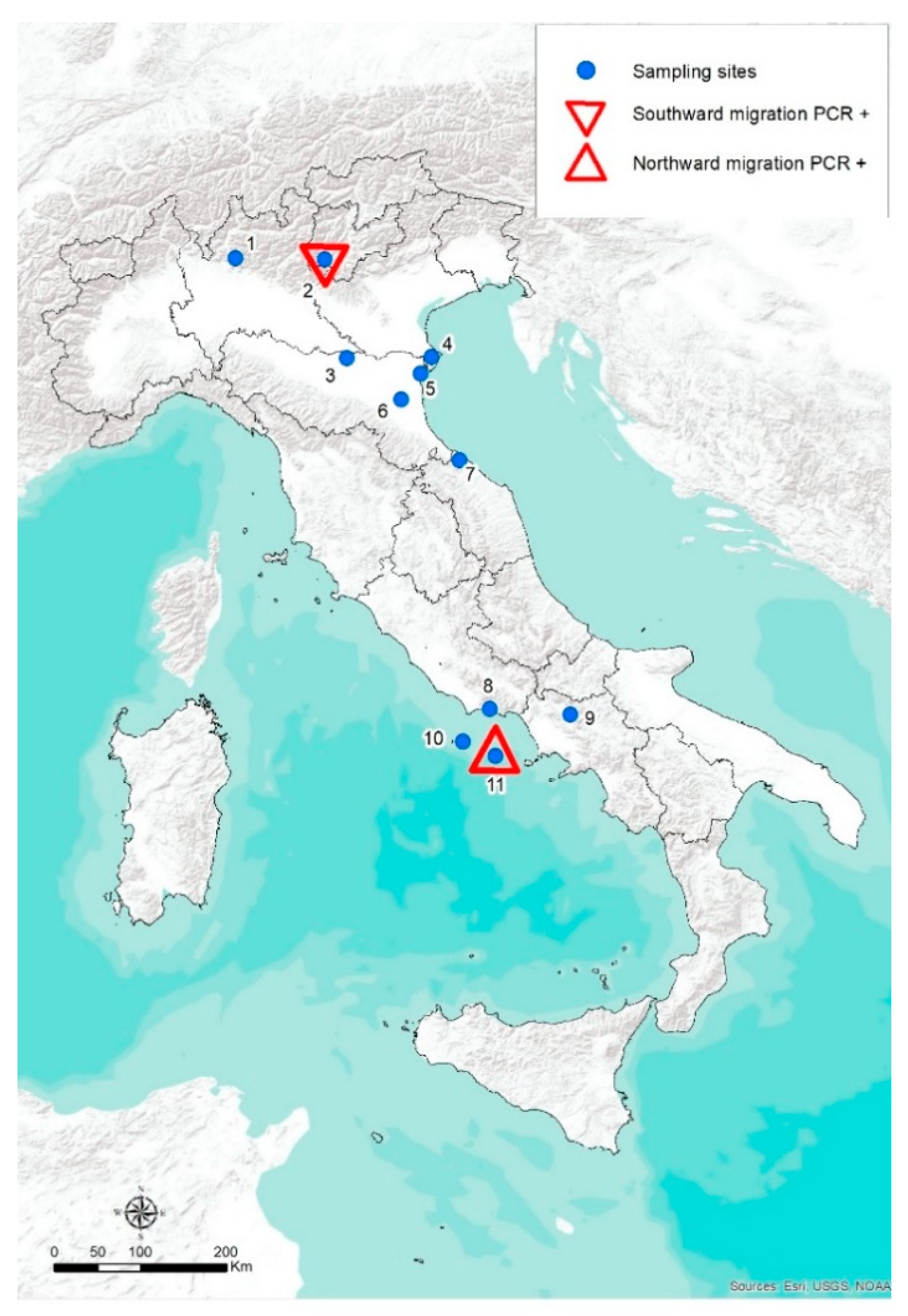
2.1. Study Areas and Bird Trapping Techniques
2.2. Sampling Protocols
2.3. Molecular and Serological Tests
2.4. Statistical Analyses
3. Results
3.1. Samples and Species Composition
3.2. Molecular and Serological Tests
3.3. Statistical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hubálek, Z.; Halouzka, J. West Nile Fever—A Reemerging Mosquito-Borne Viral Disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Cosseddu, G.M.; Sozio, G.; Valleriani, F.; Di Gennaro, A.; Pascucci, I.; Gavaudan, S.; Marianneau, P.; Monaco, F. Serological Survey of Hantavirus and Flavivirus Among Wild Rodents in Central Italy. Vector-Borne Zoonotic Dis. 2017, 17, 777–779. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Becker, N.; Campos, R.D.M.; Börstler, J.; Jöst, H.; Schmidt-Chanasit, J. Usutu Virus in Bats, Germany, 2013. Emerg. Infect. Dis. 2014, 20, 1771–1773. [Google Scholar] [CrossRef] [PubMed]
- Bowen, R.A.; Nemeth, N.M. Experimental infections with West Nile virus. Curr. Opin. Infect. Dis. 2007, 20, 293–297. [Google Scholar] [CrossRef]
- Semenza, J.C.; Tran, A.; Espinosa, L.; Sudre, B.; Domanovic, D.; Paz, S. Climate change projections of West Nile virus infections in Europe: Implications for blood safety practices. Environ. Health 2016, 15, 125–136. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, J.S.; Williams, D.T. The Zoonotic Flaviviruses of Southern, South-Eastern and Eastern Asia, and Australasia: The Potential for Emergent Viruses. Zoonoses Public Health 2009, 56, 338–356. [Google Scholar] [CrossRef]
- Vázquez, A.; Sánchez-Seco, M.P.; Ruiz, S.; Molero, F.; Hernández, L.; Moreno, J.; Magallanes, A.; Tejedor, C.G.; Tenorio, A. Putative New Lineage of West Nile Virus, Spain. Emerg. Infect. Dis. 2010, 16, 549–552. [Google Scholar] [CrossRef]
- Calistri, P.; Giovannini, A.; Savini, G.; Monaco, F.; Bonfanti, L.; Ceolin, C.; Terregino, C.; Tamba, M.; Cordioli, P.; Lelli, R. West Nile Virus Transmission in 2008 in North-Eastern Italy. Zoonoses Public Health 2010, 57, 211–219. [Google Scholar] [CrossRef]
- Rizzoli, A.; Jiménez-Clavero, M.A.; Barzon, L.; Cordioli, P.; Figuerola, J.; Koraka, P.; Martina, B.; Moreno, A.; Nowotny, N.; Pardigon, N.; et al. The challenge of West Nile virus in Europe: Knowledge gaps and research priorities. Eurosurveillance 2015, 20, 21135. [Google Scholar] [CrossRef] [Green Version]
- Monaco, F.; Çizmeci, Ş.; Polci, A.; Portanti, O.; Barut, F.; Deniz, A.; Cosseddu, G.M.; Pişkin, Ç.; Savini, G. West Nile virus lineage 2 circulation in Turkey. Vet. Ital. 2016, 51, 77–81. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Tabain, I.; Hrnjakovic-Cvjetkovic, I.; Bogdanic, M.; Klobucar, A.; et al. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Vet. Sci. 2019, 6, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.F.; Popovici, F.; Cernescu, C.; Campbell, G.L.; Nedelcu, N.I. West Nile encephalitis epidemic in southeastern Romania. Lancet 1998, 352, 767–771. [Google Scholar] [CrossRef]
- Danis, K.; Papa, A.; Theocharopoulos, G.; Dougas, G.; Athanasiou, M.; Detsis, M.; Baka, A.; Lytras, T.; Mellou, K.; Bonovas, S.; et al. Outbreak of West Nile virus infection in Greece, 2010. Emerg. Infect. Dis. 2011, 17, 1868. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Historical Data by Year—West Nile Fever Seasonal Surveillance 2018. Available online: https://ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical (accessed on 25 January 2022).
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile Virus Responsible for an Outbreak of Encephalitis in the Northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakonyi, T.; Ivanics, É.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, N. Lineage 1 and 2 Strains of Encephalitic West Nile Virus, Central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Venter, M.; Swanepoel, R. West Nile Virus Lineage 2 as a Cause of Zoonotic Neurological Disease in Humans and Horses in Southern Africa. Vector-Borne Zoonotic Dis. 2010, 10, 659–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, F.; Goffredo, M.; Briguglio, P.; Pinoni, C.; Polci, A.; Iannetti, S.; Pinto, S.M.; Marruchella, G.; Di Francesco, G.; Di Gennaro, A.; et al. The 2011 West Nile disease outbreak in Sardinia region, Italy. Vet. Ital. 2015, 51, 5–16. [Google Scholar] [CrossRef]
- Hubálek, Z.; Kosina, M.; Rudolf, I.; Mendel, J.; Straková, P.; Tomešek, M. Mortality of Goshawks (Accipiter gentilis) due to West Nile virus lineage 2. Vector-Borne Zoonotic Dis. 2018, 18, 624–627. [Google Scholar] [CrossRef]
- Hernãndez-Triana, L.M.; Jeffries, C.L.; Mansfield, K.L.; Carnell, G.; Fooks, A.R.; Johnson, N. Emergence of West Nile Virus Lineage 2 in Europe: A Review on the Introduction and Spread of a Mosquito-Borne Disease. Front. Public Health 2014, 2, 271. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Haussig, J.M. West Nile virus keeps on moving up in Europe. Eurosurveillance 2020, 25, 2001938. [Google Scholar] [CrossRef]
- Rizzo, C.; Napoli, C.; Venturi, G.; Pupella, S.; Lombardini, L.; Calistri, P.; Monaco, F.; Cagarelli, R.; Angelini, P.; Bellini, R.; et al. West Nile virus transmission: Results from the integrated surveillance system in Italy, 2008 to 2015. Eurosurveillance 2016, 21, 30340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnarelli, P.; Marinelli, K.; Trotta, D.; Monachetti, A.; Tavio, M.; Del Gobbo, R.; Capobianchi, M.R.; Menzo, S.; Nicoletti, L.; Magurano, F.; et al. Human case of autochthonous West Nile virus lineage 2 infection in Italy, September 2011. Eurosurveillance 2011, 16, 20002. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Capelli, G.; Monaco, F.; Polci, A.; Russo, F.; Di Gennaro, A.; Marini, V.; Teodori, L.; Montarsi, F.; Pinoni, C.; et al. Evidence of West Nile virus lineage 2 circulation in Northern Italy. Vet. Microbiol. 2012, 158, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Pacenti, M.; Franchin, E.; Squarzon, L.; Lavezzo, E.; Cattai, M.; Cusinato, R.; Palù, G. The Complex Epidemiological Scenario of West Nile Virus in Italy. Int. J. Environ. Res. Public Health 2013, 10, 4669–4689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne Flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenböck, H. Pathology and Viral Distribution in Fatal Usutu Virus Infections of Birds from the 2001 and 2002 Outbreaks in Austria. J. Comp. Pathol. 2004, 131, 176–185. [Google Scholar] [CrossRef]
- Savini, G.; Monaco, F.; Terregino, C.; Di Gennaro, A.; Bano, L.; Pinoni, C.; De Nardi, R.; Bonilauri, P.; Pecorari, M.; Di Gialleonardo, L.; et al. Usutu virus in ITALY: An emergence or a silent infection? Vet. Microbiol. 2011, 151, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Garigliany, M.; Linden, A.; Gilliau, G.; Levy, E.; Sarlet, M.; Franssen, M.; Benzarti, E.; Derouaux, A.; Francis, F.; Desmecht, D. Usutu virus, Belgium, 2016. Infect. Genet. Evol. 2017, 48, 116–119. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Munnink, B.B.O.; Münger, E.; Nieuwenhuijse, D.F.; Kohl, R.; Van Der Linden, A.; Schapendonk, C.M.E.; Van Der Jeugd, H.; Kik, M.; Rijks, J.M.; Reusken, C.B.E.M.; et al. Genomic monitoring to understand the emergence and spread of Usutu virus in the Netherlands, 2016–2018. Sci. Rep. 2020, 10, 2798. [Google Scholar] [CrossRef] [Green Version]
- Folly, A.J.; Lawson, B.; Lean, F.Z.; McCracken, F.; Spiro, S.; John, S.K.; Heaver, J.P.; Seilern-Moy, K.; Masters, N.; Hernández-Triana, L.M.; et al. Detection of Usutu virus infection in wild birds in the United Kingdom, 2020. Eurosurveillance 2020, 25, 2001732. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A.M.; Rossini, G.; Bonilauri, P.; Gerunda, G.E.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; et al. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August–September 2009. Eurosurveillance 2009, 14, 19448. [Google Scholar] [CrossRef] [PubMed]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.M.; Tagliazucchi, S.; Savini, G.; Monaco, F.; Simone, M.L.; Lelli, R.; et al. First human case of Usutu virus neuroinvasive infection, Italy, August–September 2009. Eurosurveillance 2009, 14, 19446. [Google Scholar] [CrossRef] [PubMed]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savić, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August–September 2013: Clinical and laboratory features. J. Neurovirol. 2014, 21, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveillance 2017, 22, 30452. [Google Scholar] [CrossRef]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M.F.; Girard, G.; Van de Perre, P.; Salinas, S.; et al. Human Usutu Virus Infection with Atypical Neurologic Presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Jungbauer, C.; Aberle, S.W.; Kolodziejek, J.; Dimmel, K.; Stiasny, K.; Allerberger, F.; Nowotny, N. Usutu virus infections among blood donors, Austria, July and August 2017—Raising awareness for diagnostic challenges. Eurosurveillance 2017, 22, 17-00644. [Google Scholar] [CrossRef]
- Cadar, D.; Maier, P.; Müller, S.; Kress, J.; Chudy, M.; Bialonski, A.; Schlaphof, A.; Jansen, S.; Jöst, H.; Tannich, E.; et al. Blood donor screening for West Nile virus (WNV) revealed acute Usutu virus (USUV) infection, Germany, September 2016. Eurosurveillance 2017, 22, 30501. [Google Scholar] [CrossRef]
- Zaaijer, H.L.; Slot, E.; Molier, M.; Reusken, C.B.; Koppelman, M.H. Usutu virus infection in Dutch blood donors. Transfusion 2019, 59, 2931–2937. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental Infection of North American Birds with the New York 1999 Strain of West Nile Virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: A review. Vet. Res. 2013, 44, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spedicato, M.; Carmine, I.; Bellacicco, A.L.; Marruchella, G.; Marini, V.; Pisciella, M.; DI Francesco, G.; Lorusso, A.; Monaco, F.; Savini, G. Experimental infection of rock pigeons (Columba livia) with three West Nile virus lineage 1 strains isolated in Italy between 2009 and 2012. Epidemiol. Infect. 2016, 144, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramírez, E.; Llorente, F.; Jiménez-Clavero, M. Ángel Experimental Infections of Wild Birds with West Nile Virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [Green Version]
- Owen, J.; Moore, F.; Panella, N.; Edwards, E.; Bru, R.; Hughes, M.; Komar, N. Migrating Birds as Dispersal Vehicles for West Nile Virus. EcoHealth 2006, 3, 79–85. [Google Scholar] [CrossRef]
- McGUILL, M.W.; Rowan, A.N. Biological Effects of Blood Loss: Implications for Sampling Volumes and Techniques. ILAR J. 1989, 31, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Gould, E.; Kolodziejek, J.; Weissenbock, H.; Nowotny, N. Complete genome analysis and molecular characterization of Usutu virus that emerged in Austria in 2001: Comparison with the South African Strain SAAR-1776 and other flaviviruses. Virology 2004, 328, 301–310. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csörgő, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; et al. Emergence of Usutu Virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef] [Green Version]
- Del Amo, J.; Sotelo, E.; Fernández-Pinero, J.; Gallardo, C.; Llorente, F.; Agüero, M.; Jimenez-Clavero, M.A. A novel quantitative multiplex real-time RT-PCR for the simultaneous detection and differentiation of West Nile virus lineages 1 and 2, and of Usutu virus. J. Virol. Methods 2013, 189, 321–327. [Google Scholar] [CrossRef]
- Di Gennaro, A.; Lorusso, A.; Casaccia, C.; Conte, A.; Monaco, F.; Savini, G. Serum Neutralization Assay Can Efficiently Replace Plaque Reduction Neutralization Test for Detection and Quantitation of West Nile Virus Antibodies in Human and Animal Serum Samples. Clin. Vaccine Immunol. 2014, 21, 1460–1462. [Google Scholar] [CrossRef] [Green Version]
- OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animal 2021, West Nile Fever (Version Adopted in May 2018) Chapter 3.1.24. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.24_WEST_NILE.pdf (accessed on 28 January 2022).
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic Relationships between Flaviviruses as Determined by Cross-neutralization Tests with Polyclonal Antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Spina, F.; Volponi, S. Atlante della Migrazione degli Uccelli in Italia. 2. Passeriformi. Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA); Tipografia SCR-Roma: Roma, Italy, 2008. [Google Scholar]
- Andreotti, A.; Bendini, L.; Piacentini, D.; Spina, F. The role of Italy within the Song Thrush (Turdus philomelos) migratory system analysed on the basis of ringing-recovery data. Vogelwarte 1999, 40, 28–51. [Google Scholar]
- Filipe, A.R.; Pinto, M.R. Survey for Antibodies to Arboviruses in Serum of Animals from Southern Portugal. Am. J. Trop. Med. Hyg. 1969, 18, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidemiology of the West Nile virus: Study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse. Ann. de l’Institut Pasteur 1970, 118, 239–247. [Google Scholar]
- Paz, S.; Malkinson, D.; Green, M.S.; Tsioni, G.; Papa, A.; Danis, K.; Sirbu, A.; Ceianu, C.; Katalin, K.; Ferenczi, E.; et al. Permissive Summer Temperatures of the 2010 European West Nile Fever Upsurge. PLoS ONE 2013, 8, e56398. [Google Scholar] [CrossRef] [Green Version]
- Young, J.J.; Haussig, J.M.; Aberle, S.W.; Pervanidou, D.; Riccardo, F.; Sekulić, N.; Bakonyi, T.; Gossner, C.M. Epidemiology of human West Nile virus infections in the European Union and European Union enlargement countries, 2010 to 2018. Eurosurveillance 2021, 26, 2001095. [Google Scholar] [CrossRef]
- Sikkema, R.S.; Schrama, M.; Berg, T.V.D.; Morren, J.; Munger, E.; Krol, L.; Van Der Beek, J.G.; Blom, R.; Chestakova, I.; Van Der Linden, A.; et al. Detection of West Nile virus in a common whitethroat (Curruca communis) and Culex mosquitoes in the Netherlands, 2020. Eurosurveillance 2020, 25, 2001704. [Google Scholar] [CrossRef]
- Beck, C.; Goffart, I.L.; Franke, F.; Gonzalez, G.; Dumarest, M.; Lowenski, S.; Blanchard, Y.; Lucas, P.; Lamballerie, X.; Grard, G.; et al. Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019. Pathogens 2020, 9, 908. [Google Scholar] [CrossRef]
- Rodríguez-Alarcón, L.G.S.M.; Fernández-Martínez, B.; Moros, M.J.S.; Vázquez, A.; Pachés, P.J.; Villacieros, E.G.; Martín, M.B.G.; Borras, J.F.; Lorusso, N.; Aceitero, J.M.R.; et al. Unprecedented increase of West Nile virus neuroinvasive disease, Spain, summer 2020. Eurosurveillance 2021, 26, 2002010. [Google Scholar] [CrossRef]
- Zehender, G.; Veo, C.; Ebranati, E.; Carta, V.; Rovida, F.; Percivalle, E.; Moreno, A.; Lelli, D.; Calzolari, M.; Lavazza, A.; et al. Reconstructing the recent West Nile virus lineage 2 epidemic in Europe and Italy using discrete and continuous phylogeography. PLoS ONE 2017, 12, e0179679. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control (ECDC). Historical Data by Year—West Nile Fever Seasonal Surveillance 2020. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-west-nile-virus-transmission-season-europe-2020 (accessed on 18 November 2021).
- Kramer, L.D.; Bernard, K.A. West Nile Virus Infection in Birds and Mammals. Ann. N. Y. Acad. Sci. 2006, 951, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Fang, Y.; Lothrop, H.D.; Martinez, V.M.; Wilson, J.; Oconnor, P.; Carney, R.; Cahoon-Young, B.; Shafii, M.; Brault, A.C. Overwintering of West Nile virus in Southern California. J. Med. Entomol. 2006, 43, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.; Young, G.; Ndaluka, C.; Bielefeldt-Ohmann, H.; Komar, N.; Bowen, R. Persistent West Nile virus infection in the house sparrow (Passer domesticus). Arch. Virol. 2009, 154, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Lafri, I.; Hachid, A.; Bitam, I. West Nile virus in Algeria: A comprehensive overview. New Microbes New Infect. 2018, 27, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Amdouni, J.; Monaco, F.; Portanti, O.; Sghaier, S.; Conte, A.; Ben Hassine, T.; Polci, A.; Valleriani, F.; Di Gennaro, A.; Zoueri, M.; et al. Detection of enzootic circulation of a new strain of West Nile virus lineage 1 in sentinel chickens in the north of Tunisia. Acta Trop. 2019, 202, 105223. [Google Scholar] [CrossRef]
- Conte, A.; Candeloro, L.; Ippoliti, C.; Monaco, F.; De Massis, F.; Bruno, R.; Di Sabatino, D.; Danzetta, M.L.; Benjelloun, A.; Belkadi, B.; et al. Spatio-Temporal Identification of Areas Suitable for West Nile Disease in the Mediterranean Basin and Central Europe. PLoS ONE 2015, 10, e0146024. [Google Scholar] [CrossRef]
- Erdélyi, K.; Ursu, K.; Ferenczi, E.; Szeredi, L.; Rátz, F.; Skáre, J.; Bakonyi, T. Clinical and Pathologic Features of Lineage 2 West Nile Virus Infections in Birds of Prey in Hungary. Vector-Borne Zoonotic Dis. 2007, 7, 181–188. [Google Scholar] [CrossRef]
- Vidaña, B.; Busquets, N.; Napp, S.; Pérez-Ramírez, E.; Jiménez-Clavero, M.Á.; Johnson, N. The Role of Birds of Prey in West Nile Virus Epidemiology. Vaccines 2020, 8, 550. [Google Scholar] [CrossRef]
- Ana, A.; Perez Andrés, M.; Julia, P.; Puig, P.; Wünschmann, A.; Vander Waal, K.; Alvarez, J.; Willette, M. Syndromic surveillance for West Nile virus using raptors in rehabilitation. BMC Vet. Res. 2017, 13, 368. [Google Scholar] [CrossRef] [Green Version]
- Figuerola, J.; Jimenez-Clavero, M.A.; López, G.; Rubio, C.; Soriguer, R.; Gómez-Tejedor, C.; Tenorio, A. Size matters: West Nile Virus neutralizing antibodies in resident and migratory birds in Spain. Vet. Microbiol. 2008, 132, 39–46. [Google Scholar] [CrossRef]
- Valiakos, G.; Touloudi, A.; Athanasiou, L.V.; Giannakopoulos, A.; Iacovakis, C.; Birtsas, P.; Spyrou, V.; Dalabiras, Z.; Petrovska, L.; Billinis, C. Serological and molecular investigation into the role of wild birds in the epidemiology of West Nile virus in Greece. Virol. J. 2012, 9, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernek, E.; Kozuch, O.; Nosek, J.; Teplan, J.; Folk, C. Arboviruses in birds captured in Slovakia. J. Hyg. Epidemiol. Microbiol. Immunol. 1977, 21, 353–359. [Google Scholar] [PubMed]
- López, G.; Jiménez-Clavero, M.Á.; Tejedor, C.G.; Soriguer, R.; Figuerola, J. Prevalence of West Nile Virus Neutralizing Antibodies in Spain Is Related to the Behavior of Migratory Birds. Vector-Borne Zoonotic Dis. 2008, 8, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Llopis, I.V.; Rossi, L.; Di Gennaro, A.; Mosca, A.; Teodori, L.; Tomassone, L.; Grego, E.; Monaco, F.; Lorusso, A.; Savini, G. Further circulation of West Nile and Usutu viruses in wild birds in Italy. Infect. Genet. Evol. 2015, 32, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, S.E.J.; Hoffman, D.M.; Stark, L.M.; Marlenee, N.L.; Blitvich, B.J.; Beaty, B.J.; Stallknecht, D.E. Persistence of Antibodies to West Nile Virus in Naturally Infected Rock Pigeons (Columba livia). Clin. Vaccine Immunol. 2005, 12, 665–667. [Google Scholar] [CrossRef] [Green Version]
- Figuerola, J.; Soriguer, R.; Rojo, G.; Tejedor, C.G.; Jimenez-Clavero, M.A. Seroconversion in Wild Birds and Local Circulation of West Nile Virus, Spain. Emerg. Infect. Dis. 2007, 13, 1915–1917. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef]
- Rappole, J.H.; Hubálek, Z. Migratory birds and West Nile virus. J. Appl. Microbiol. 2003, 94, 47–58. [Google Scholar] [CrossRef]
- Mencattelli, G.; Iapaolo, F.; Monaco, F.; Fusco, G.; de Martinis, C.; Portanti, O.; Di Gennaro, A.; Curini, V.; Polci, A.; Berjaoui, S.; et al. West Nile Virus Lineage 1 in Italy: Newly Introduced or a Re-Occurrence of a Previously Circulating Strain? Viruses 2021, 14, 64. [Google Scholar] [CrossRef]
- Zecchin, B.; Fusaro, A.; Milani, A.; Schivo, A.; Ravagnan, S.; Ormelli, S.; Mavian, C.; Michelutti, A.; Toniolo, F.; Barzon, L.; et al. The central role of Italy in the spatial spread of USUTU virus in Europe. Virus Evol. 2021, 7, veab048. [Google Scholar] [CrossRef]


{kind=link}
| Year | Sampling Season | Order | Species | Migratory Strategy 1 | N. Birds per Sample | N. Positive Samples | Total Positive Samples | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WNV ND50 2 | USUV ND50 | ||||||||||||||
| 1:10 | 1:20 | 1:40 | 1:80 | 1:160 | 1:320 | 1:10 | 1:20 | 1:40 | |||||||
| 2013 | Spring | Bucerotiformes | Hoopoe (Upupa epops) | L | 1 | 1 | 1 | ||||||||
| 1 | 1 | 1 | |||||||||||||
| 1 | 2 | 2 | |||||||||||||
| Caprimulgiformes | European Nightjar (Caprimulgus europaeus) | L | 2 | 1 | 1 | ||||||||||
| Charadriiformes | Ruff (Philomachus pugnax) | L | 1 | 1 | 1 | ||||||||||
| Wood Sandpiper (Tringa glareola) | L | 2 | 1 | 1 | |||||||||||
| 3 | 1 | 1 | |||||||||||||
| Columbiformes | European Turtle Dove (Streptopelia turtur) | L | 1 | 4 | 5 | 2 | 1 | 13 | |||||||
| Galliformes | Common Quail (Coturnix coturnix) | L | 1 | 1 | 1 | 3 | |||||||||
| Passeriformes | Common Whitethroat (Sylvia communis) | L | 8 | 4 | 4 | ||||||||||
| 9 | 1 | 1 | 2 | ||||||||||||
| Eurasian Golden Oriole (Oriolus oriolus) | L | 1 | 3 | 1 | 2 | 6 | |||||||||
| 2 | 1 | 1 | |||||||||||||
| Garden Warbler (Sylvia borin) | L | 8 | 1 | 1 | |||||||||||
| 9 | 1 | 1 | |||||||||||||
| Whinchat (Saxicola rubetra) | L | 8 | 1 | 1 | |||||||||||
| Strigiformes | Eurasian Scops-Owl (Otus scops) | L | 1 | 1 | 1 | ||||||||||
| Autumn | Charadriiformes | Redshank (Tringa totanus) | L | 1 | 1 | 1 | |||||||||
| Coraciiformes | European Bee Eater (Merops apiaster) | L | 1 | 1 | 1 | 2 | |||||||||
| Galliformes | Common Quail (Coturnix coturnix) | L | 1 | 1 | 1 | ||||||||||
| Gruiformes | Spotted Crake (Porzana porzana) | L | 1 | 3 | 3 | ||||||||||
| Passeriformes | Song Thrush (Turdus philomelos) | P | 2 | 1 | 1 | ||||||||||
| 2014 | Spring | Accipitriformes | Eurasian Sparrowhawk (Accipiter nisus) | P | 1 | 1 | 1 | ||||||||
| Marsh Harrier (Circus aeruginosus) | P | 1 | 1 | 1 | |||||||||||
| Bucerotiformes | Hoopoe (Upupa epops) | L | 1 | 2 | 1 | 3 | |||||||||
| Columbiformes | European Turtle Dove (Streptopelia turtur) | L | 1 | 5 | 5 | 1 | 11 | ||||||||
| Passeriformes | Common Whitethroat (Sylvia communis) | L | 6 | 1 | 1 | ||||||||||
| 8 | 1 | 1 | |||||||||||||
| Eurasian Golden Oriole (Oriolus oriolus) | L | 2 | 1 | 1 | |||||||||||
| 1 | 1 | 1 | 1 | 1 | 4 | ||||||||||
| 2 | 3 | 3 | |||||||||||||
| Strigiformes | Eurasian Scops-Owl (Otus scops) | L | 1 | 1 | 1 | ||||||||||
| Autumn | Passeriformes | Hawfinch (Coccothraustes coccothraustes) | S | 3 | 2 | 2 | |||||||||
| 3 | 1 | 1 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, E.; Cecere, J.G.; Iapaolo, F.; Di Gennaro, A.; Sacchi, M.; Savini, G.; Spina, F.; Monaco, F. West Nile and Usutu Virus Introduction via Migratory Birds: A Retrospective Analysis in Italy. Viruses 2022, 14, 416. https://doi.org/10.3390/v14020416
Mancuso E, Cecere JG, Iapaolo F, Di Gennaro A, Sacchi M, Savini G, Spina F, Monaco F. West Nile and Usutu Virus Introduction via Migratory Birds: A Retrospective Analysis in Italy. Viruses. 2022; 14(2):416. https://doi.org/10.3390/v14020416
Chicago/Turabian StyleMancuso, Elisa, Jacopo Giuseppe Cecere, Federica Iapaolo, Annapia Di Gennaro, Massimo Sacchi, Giovanni Savini, Fernando Spina, and Federica Monaco. 2022. "West Nile and Usutu Virus Introduction via Migratory Birds: A Retrospective Analysis in Italy" Viruses 14, no. 2: 416. https://doi.org/10.3390/v14020416
APA StyleMancuso, E., Cecere, J. G., Iapaolo, F., Di Gennaro, A., Sacchi, M., Savini, G., Spina, F., & Monaco, F. (2022). West Nile and Usutu Virus Introduction via Migratory Birds: A Retrospective Analysis in Italy. Viruses, 14(2), 416. https://doi.org/10.3390/v14020416