
A Conserved Receptor-Binding Domain in the VP1u of Primate Erythroparvoviruses Determines the Marked Tropism for Erythroid Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, and Bacteria
2.2. PP7 production
2.3. B19V and Simian VP1u Expression
2.4. PP7-VP1u Bioconjugation
2.5. SDS-PAGE and Western blot
2.6. Detection of PP7-VP1u and B19V Uptake by qPCR and Confocal Microscopy
2.7. Detection of Recombinant VP1u in Cells by Confocal Microscopy
2.8. Competition and Neutralization Assays
2.9. In Silico Predictions
3. Results
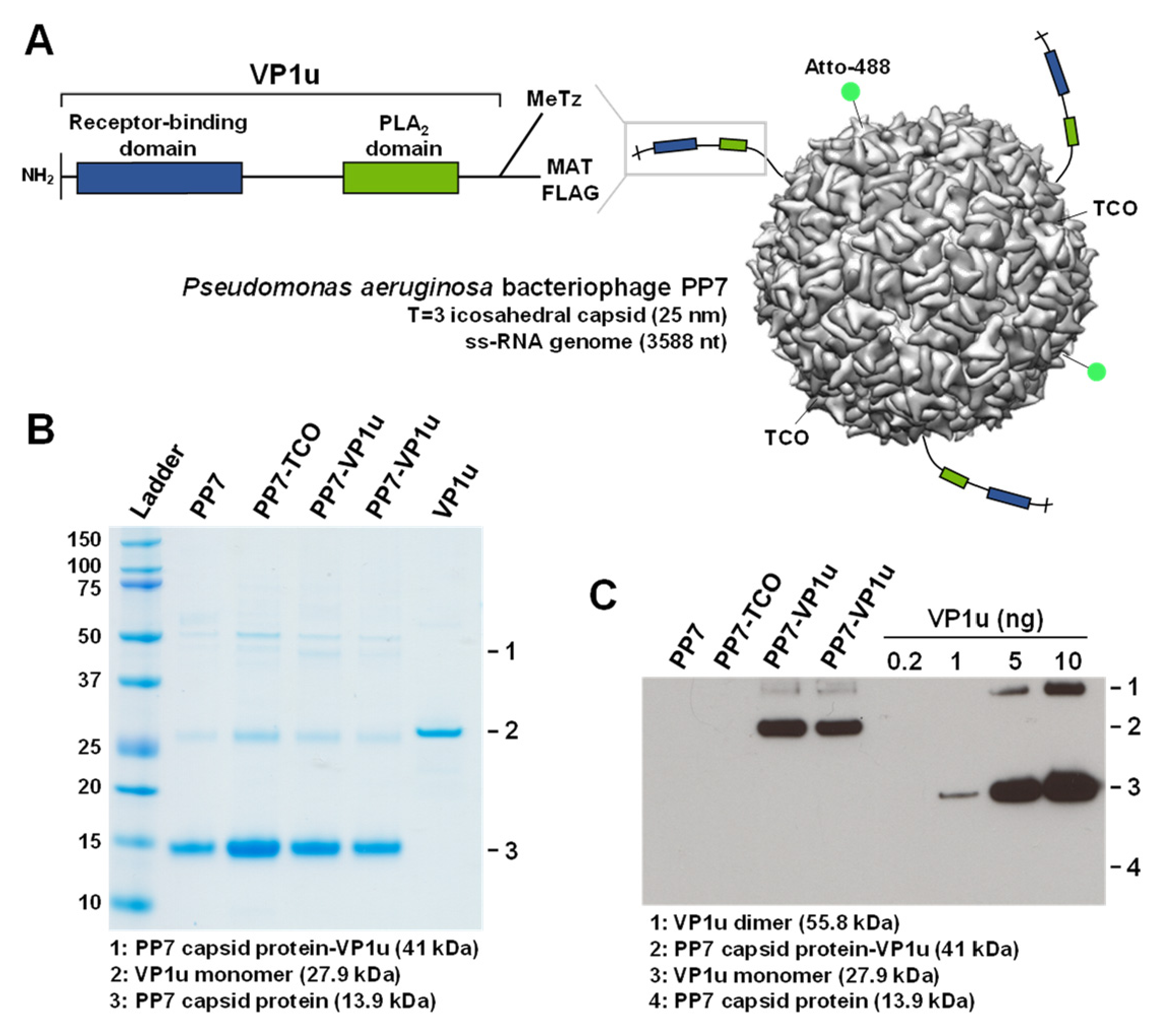
3.1. PP7-VP1u Bioconjugation
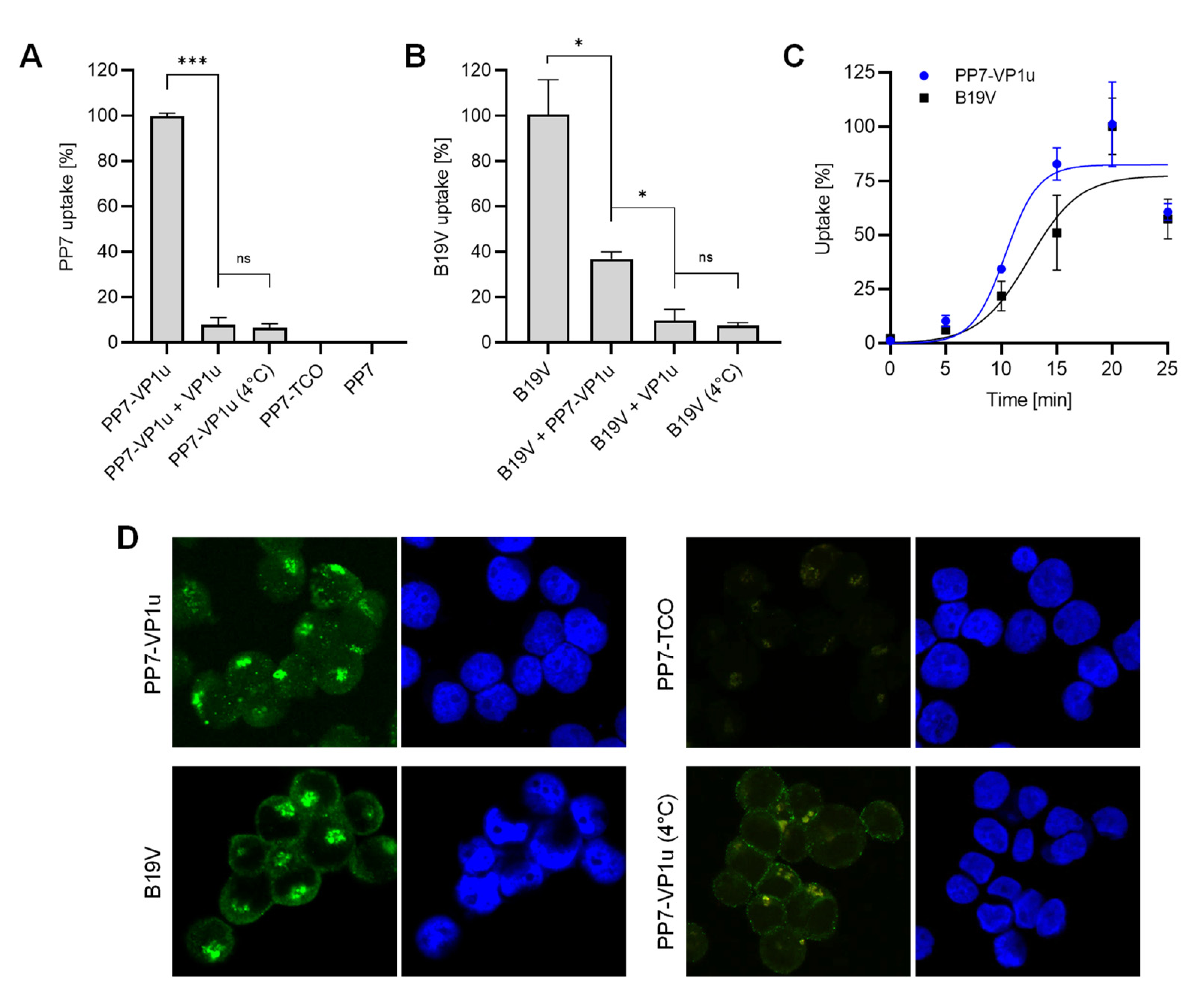
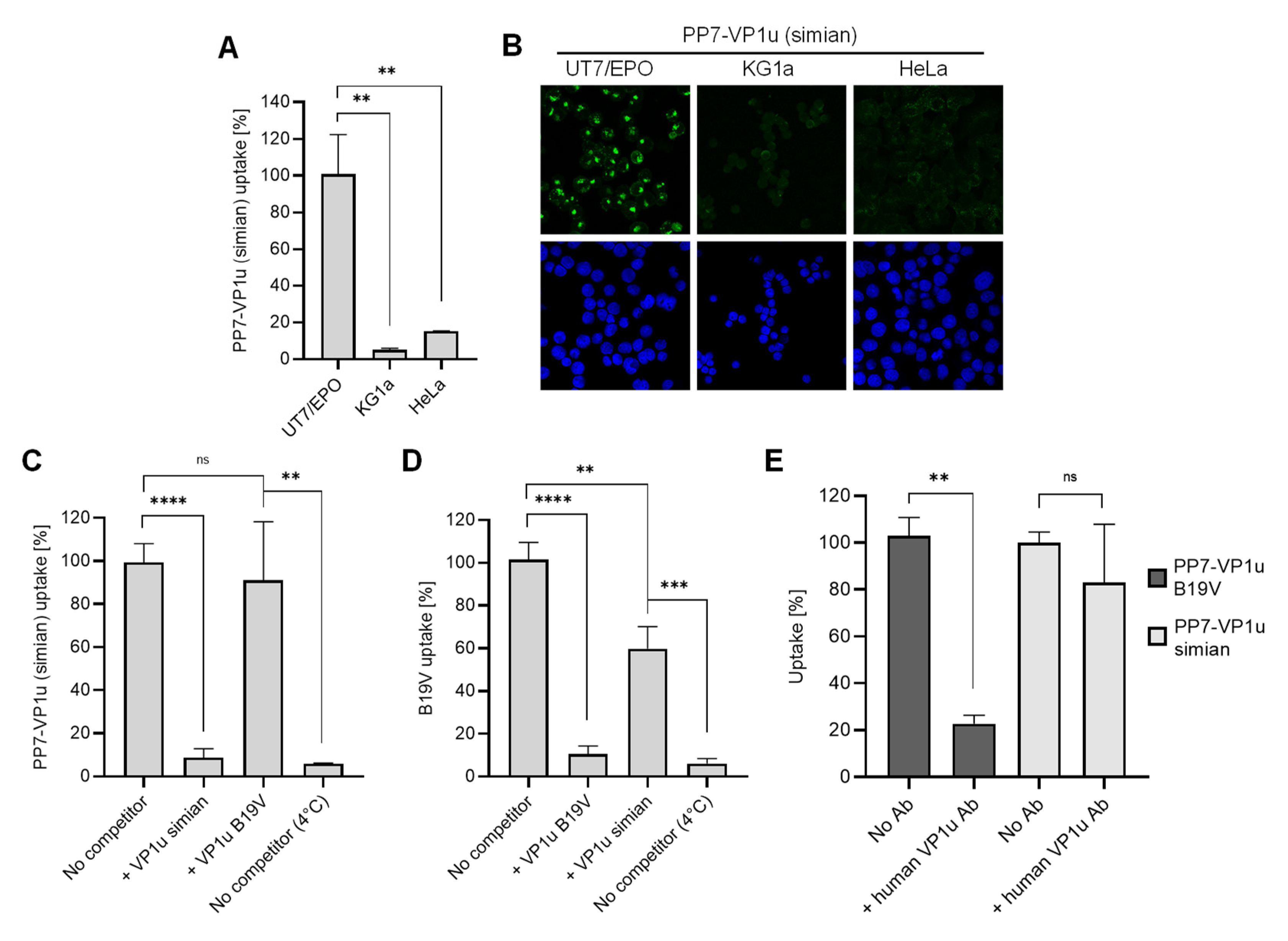
3.2. PP7-VP1u Is Internalized into UT7/Epo Cells and Competes with B19V
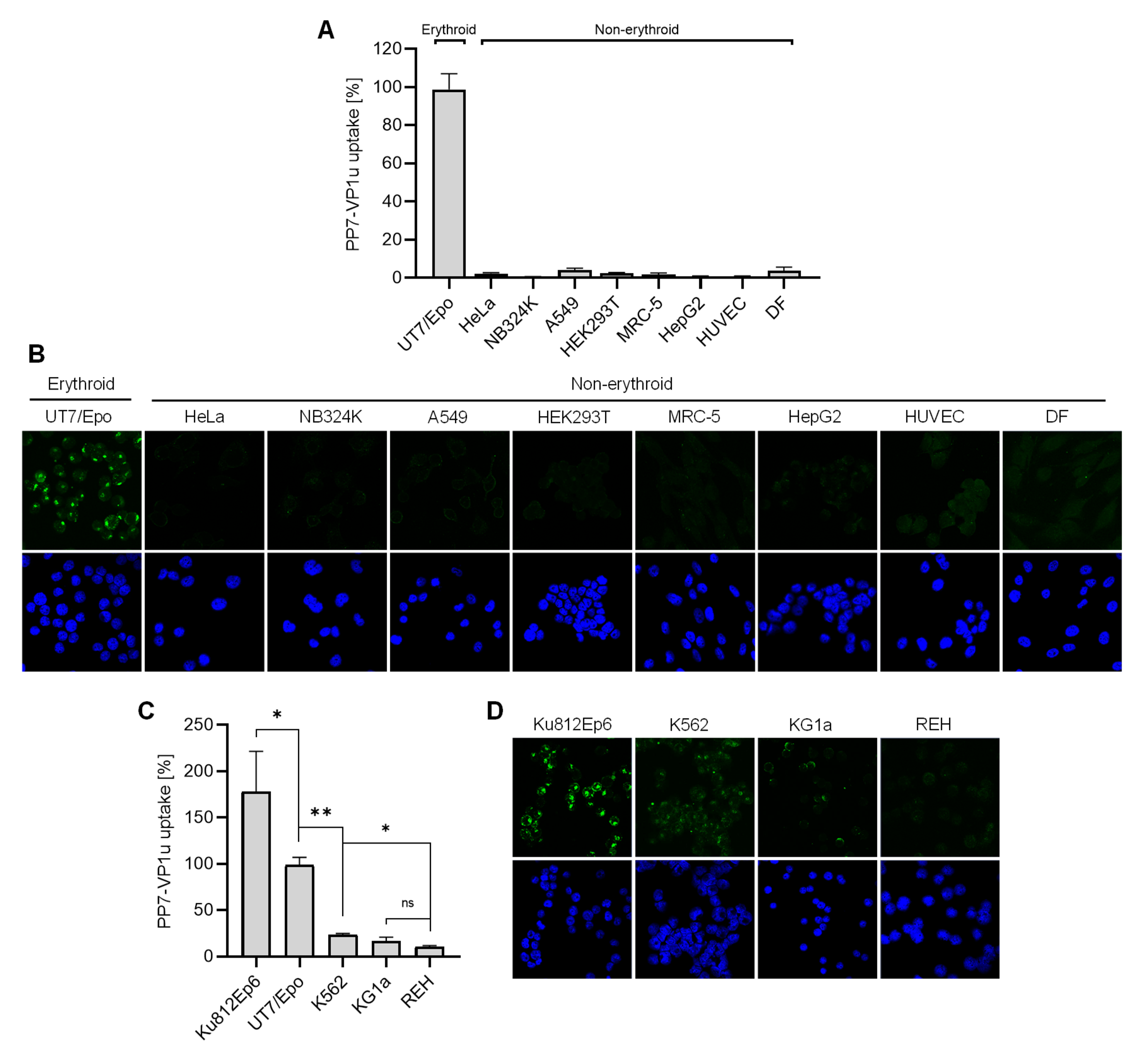
3.3. VP1uR Expression is Restricted to Epo-Dependent Erythroid Cells
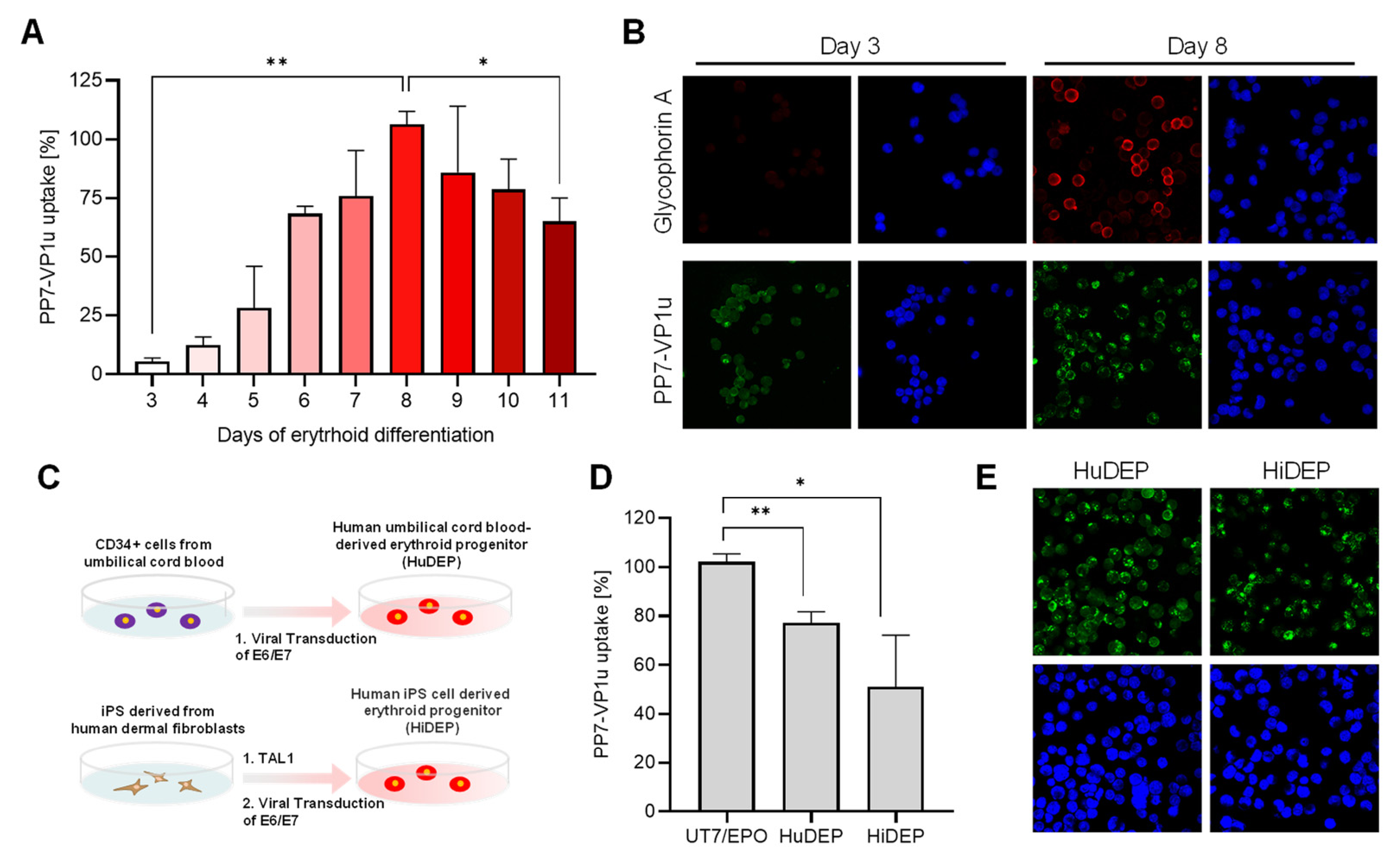
3.4. Quantification of VP1uR Expression at Progressive Erythroid Differentiation Stages
3.5. VP1uR Expression Can Be Induced by Cellular Reprogramming towards Erythroid lineage
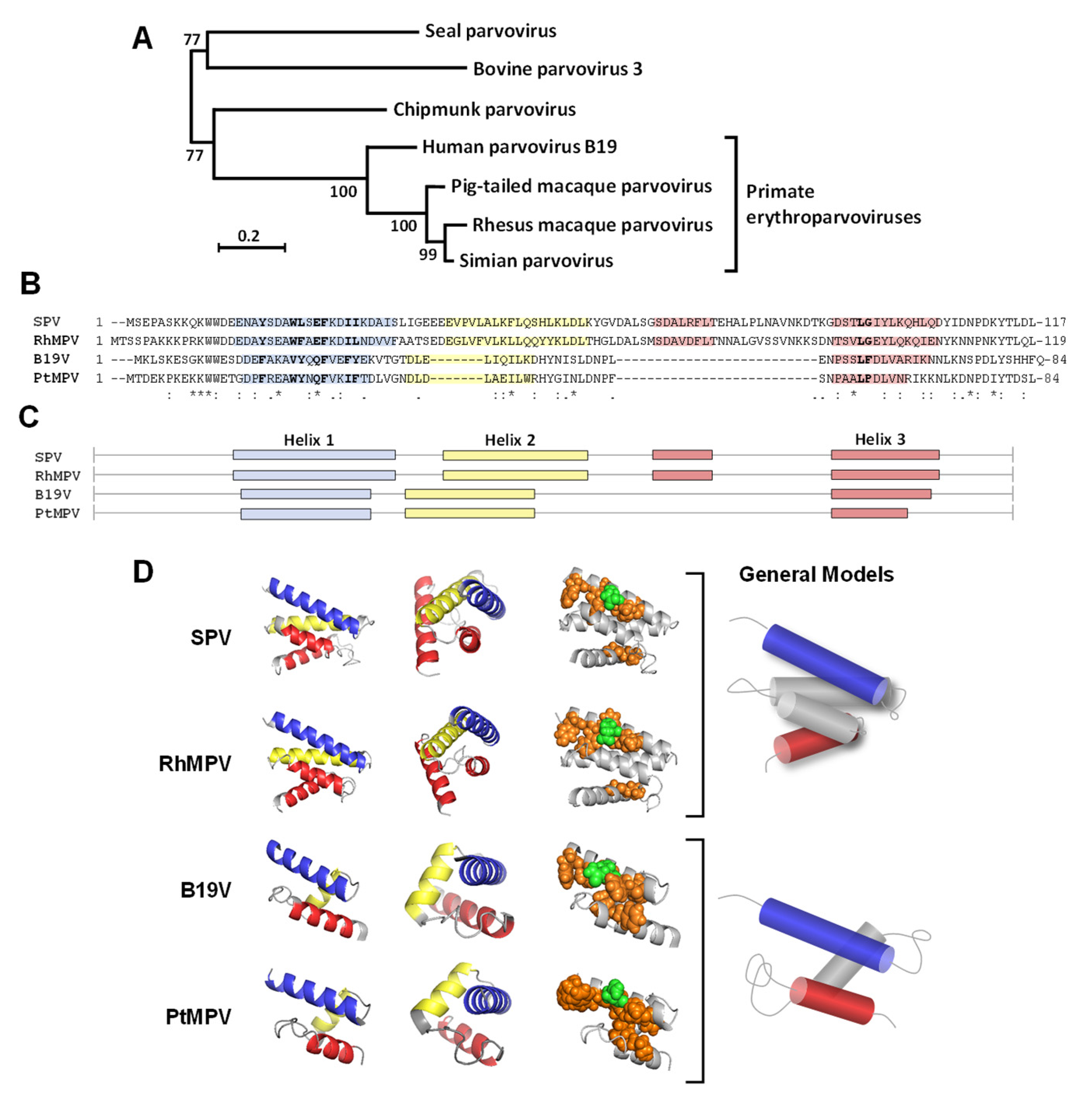
3.6. Structural Comparison of the RBD of B19V and Related Primate Erythroparvoviruses
3.7. N-VP1u of SPV Harbors a Functional RBD, Mediating Virus Uptake into Erythroid Cells
4. Discussion
4.1. Expression Profile of VP1uR
4.2. Characterization of RBDs from Non-Human Primate Erythroparvoviruses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Söderlund-Venermo, M.; Young, N.S. Human Parvoviruses. Clin. Microbiol. Rev. 2017, 30, 43–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Survey, J.T.; Reamy, B.V.; Hodge, J. Clinical presentations of parvovirus B19 infection. Am. Fam. Physician 2007, 75, 75. [Google Scholar]
- Papadogiannakis, N.; Tolfvenstam, T.; Fischler, B.; Norbeck, O.; Broliden, K. Active, Fulminant, Lethal Myocarditis Associated with Parvovirus B19 Infection in an Infant. Clin. Infect. Dis. 2002, 35, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgio, E.; De Oronzo, M.A.; Iozza, I.; Di Natale, A.; Cianci, S.; Garofalo, G.; Giacobbe, A.M.; Politi, S. Parvovirus B19 during pregnancy: A review. J. Prenat. Med. 2010, 4, 63–66. [Google Scholar]
- Ornoy, A.; Ergaz, Z. Parvovirus B19 infection during pregnancy and risks to the fetus. Birth Defects Res. 2017, 109, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Heegaard, E.D.; Brown, K.E. Human Parvovirus B19, Clin. Microbiol. Rev. 2002, 15, 485–505. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.E.; Young, N.S.; Alving, B.M.; Barbosa, L.H. Parvovirus B19: Implications for transfusion medicine. Summary of a workshop. Transfus. 2001, 41, 130–135. [Google Scholar] [CrossRef]
- Eid, A.J.; Brown, R.A.; Patel, R.; Razonable, R.R. Parvovirus B19 Infection after Transplantation: A Review of 98 Cases. Clin. Infect. Dis. 2006, 43, 40–48. [Google Scholar] [CrossRef]
- Parsyan, A.; Candotti, D. Human erythrovirus B19 and blood transfusion? An update. Transfus. Med. 2007, 17, 263–278. [Google Scholar] [CrossRef]
- Stramer, S.L.; Dodd, R.Y.; Subgroup, D. Transfusion-transmitted emerging infectious diseases: 30 years of challenges and progress. Transfus. 2013, 53, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, S.S.; Qiu, J. Recent Advances in Replication and Infection of Human Parvovirus B19. Front. Cell. Infect. Microbiol. 2018, 8, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munakata, Y.; Kato, I.; Saito, T.; Kodera, T.; Ishii, K.K.; Sasaki, T. Human parvovirus B19 infection of monocytic cell line U937 and antibody-dependent enhancement. Virology 2006, 345, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Kietzell, K.; Pozzuto, T.; Heilbronn, R.; Gröss, L.T.; Fechner, H.; Weger, S. Antibody-Mediated Enhancement of Parvovirus B19 Uptake into Endothelial Cells Mediated by a Receptor for Complement Factor C1q. J. Virol. 2014, 88, 8102–8115. [Google Scholar] [CrossRef] [Green Version]
- Adamson-Small, L.A.; Ignatovich, I.V.; Laemmerhirt, M.G.; Hobbs, J.A. Persistent parvovirus B19 infection in non-erythroid tissues: Possible role in the inflammatory and disease process. Virus Res. 2014, 190, 8–16. [Google Scholar] [CrossRef]
- Schenk, T.; Enders, M.; Pollak, S.; Hahn, R.; Huzly, D. High Prevalence of Human Parvovirus B19 DNA in Myocardial Autopsy Samples from Subjects without Myocarditis or Dilative Cardiomyopathy. J. Clin. Microbiol. 2009, 47, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Verdonschot, J.; Hazebroek, M.; Merken, J.; Debing, Y.; Dennert, R.; Rocca, H.-P.B.-L.; Heymans, S. Relevance of cardiac parvovirus B19 in myocarditis and dilated cardiomyopathy: Review of the literature. Eur. J. Hear. Fail. 2016, 18, 1430–1441. [Google Scholar] [CrossRef]
- Cotmore, S.F.; McKie, V.C.; Anderson, L.J.; Astell, C.R.; Tattersall, P. Identification of the major structural and nonstructural proteins encoded by human parvovirus B19 and mapping of their genes by procaryotic expression of isolated genomic fragments. J. Virol. 1986, 60, 548–557. [Google Scholar] [CrossRef] [Green Version]
- Saikawa, T.; Anderson, S.; Momoeda, M.; Kajigaya, S.; Young, N.S. Neutralizing linear epitopes of B19 parvovirus cluster in the VP1 unique and VP1-VP2 junction regions. J. Virol. 1993, 67, 3004–3009. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.; Momoeda, M.; Kawase, M.; Kajigaya, S.; Young, N.S. Peptides derived from the unique region of B19 parvovirus minor capsid protein elicitneutralizing antibodies in rabbits. Virology 1995, 206, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Zuffi, E.; Manaresi, E.; Gallinella, G.; Gentilomi, G.A.; Venturoli, S.; Zerbini, M.; Musiani, M. Identification of an Immunodominant Peptide in the Parvovirus B19 VP1 Unique Region Able to Elicit a Long-Lasting Immune Response in Humans. Viral Immunol. 2001, 14, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ros, C.; Gerber, M.; Kempf, C. Conformational Changes in the VP1-Unique Region of Native Human Parvovirus B19 Lead to Exposure of Internal Sequences That Play a Role in Virus Neutralization and Infectivity. J. Virol. 2006, 80, 12017–12024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bönsch, C.; Kempf, C.; Ros, C. Interaction of Parvovirus B19 with Human Erythrocytes Alters Virus Structure and Cell Membrane Integrity. J. Virol. 2008, 82, 11784–11791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bönsch, C.; Zuercher, C.; Lieby, P.; Kempf, C.; Ros, C. The Globoside Receptor Triggers Structural Changes in the B19 Virus Capsid That Facilitate Virus Internalization. J. Virol. 2010, 84, 11737–11746. [Google Scholar] [CrossRef] [Green Version]
- Leisi, R.; Ruprecht, N.; Kempf, C.; Ros, C. Parvovirus B19 Uptake Is a Highly Selective Process Controlled by VP1u, a Novel Determinant of Viral Tropism. J. Virol. 2013, 87, 13161–13167. [Google Scholar] [CrossRef] [Green Version]
- Leisi, R.; Von Nordheim, M.; Ros, C.; Kempf, C. The VP1u Receptor Restricts Parvovirus B19 Uptake to Permissive Erythroid Cells. Viruses 2016, 8, 265. [Google Scholar] [CrossRef] [Green Version]
- Leisi, R.; Di Tommaso, C.; Kempf, C.; Ros, C. The Receptor-Binding Domain in the VP1u Region of Parvovirus B19. Viruses 2016, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, M.G.; Anderson, D.C.; Fikes, J.D.; Bain, F.T.; Carlson, C.S.; Green, S.W.; Young, N.S.; Brown, K.E. Identification of a novel simian parvovirus in cynomolgus monkeys with severe anemia. A paradigm of human B19 parvovirus infection. J. Clin. Investig. 1994, 93, 1571–1576. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.E.; Green, S.W.; O’Sullivan, M.; Young, N.S. Cloning and Sequencing of the Simian Parvovirus Genome. Virology 1995, 210, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Green, S.W.; Malkovska, I.; O’Sullivan, M.; Brown, K.E. Rhesus and Pig-Tailed Macaque Parvoviruses: Identification of Two New Members of the Erythrovirus Genus in Monkeys. Virology 2000, 269, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.E.; Young, N.S. The simian parvoviruses. Rev. Med. Virol. 1997, 7, 211–218. [Google Scholar] [CrossRef]
- O’Sullivan, M.G.; Anderson, D.K.; Lund, J.E.; Brown, W.P.; Green, S.W.; Young, N.S.; Brown, K.E. Clinical and epidemiological features of simian parvovirus infection in cynomolgus macaques with severe anemia. Lab. Anim. Sci. 1996, 46, 291–297. [Google Scholar] [PubMed]
- O’Sullivan, M.G.; Anderson, D.K.; Goodrich, J.A.; Tulli, H.; Green, S.W.; Young, N.S.; Brown, K.E. Experimental infection of cynomolgus monkeys with simian parvovirus. J. Virol. 1997, 71, 4517–4521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.E.; Liu, Z.; Gallinella, G.; Wong, S.; Mills, I.P.; O’Sullivan, M.G. Simian Parvovirus Infection: A Potential Zoonosis. J. Infect. Dis. 2004, 190, 1900–1907. [Google Scholar] [CrossRef] [PubMed]
- Gallinella, G.; Anderson, S.M.; Young, N.S.; Brown, K.E. Human parvovirus B19 can infect cynomolgus monkey marrow cells in tissue culture. J. Virol. 1995, 69, 3897–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisi, R.; Von Nordheim, M.; Kempf, C.; Ros, C. Specific Targeting of Proerythroblasts and Erythroleukemic Cells by the VP1u Region of Parvovirus B19. Bioconjugate Chem. 2015, 26, 1923–1930. [Google Scholar] [CrossRef]
- Gigler, A.; Dorsch, S.; Hemauer, A.; Williams, C.; Kim, S.; Young, N.S.; Zolla-Pazner, S.; Wolf, H.; Gorny, M.K.; Modrow, S. Generation of Neutralizing Human Monoclonal Antibodies against Parvovirus B19 Proteins. J. Virol. 1999, 73, 1974–1979. [Google Scholar] [CrossRef] [Green Version]
- Dorsch, S.; Kaufmann, B.; Schaible, U.; Prohaska, E.; Wolf, H.; Modrow, S. The VP1-unique region of parvovirus B19: Amino acid variability and antigenic stability. J. Gen. Virol. 2001, 82, 191–199. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Ab initio protein structure assembly using continuous structure fragments and optimized knowledge-based force field. Proteins: Struct. Funct. Bioinform. 2012, 80, 1715–1735. [Google Scholar] [CrossRef] [Green Version]
- Kurita, R.; Suda, N.; Sudo, K.; Miharada, K.; Hiroyama, T.; Miyoshi, H.; Tani, K.; Nakamura, Y. Establishment of Immortalized Human Erythroid Progenitor Cell Lines Able to Produce Enucleated Red Blood Cells. PLoS ONE 2013, 8, e59890. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishi, K. A new leukemia cell line with philadelphia chromosome characterized as basophil precursors. Leuk. Res. 1985, 9, 381–390. [Google Scholar] [CrossRef]
- Miyagawa, E.; Yoshida, T.; Takahashi, H.; Yamaguchi, K.; Nagano, T.; Kiriyama, Y.; Okochi, K.; Sato, H. Infection of the erythroid cell line, KU812Ep6 with human parvovirus B19 and its application to titration of B19 infectivity. J. Virol. Methods 1999, 83, 45–54. [Google Scholar] [CrossRef]
- Takahashi, T.; Ozawa, K.; Asano, S.; Takaku, F. Susceptibility of human erythropoietic cells to B19 parvovirus in vitro increases with differentiation. Blood 1990, 75, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, L.; Lyons, I.; Li, R.; Hartley, L.; Kontgen, F.; Harvey, R.; Metcalf, D.; Begley, C.G. Absence of yolk sac hematopoiesis from mice with a targeted disruption of the scl gene. Proc. Natl. Acad. Sci. USA 1995, 92, 7075–7079. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.A.; Curtis, D.; Metcalf, D.; Elefanty, A.; Sourris, K.; Robb, L.; Göthert, J.; Jane, S.M.; Begley, C.G. The critical regulator of embryonic hematopoiesis, SCL, is vital in the adult for megakaryopoiesis, erythropoiesis, and lineage choice in CFU-S12. Proc. Natl. Acad. Sci. USA 2003, 100, 992–997. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Wang, X.; Maeda, M.; Canver, M.C.; Sher, F.; Funnell, A.P.W.; Fisher, C.; Suciu, M.; Martyn, G.E.; Norton, L.J.; et al. Transcription factors LRF and BCL11A independently repress expression of fetal hemoglobin. Science 2016, 351, 285–289. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bircher, C.; Bieri, J.; Assaraf, R.; Leisi, R.; Ros, C. A Conserved Receptor-Binding Domain in the VP1u of Primate Erythroparvoviruses Determines the Marked Tropism for Erythroid Cells. Viruses 2022, 14, 420. https://doi.org/10.3390/v14020420
Bircher C, Bieri J, Assaraf R, Leisi R, Ros C. A Conserved Receptor-Binding Domain in the VP1u of Primate Erythroparvoviruses Determines the Marked Tropism for Erythroid Cells. Viruses. 2022; 14(2):420. https://doi.org/10.3390/v14020420
Chicago/Turabian StyleBircher, Cornelia, Jan Bieri, Ruben Assaraf, Remo Leisi, and Carlos Ros. 2022. "A Conserved Receptor-Binding Domain in the VP1u of Primate Erythroparvoviruses Determines the Marked Tropism for Erythroid Cells" Viruses 14, no. 2: 420. https://doi.org/10.3390/v14020420
APA StyleBircher, C., Bieri, J., Assaraf, R., Leisi, R., & Ros, C. (2022). A Conserved Receptor-Binding Domain in the VP1u of Primate Erythroparvoviruses Determines the Marked Tropism for Erythroid Cells. Viruses, 14(2), 420. https://doi.org/10.3390/v14020420