
Epidemiological and Genetic Characterization of Norovirus Outbreaks That Occurred in Catalonia, Spain, 2017–2019

 , , ,
, , , 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. HuNoV RTqPCR and Genotyping Assays
2.3. Statistical Analysis
3. Results
3.1. Epidemiological Features of HuNoV Outbreaks and Cases
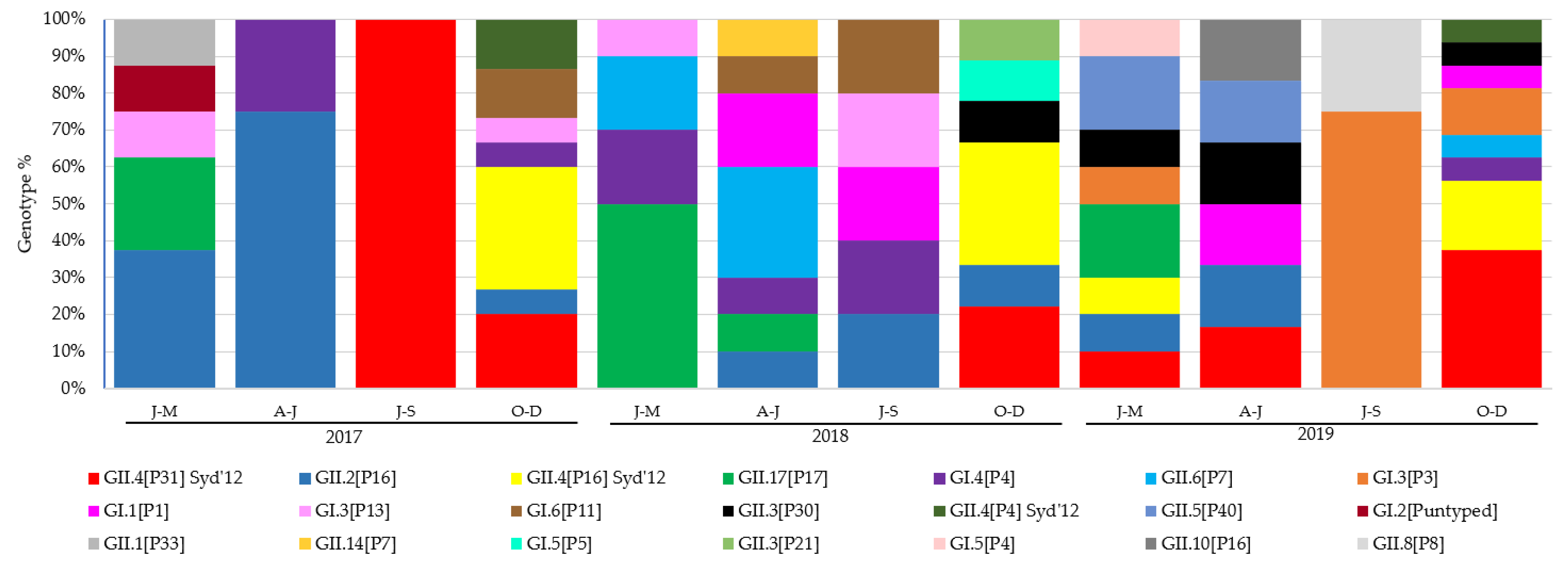
3.2. Prevalence and Evolution of HuNoV Genotypes
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, S.; Hall, A.; Robinson, A.; Verhoef, L.; Premkumar, P.; Parashar, U. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Glass, R.I.; Parashar, U.D.; Estes, M.K. Norovirus gastroenteritis. N. Engl. J. Med. 2009, 361, 1776–1785. [Google Scholar] [CrossRef] [Green Version]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Rushton, S.P.; Sanderson, R.A.; Reid, W.D.K.; Shirley, M.D.F.; Harris, J.P.; Hunter, P.R.; O’Brien, S.J. Transmission routes of rare seasonal diseases: The case of norovirus infections. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180267. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Wang, M.; Wang, K.; Estes, M.K. Sequence and genomic organization of norwalk virus. Virology 1993, 195, 51–61. [Google Scholar] [CrossRef]
- Thorne, L.G.; Goodfellow, I.G. Norovirus gene expression and replication. J. Gen. Virol. 2014, 95, 278–291. [Google Scholar] [CrossRef] [PubMed]
- White, P.A. Evolution of norovirus. Clin. Microbiol. Infect. 2014, 20, 741–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Beek, J.; de Graaf, M.; Al-Hello, H.; Allen, D.J.; Ambert-Balay, K.; Botteldoorn, N.; Brytting, M.; Buesa, J.; Cabrerizo, M.; Chan, M.C.-W.; et al. Molecular surveillance of norovirus, 2005–2016: An epidemiological analysis of data collected from the NoroNet network. Lancet Infect. Dis. 2018, 18, 545–553. [Google Scholar] [CrossRef]
- Vega, E.; Barclay, L.; Gregoricus, N.; Shirley, S.H.; Lee, D.; Vinjé, J. Genotypic and epidemiologic trends of norovirus outbreaks in the united states, 2009 to 2013. J. Clin. Microbiol. 2014, 52, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- De Graaf, M.; van Beek, J.; Vennema, H.; Podkolzin, A.T.; Hewitt, J.; Bucarado, F.; Templeton, K.; Mans, J.; Nordgren, J.; Reuter, G.; et al. Emergence of a novel GII.17 norovirus—End of the GII.4 era? Eurosurveillance 2018, 20, 21178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.C.; Brewer-Jensen, P.D.; Mallory, M.L.; Debbink, K.; Swann, E.W.; Vinjé, J.; Baric, R.S. Antigenic characterization of a novel recombinant GII.P16-GII.4 Sydney norovirus strain with minor sequence variation leading to antibody escape. J. Infect. Dis. 2018, 217, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.M.; Silva, L.D.; Sousa, E.C., Jr.; Cardoso, J.F.; Reymão, T.K.A.; Portela, A.C.R.; de Lima, C.P.S.; Teixeira, D.M.; Lucena, M.S.S.; Nunes, M.R.T.; et al. Evolutionary and molecular analysis of complete genome sequences of Norovirus from Brazil: Emerging recombinant strain GII.P16/GII.4. Front. Microbiol. 2020, 11, 1870. [Google Scholar] [CrossRef] [PubMed]
- Hasing, M.E.; Lee, B.E.; Qiu, Y.; Xia, M.; Pabbaraju, K.; Wong, A.; Tipples, G.; Jiang, X.; Pang, X.L. Changes in norovirus genotype diversity in gastroenteritis outbreaks in Alberta, Canada: 2012-2018. BMC Infect. Dis. 2019, 19, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lun, J.H.; Hewitt, J.; Yan, G.J.H.; Tuipulotu, D.E.; Rawlinson, W.D.; White, P.A. Recombinant GII.P16/GII.4 sydney 2012 was the dominant norovirus identified in Australia and New Zealand in 2017. Viruses 2018, 10, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niendorf, S.; Faber, M.; Tröger, A.; Hackler, J.; Jacobsen, S. Diversity of noroviruses throughout outbreaks in Germany 2018. Viruses 2020, 12, 1157. [Google Scholar] [CrossRef] [PubMed]
- Thanusuwannasak, T.; Puenpa, J.; Chuchaona, W.; Vongpunsawad, S.; Poovorawan, Y. Emergence of multiple norovirus strains in Thailand, 2015–2017. Infect. Genet. Evol. 2018, 61, 108–112. [Google Scholar] [CrossRef]
- Liu, L.T.; Kuo, T.; Wu, C.; Liao, W.; Hall, A.J.; Wu, F. Recombinant GII.P16-GII.2 Norovirus, Taiwan, 2016. Emerg. Infec. Dis. 2017, 23, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Niendorf, S.; Jacobsen, S.; Faber, M.; Eis-hübinger, A.M.; Hofmann, J.; Zimmermann, O.; Höhne, M.; Bock, C. Niendorf 2016 steep rise in norovirus cases and emergence of a new GII.P16_GII.2. Eurosurveillance 2017, 22, 30447. [Google Scholar] [CrossRef]
- Jin, M.; Wu, S.; Kong, X.; Xie, H.; Fu, J.; He, Y.; Feng, W.; Liu, N.; Li, J.; Rainey, J.J.; et al. Norovirus outbreak surveillance, China, 2016–2018. Emerg. Infect. Dis. 2020, 26, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Sun, L.; Fang, L.; Yang, F.; Mo, Y.; Lao, J.; Zheng, H.; Tan, X.; Lin, H.; Rutherford, S.; et al. Gastroenteritis outbreaks caused by norovirus GII.17, Guangdong Province, China, 2014–2015. Emerg. Infect. Dis. 2015, 21, 1240–1242. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.C.W.; Hu, Y.; Chen, H.; Podkolzin, A.T.; Zaytseva, E.V.; Komano, J.; Sakon, N.; Poovorawan, Y.; Vongpunsawad, S.; Thanusuwannasak, T.; et al. Global spread of norovirus GII.17 Kawasaki 308, 2014–2016. Emerg. Infect. Dis. 2017, 23, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Bidalot, M.; Théry, L.; Kaplon, J.; de Rougemont, A.; Ambert-Balay, K. Emergence of new recombinant noroviruses GII.p16- GII.4 and GII.p16-GII.2, France, winter 2016 to 2017. Eurosurveillance 2017, 22, 30508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.L.; Barclay, L.; Collins, N.R.; Wikswo, M.E.; Castro, C.J.; Magaña, C.; Gregoricus, N.; Marine, R.; Chhabra, P.; Vinjé, J. Genetic and epidemiologic trends of Norovirus outbreaks in the United States from 2013 to 2016 demonstrated emergence of novel GII.4 recombinant viruses. J. Clin. Microbiol. 2017, 55, 2208–2221. [Google Scholar] [CrossRef] [Green Version]
- Kroneman, A.; Vega, E.; Vennema, H.; Vinjé, J.; White, P.A.; Hansman, G.; Green, K.; Martella, V.; Katayama, K.; Koopmans, M. Proposal for a unified norovirus nomenclature and genotyping. Arch. Virol. 2013, 158, 2059–2068. [Google Scholar] [CrossRef] [Green Version]
- Robilotti, E.; Deresinski, S.; Pinsky, B.A. Norovirus. Clin. Microbiol. Rev. 2015, 28, 134–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, L.; Park, G.; Vega, E.; Hall, A.; Parashar, U.; Vinjé, J.; Lopman, B. Infection control for norovirus. Clin. Microbiol. Infect. 2014, 20, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Sabrià, A.; Pintó, R.M.; Bosch, A.; Bartolomé, R.; Cornejo, T.; Torner, N.; Martínez, A.; De Simón, M.; Domínguez, A.; Guix, S. Molecular and clinical epidemiology of norovirus outbreaks in Spain during the emergence of GII.4 2012 variant. J. Clin. Virol. 2014, 60, 96–104. [Google Scholar] [CrossRef]
- ISO 15216-2:2019; Microbiology of the Food Chain—Horizontal Method for Determination of Hepatitis A Virus and Norovirus Using Real-Time RT-PCR—Part 2: Method for Detection. ISO: Geneva, Switzerland, 2019.
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.; Peñaranda, S.; Oberste, M.S.; Vinje, J.; Koopmans, M. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Gallimore, C.I.; Iturriza-Gomara, M.; Xerry, J.; Adigwe, J.; Gray, J.J. Inter-seasonal diversity of norovirus genotypes: Emergence and selection of virus variants. Arch. Virol. 2007, 152, 1295–1303. [Google Scholar] [CrossRef] [PubMed]
- Chuchaona, W.; Chansaenroj, J.; Wanlapakorn, N.; Poovorawan, Y. Recombinant GII.Pe-GII.4 Norovirus, Thailand, 2017–2018. Emerg. Infect. Dis. 2019, 25, 2017–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, X.; Teng, Z.; Zhang, X. Genotypic prevalence of norovirus GII in gastroenteritis outpatients in Shanghai from 2016 to 2018. Gut Pathog. 2019, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Santiso-Bellón, C.; Randazzo, W.; Pérez-Cataluña, A.; Vila-Vicent, S.; Gozalbo-Rovira, R.; Muñoz, C.; Buesa, J.; Sanchez, G.; Díaz, J.R. Epidemiological surveillance of norovirus and rotavirus in sewage (2016–2017) in Valencia (Spain). Microorganisms 2020, 8, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barclay, L.; Cannon, J.L.; Wikswo, M.E.; Phillips, A.R.; Browne, H.; Montmayeur, A.M.; Tatusov, R.L.; Burke, R.M.; Hall, A.J.; Vinjé, J. Emerging novel GII.P16 noroviruses associated with multiple capsid genotypes. Viruses 2019, 11, 535. [Google Scholar] [CrossRef] [Green Version]
- Ruis, C.; Roy, S.; Brown, J.R.; Allen, D.J.; Goldstein, R.A.; Breuer, J. The emerging GII.P16-GII.4 Sydney 2012 norovirus lineage is circulating worldwide, arose by late-2014 and contains polymerase changes that may increase virus transmission. PLoS ONE 2017, 12, e0179572. [Google Scholar] [CrossRef] [PubMed]
- Parra, G.I.; Squires, R.B.; Karangwa, C.K.; Johnson, J.A.; Lepore, C.J.; Sosnovtsev, S.V.; Green, K.Y. Static and Evolving Norovirus Genotypes: Implications for Epidemiology and Immunity. PLoS Pathog. 2017, 13, e1006136. [Google Scholar] [CrossRef]
- Xue, L.; Cai, W.; Gao, J.; Zhang, L.; Dong, R.; Li, Y.; Wu, H.; Chen, M.; Zhang, J.; Wang, J.; et al. The resurgence of the norovirus GII.4 variant associated with sporadic gastroenteritis in the post-GII.17 period in South China, 2015 to 2017. BMC Infect. Dis. 2019, 19, 696. [Google Scholar] [CrossRef]
- Aoki, Y.; Suto, A.; Mizuta, K.; Ahiko, T.; Osaka, K.; Matsuzaki, Y. Duration of norovirus excretion and the longitudinal course of viral load in norovirus-infected elderly patients. J. Hosp. Infect. 2010, 75, 42–46. [Google Scholar] [CrossRef]
- Kroneman, A.; Verhoef, L.; Harris, J.; Vennema, H.; Duizer, E.; van Duynhoven, Y.; Gray, J.; Iturriza, M.; Böttiger, B.; Falkenhorst, G.; et al. Analysis of integrated virological and epidemiological reports of norovirus outbreaks collected within the Foodborne Viruses in Europe network from 1 July 2001 to 30 June 2006. J. Clin. Microbiol. 2008, 46, 2959–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopalan, S.; Yoshikawa, T.T. Norovirus infections in long-term care facilities. J. Am. Geriatr. Soc. 2016, 64, 1097–1103. [Google Scholar] [CrossRef]
- Mossong, J.; Hens, N.; Jit, M.; Beutels, P.; Auranen, K.; Mikolajczyk, R.; Massari, M.; Salmaso, S.; Tomba, G.S.; Wallinga, J.; et al. Social contacts and mixing patterns relevant to the spread of infectious diseases. PLoS Med. 2008, 5, e74. [Google Scholar] [CrossRef]
- Kumazaki, M.; Usuku, S. Norovirus genotype distribution in outbreaks of acute gastroenteritis among children and older people: An 8-year study. BMC Infect. Dis. 2016, 16, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, G.; Tam, C.C.; Rodrigues, L.C.; Lopman, B. Risk factors for symptomatic and asymptomatic norovirus infection in the ommunity. Epidemiol. Infect. 2011, 139, 1676–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderwood, L.E.; Wikswo, M.E.; Mattison, C.P.; Kambhampati, A.K.; Balachandran, N.; Vinjé, J.; Barclay, L.; Hall, A.J.; Parashar, U.; A Mirza, S. Norovirus outbreaks in long-term care facilities in the United States, 2009–2018: A decade of surveillance. Clin. Infect. Dis. 2022, 74, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Barret, A.S.; Silva, N.J.D.; Ambert-Balay, K.; Delmas, G.; Bone, A.; Thiolet, J.M.; Vaillant, V. Surveillance for outbreaks of gastroenteritis in elderly long-term care facilities in France, November 2010 to May 2012. Eurosurveillance 2015, 19, 20859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, M.; Fukuda, S.; Nishio, O. Statistical analysis of attack rate in norovirus foodborne outbreaks. Int. J. Food Microbiol. 2008, 122, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.; Dickey, B.; Miller, R.; Felzer, J.; Dawson, B.; See, A.; Rocks, J.J.; Kiel, J.; Montes, J.S.; Moe, C.L.; et al. The epidemiology of published norovirus outbreaks: A systematic review of risk factors associated with attack rate and genogroup. Epidemiol. Infect. 2012, 140, 1161–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, A.; Gkogka, E.; Le Guyader, F.S.; Loisy-Hamon, F.; Lee, A.; van Lieshout, L.; Marthi, B.; Myrmel, M.; Sansom, A.; Schultz, A.C.; et al. Foodborne viruses: Detection, risk assessment, and control options in food processing. Int. J. Food Microbiol. 2018, 285, 110–128. [Google Scholar] [CrossRef] [PubMed]
- Bon, F.; Ambert-Balay, K.; Giraudon, H.; Kaplon, J.; Le Guyader, S.; Pommepuy, M.; Gallay, A.; Vaillant, V.; de Valk, H.; Chikhi-Brachet, R.; et al. Molecular epidemiology of caliciviruses detected in sporadic and outbreak cases of gastroenteritis in France from December 1998 to February 2004. J. Clin. Microbiol. 2005, 43, 4659–4664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopman, B.A.; Adak, G.K.; Reacher, M.H.; Brown, D.W.G. Two epidemiologic patterns of norovirus outbreaks: Surveillance in England and Wales, 1992-2000. Emerg. Infect. Dis. 2003, 9, 71–77. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, M.; Villabruna, N.; Koopmans, M.P. Capturing norovirus transmission. Curr. Opin. Virol. 2017, 22, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Tohma, K.; Lepore, C.J.; Ford-Siltz, L.A.; Parra, G.I. Phylogenetic analyses suggest that factors other than the capsid protein play a role in the epidemic potential of GII.2 Norovirus. MSphere 2017, 2, e00187-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode of Transmission | Total Outbreaks | Duration of the Outbreak (Days) a | Total Number of Affected Individuals | Mean of Affected Individuals per Outbreak | Average Attack Rate (%) a |
|---|---|---|---|---|---|
| P/P | 74 | 8.93 ± 6.31 b | 1795 c | 29.42 ± 18.13 c | 29.71 ± 20.85 c |
| Foodborne | 22 | 3.20 ± 2.54 d | 696 e | 38.66 ± 65.13 e | 45.76 ± 26.66 e |
| Foodborne + P/P | 3 | 4.33 ± 4.16 | 145 | 48.33 ± 32 | 48.12 ± 18.96 |
| Waterborne | 1 | 4 | 41 | 41 | 64 |
| Setting | Total Outbreaks | Mode of Transmission | Season | Duration of the Outbreak (Days) a | Total Number of Affected Individuals | Average Attack Rate (%) a |
|---|---|---|---|---|---|---|
| Nursing home | 48 | P/P (43), Foodborne (4), Foodborne + P/P (1) | Cold (42), warm (6) | 8.86 ± 5.89 b | 1208 c | 28.62 ± 20.03 c |
| Youth hostel/Campground | 20 | Foodborne (12), P/P (5), Foodborne + P/P (2), Waterborne (1) | Warm (12), cold (8) | 3.18 ± 2.86 d | 537 e | 49.22 ± 22.42 e |
| School | 13 | P/P (11), Foodborne (2) | Cold (8), warm (5) | 9.55 ± 6.25 f | 637 f | 32.61 ± 23.21 f |
| Kindergarten/Preschool | 6 | P/P (6) | Cold (5), warm (1) | 12.50 ± 9.09 | 40 g | 23.43 ± 6.16 g |
| Hotel | 6 | Foodborne (4), P/P (2) | Cold (3), warm (3) | 3.40 ± 3.71 h | 113 h | 39.12 ± 35.86 h |
| Long-term care facility | 6 | P/P (6) | Warm (4), cold (2) | 9.33 ± 2.08 g | 131 i | 35.13 ± 29.17 i |
| Hospital | 1 | P/P (1) | Warm (1) | 3 | 11 | 17 |
| <15 Years | 16–65 Years | >65 Years | |
|---|---|---|---|
| Symptom | n (%) | n (%) | n (%) |
| Diarrhea | 123 (61.5) | 113 (83.1) | 147 (96.7) |
| Vomiting | 196 (91.6) | 121 (88.3) | 110 (70.1) |
| Fever | 75 (39.9) | 53 (47.3) | 7 (0.1) |
| Genotype | Total Outbreaks | Setting (Number of Outbreaks) | Mode of Transmission | Season | Mean Age of Cases (Years) | Duration of Outbreak (Days) | Total Number of Affected Individuals | Average Attack Rate (%) a |
|---|---|---|---|---|---|---|---|---|
| GI.1[P1] | 5 | Long-term care facility (2), Youth hostel/Campground (2), Hotel (1) | P/P (3), Foodborne (2) | Warm (4), Cold (1) | 36.66 ± 20.56 | 6 ± 5.23 b | 107 b | 28.35 ± 30.84 b |
| GI.3[P3] | 6 | Youth hostel/Campground (4), Nursing home (2) | P/P (3), Foodborne (3) | Warm (3), Cold (3) | 46.71 ± 33.12 b | 7.67 ± 4.62 c | 189 | 36.93 ± 17.96 |
| GI.3[P13] | 2 | Youth hostel/Campground (1), School (1) | P/P (1), Foodborne (1) | Cold (1), Warm (1) | 19.48 ± 11.72 | 5.5 ± 3.54 | 45 | 48 ± 1.41 |
| GI.4[P4] | 6 | Nursing home (3), School (2), Hotel (1) | P/P (6) | Cold (3), Warm (3) | 43.08 ± 29.89 | 9.4 ± 8.52 b | 115 b | 18.23 ± 17.68 b |
| GI.5[P4] | 1 | Nursing home (1) | P/P (1) | Cold (1) | 62.8 | 2 | 18 | 11.69 |
| GI.5[P5] | 1 | Nursing home (1) | P/P (1) | Cold (1) | 71.31 | 7 | 28 | 17.18 |
| GI.6[P11] | 3 | Nursing home (2), Youth hostel/Campground (1) | P/P (3) | Warm (2), Cold (1) | 52.3 ± 31.74 | 7 ± 5.29 | 135 | 36.34 ± 8.47 |
| Multiple GI genotypes k | 1 | Nursing home (1) | P/P (1) | Cold (1) | 60.72 | 17 | 59 | 83.1 |
| Total GI | 25 | Nursing home (10), Youth hostel/Campground (8), School (3), Long-term care facility (2), Hotel (2) | P/P (19), Foodborne (6) | Cold (13), Warm (12) | 41.85 ± 26.8 b | 7.73 ± 6.13 d | 721 e | 30.71 ± 21.93 e |
| GII.1[P33] | 1 | School (1) | P/P (1) | Cold (1) | 9.81 | 9 | 45 | 12.47 |
| GII.2[P16] | 11 | Youth hostel/Campground (6), Nursing home (3), School (1), Hotel (1) | P/P (5), Foodborne (3), Foodborne+ P/P (2), Waterborne (1) | Warm (6), Cold (5) | 34.73 ± 25.26 | 4.82 ± 4.51 | 434 | 55.56 ± 24.03 |
| GII.3[P21] | 1 | School (1) | P/P (1) | Cold (1) | 18 | ND | ND | ND |
| GII.3[P30] | 3 | School (1), Nursing home (1), Hotel (1) | P/P (2), Foodborne (1) | Cold (2), Warm (1) | 51.77 ± 27.94 | 10.5 ± 0.71 b | 45 b | 42.17 ± 46.43 b |
| GII.4[P4] Sydney 2012 | 2 | Nursing home (2) | P/P (2) | Cold (2) | 94.6 b | 12 b | 32 b | 19.75 b |
| GII.4[P16] Sydney 2012 | 11 | Nursing home (7) Kindergarten/Preschool (3), Long-term care facility (1) | P/P (11) | Cold (11) | 59.38 ± 37.73 b | 12.37 ± 8.49 c | 255 f | 37.52 ± 14.4 f |
| GII.4[P31] Sydney 2012 | 14 | Nursing home (11), Kindergarten/Preschool (1), Long-term care facility (1), Hotel (1) | P/P (11), Foodborne (3) | Cold (12), Warm (2) | 64.02 ± 24.81 d | 6.89 ± 6.15 d | 206 g | 21.24 ± 18.23 g |
| GII.5[P40] | 3 | Nursing home (1), Youth hostel/Campground (1), School (1) | Foodborne (2), P/P (1) | Cold (2), Warm (1) | 43.15 ± 14.06 | 4.33 ± 2.31 | 87 | 43.36 ± 28 |
| GII.6[P7] | 6 | Nursing home (2), Youth hostel/Campground (2), School (1), Hotel (1) | P/P (3), Foodborne (2), Foodborne+ P/P (1) | Warm (3), Cold (3) | 35.18 ± 22.29 | 5.33 ± 4.41 | 153 b | 51.24 ± 27.93 b |
| GII.8[P8] | 1 | Youth hostel/Campground (1) | Foodborne (1) | Warm (1) | 15.6 | ND | 20 | 20.41 |
| GII.10[P16] | 1 | Socio-health centre (1) | P/P (1) | Warm (1) | 62 | 10 | 32 | 17.58 |
| GII.14[P7] | 1 | School (1) | P/P (1) | Warm (1) | 18.69 | ND | 11 | 42.31 |
| GII.17[P17] | 9 | Nursing home (6), Youth hostel/Campground (1), School (1) Long-term care facility (1) | P/P (7), Foodborne (2) | Cold (8), Warm (1) | 58.89 ± 24.75 | 6.29 ± 3.55 e | 466 | 32.06 ± 20.8 |
| Multiple GII genotypes l | 1 | School (1) | P/P (1) | Cold (1) | 18.82 | 12 | 55 | 11.58 |
| Total GII | 65 | Nursing home (33), Youth hostel/Campground (11), School (9), Kindergarten/Preschool (4), Hotel (4), Long-term care facility (3), Hospital (1) | P/P (47), Foodborne (14), Foodborne + P/P (3), Waterborne (1) | Cold (48), Warm (17) | 48 ± 31.05 h | 7.56 ± 6.22 i | 1905 j | 37.23 ± 23.4 j |
| Multiple GI + GII genotypes m | 3 | Nursing home (2), Kindergarten/Preschool (1) | P/P (2), Foodborne (1) | Cold (3) | 49.46 ± 37.1 | 7 ± 5.66 b | 51 | 13.17 ± 11.26 |
| Total | 93 | Nursing home (45), Youth hostel/Campground (19), School (12), Kindergarten/Preschool (5), Hotel (6), Long-term care facility (5), Hospital (1) | P/P (68), Foodborne (21), Foodborne + P/P (3), Waterborne (1) | Cold (64), Warm (29) | 46.31 ± 30.03 | 7.59 ± 6.12 | 2677 | 34.22 ± 23.14 |
| 1608 | 1609 | 1610 | 1611 | 1612 | 1613 | 1614 | 1615 | 1616 | 1617 | 1618 | 1619 | 1620 | 1621 | 1622 | 1623 | 1624 | 1625 | 1626 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GI.P1(QCT04921) | E | R | Q | I | F | W | T | R | G | P | N | H | S | D | P | S | E | T | L |
| VV165.19 | E | R | Q | I | F | W | T | R | G | S | N | H | S | D | P | S | E | T | L |
| 1760 | 1761 | 1762 | 1763 | 1764 | 1765 | 1766 | 1767 | 1768 | 1769 | 1770 | 1771 | 1772 | 1773 | 1774 | 1775 | 1776 | 1777 | 1778 | |
| GI.P11(QJC14597) | M | F | R | W | M | R | F | H | D | L | G | L | W | T | G | D | R | N | L |
| RSBS87.17 | M | F | R | W | M | R | F | H | D | F | G | L | W | T | G | D | R | N | L |
| 1629 | 1630 | 1631 | 1632 | 1633 | 1634 | 1635 | 1636 | 1637 | 1638 | 1639 | 1640 | 1641 | 1642 | 1643 | 1644 | 1645 | 1646 | 1647 | |
| GII.P16 (QCE20812) | L | M | A | L | L | G | E | A | S | L | H | G | P | S | F | Y | S | K | I |
| RSBS74.17 | L | M | A | L | L | G | E | A | S | R | H | G | P | S | F | Y | S | K | I |
| 1635 | 1636 | 1637 | 1638 | 1639 | 1640 | 1641 | 1642 | 1643 | 1644 | 1645 | 1646 | 1647 | 1648 | 1649 | 1650 | 1651 | 1652 | 1653 | |
| GII.P31(QCQ77492) | I | V | S | T | D | I | K | L | D | P | E | K | L | T | A | K | L | K | E |
| ASPB140.17 | I | V | S | T | D | I | K | L | D | T | E | R | L | T | A | K | L | K | E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anfruns-Estrada, E.; Sabaté, S.; Razquin, E.; Cornejo Sánchez, T.; Bartolomé, R.; Torner, N.; Izquierdo, C.; Soldevila, N.; Coronas, L.; Domínguez, À.; et al. Epidemiological and Genetic Characterization of Norovirus Outbreaks That Occurred in Catalonia, Spain, 2017–2019. Viruses 2022, 14, 488. https://doi.org/10.3390/v14030488
Anfruns-Estrada E, Sabaté S, Razquin E, Cornejo Sánchez T, Bartolomé R, Torner N, Izquierdo C, Soldevila N, Coronas L, Domínguez À, et al. Epidemiological and Genetic Characterization of Norovirus Outbreaks That Occurred in Catalonia, Spain, 2017–2019. Viruses. 2022; 14(3):488. https://doi.org/10.3390/v14030488
Chicago/Turabian StyleAnfruns-Estrada, Eduard, Sara Sabaté, Efrén Razquin, Thais Cornejo Sánchez, Rosa Bartolomé, Nuria Torner, Conchita Izquierdo, Nuria Soldevila, Lorena Coronas, Àngela Domínguez, and et al. 2022. "Epidemiological and Genetic Characterization of Norovirus Outbreaks That Occurred in Catalonia, Spain, 2017–2019" Viruses 14, no. 3: 488. https://doi.org/10.3390/v14030488
APA StyleAnfruns-Estrada, E., Sabaté, S., Razquin, E., Cornejo Sánchez, T., Bartolomé, R., Torner, N., Izquierdo, C., Soldevila, N., Coronas, L., Domínguez, À., Fuentes, C., Pintó, R. M., Bosch, A., Guix, S., & The Working Group for the Study of Outbreaks of Acute Gastroenteritis in Catalonia (PI16/02005). (2022). Epidemiological and Genetic Characterization of Norovirus Outbreaks That Occurred in Catalonia, Spain, 2017–2019. Viruses, 14(3), 488. https://doi.org/10.3390/v14030488