
Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
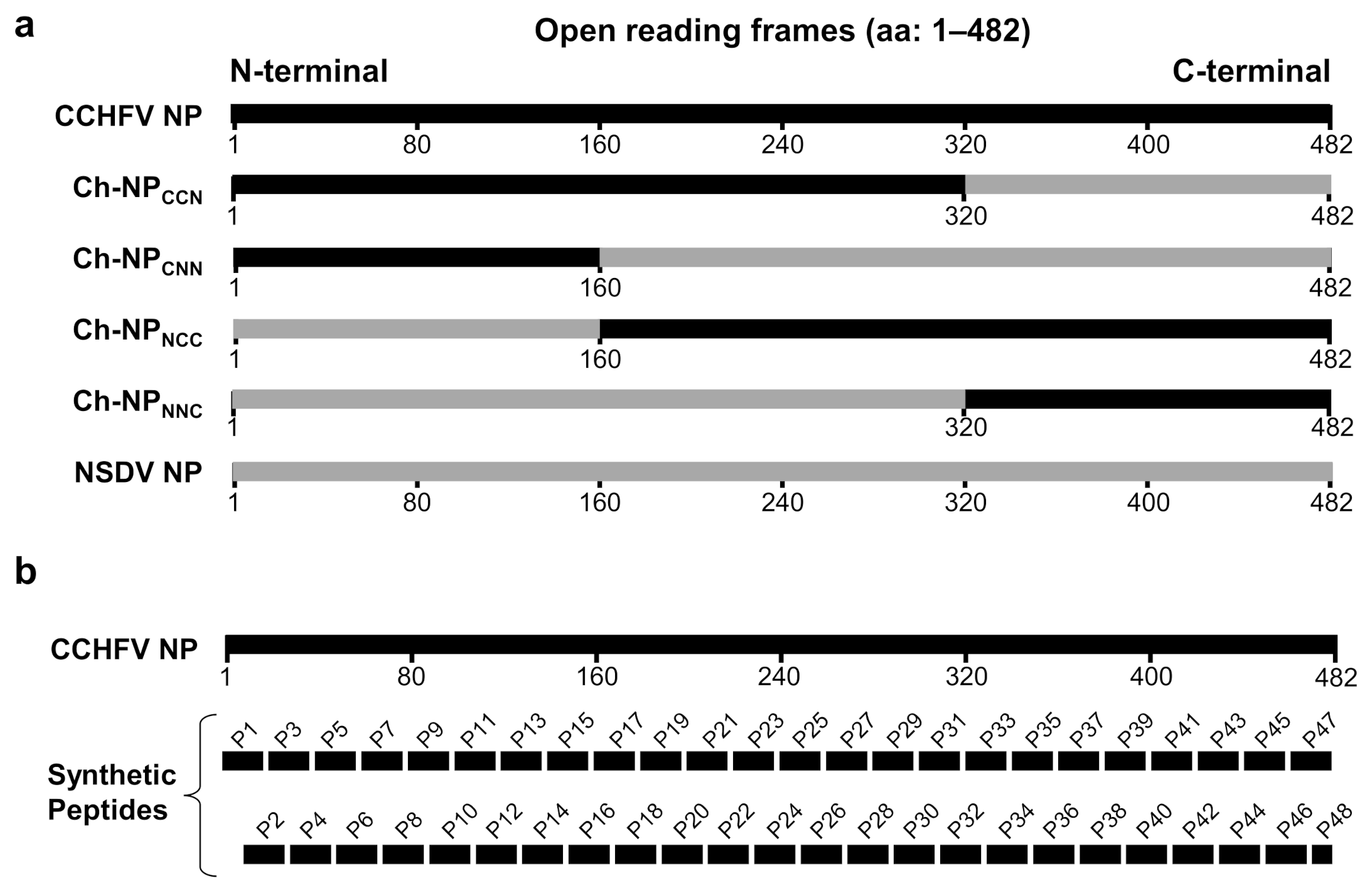
2.1. Construction of Plasmids Expressing CCHFV and NSDV NPs
2.2. Expression and Purification of NPs
2.3. Serum and Mouse Immune Ascites Fluids (MIAFs) Samples
2.4. Production and Characterization of Mouse mAbs to CCHFV NP
2.5. ELISA for mAb Screening
2.6. Western Blotting
2.7. ELISA with Synthetic Peptides as Antigens
2.8. Structural Analysis
2.9. Amino Acid Sequence Analysis of Cross-Reactive Peptides
2.10. Statistical Analysis
2.11. Ethics Statement
3. Results
3.1. Reactivity of Antisera/MIAF and CCHFV-Infected Monkey/Human Sera to CCHFV and NSDV Chimeric NPs in Western Blotting
3.2. Reactivity of Anti-NP mAbs in Western Blotting
3.3. Identification of Epitope Peptides on the CCHFV NP Molecule in ELISA
3.4. Amino Acid Sequence Comparison for the Common Epitope Peptides
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fillâtre, P.; Revest, M.; Tattevin, P. Crimean-Congo hemorrhagic fever: An update. Med. Mal. Infect. 2019, 49, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Raabe, V.N. Diagnostic testing for Crimean-Congo hemorrhagic fever. J. Clin. Microbiol. 2020, 58, e01580-19. [Google Scholar] [CrossRef]
- Bente, D.A.; Forrester, N.L.; Watts, D.M.; McAuley, A.J.; Whitehouse, C.A.; Bray, M. Crimean-Congo hemorrhagic fever: History, epidemiology, pathogenesis, clinical syndrome and genetic diversity. Antivir. Res. 2013, 100, 159–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehand, M.; Millett, S.P.; Al-Shorbaji, F.; Roth, C.; Kieny, M.P.; Murgue, B. World Health Organization methodology to prioritize emerging infectious diseases in need of research and development. Emerg. Infect. Dis. 2018, 24, e171427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, L.T.; Kelly-Cirino, C. Diagnostic tests for Crimean-Congo haemorrhagic fever: A widespread tickborne disease. BMJ Glob. Health 2019, 4, e001114. [Google Scholar] [CrossRef] [Green Version]
- Mattiuzzo, G.; Bentley, E.M.; Page, M. The role of reference materials in the research and development of diagnostic tools and treatments for haemorrhagic fever viruses. Viruses 2019, 11, 781. [Google Scholar] [CrossRef] [Green Version]
- Spengler, J.; Bergeron, R.É.; Spiropoulou, C.F. Crimean-Congo hemorrhagic fever and expansion from endemic regions. Curr. Opin. Virol. 2019, 34, 70–78. [Google Scholar] [CrossRef]
- Shrivastava, N.; Shrivastava, A.; Ninawe, S.M.; Sharma, S.; Kumar, J.S.; Alam, S.I.; Kanani, A.; Sharma, S.K.; Dash, P.K. Development of multispecies recombinant nucleoprotein-based indirect ELISA for high-throughput screening of Crimean-Congo hemorrhagic fever virus-specific antibodies. Front. Microbiol. 2019, 10, 1822. [Google Scholar] [CrossRef]
- Liu, D.; Li, Y.; Zhao, J.; Deng, F.; Duan, X.; Kou, C.; Wu, T.; Li, Y.; Wang, Y.; Ma, J.; et al. Fine epitope mapping of the central immunodominant region of nucleoprotein from Crimean-Congo hemorrhagic fever virus (CCHFV). PLoS ONE 2014, 9, e108419. [Google Scholar] [CrossRef]
- Saijo, M.; Qing, T.; Niikura, M.; Maeda, A.; Ikegami, T.; Prehaud, C.; Kurane, I.; Morikawa, S. Recombinant nucleoprotein-based enzyme-linked immunosorbent assay for detection of immunoglobulin G antibodies to Crimean-Congo hemorrhagic fever virus. J. Clin. Microbiol. 2002, 40, 1587–1591. [Google Scholar] [CrossRef] [Green Version]
- Saijo, M.; Qing, T.; Niikura, M.; Maeda, A.; Ikegami, T.; Sakai, K.; Prehaud, C.; Kurane, I.; Morikawa, S. Immunofluorescence technique using HeLa cells expressing recombinant nucleoprotein for detection of immunoglobulin G antibodies to Crimean-Congo hemorrhagic fever virus. J. Clin. Microbiol. 2002, 40, 372–375. [Google Scholar] [CrossRef] [Green Version]
- Garcia, S.; Chinikar, S.; Coudrier, D.; Billecocq, A.; Hooshmand, B.; Crance, J.M.; Garin, D.; Bouloy, M. Evaluation of a Crimean-Congo hemorrhagic fever virus recombinant antigen expressed by Semliki Forest suicide virus for IgM and IgG antibody detection in human and animal sera collected in Iran. J. Clin. Virol. 2006, 35, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Samudzi, R.R.; Leman, P.A.; Paweska, J.T.; Swanepoel, R.; Burt, F.J. Bacterial expression of Crimean-Congo hemorrhagic fever virus nucleoprotein and its evaluation as a diagnostic reagent in an indirect ELISA. J. Virol. Methods 2012, 179, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.; Burt, F.; Rybicki, E.P.; Meyers, A.E. Plant-produced Crimean-Congo haemorrhagic fever virus nucleoprotein for use in indirect ELISA. J. Virol. Methods 2016, 236, 170–177. [Google Scholar] [CrossRef]
- Emmerich, P.; Mika, A.; von Possel, R.; Rackow, A.; Liu, Y.; Schmitz, H.; Gunther, S.; Sherifi, K.; Halili, B.; Jakupi, X.; et al. Sensitive and specific detection of Crimean-Congo hemorrhagic fever virus (CCHFV)-specific IgM and IgG antibodies in human sera using recombinant CCHFV nucleoprotein as antigen in mu-capture and IgG immune complex (IC) ELISA tests. PLoS Negl. Trop. Dis. 2018, 12, e0006366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombe, B.P.; Miyamoto, H.; Saito, T.; Yoshida, R.; Manzoor, R.; Kajihara, M.; Shimojima, M.; Fukushi, S.; Morikawa, S.; Yoshikawa, T.; et al. Purification of Crimean–Congo hemorrhagic fever virus nucleoprotein and its utility for serological diagnosis. Sci. Rep. 2021, 11, 2324. [Google Scholar] [CrossRef] [PubMed]
- Leblebicioglu, H.; Ozaras, R.; Irmak, H.; Sencan, I. Crimean-Congo hemorrhagic fever in Turkey: Current status and future challenges. Antivir. Res. 2016, 126, 21–34. [Google Scholar] [CrossRef]
- Krasteva, S.; Jara, M.; Frias-De-Diego, A.; Machado, G. Nairobi sheep disease virus: A historical and epidemiological perspective. Front. Vet. Sci. 2020, 7, 419. [Google Scholar] [CrossRef] [PubMed]
- Hartlaub, J.; Keller, M.; Groschup, M.H. Deciphering antibody responses to orthonairoviruses in ruminants. Microorganisms 2021, 9, 1493. [Google Scholar] [CrossRef]
- Kajihara, M.; Simuunza, M.; Saasa, N.; Dautu, G.; Mori-Kajihara, A.; Qiu, Y.; Nakao, R.; Eto, Y.; Furumoto, H.; Hang’ombe, B.M.; et al. Serologic and molecular evidence for circulation of Crimean-Congo hemorrhagic fever virus in ticks and cattle in Zambia. PLoS Negl. Trop. Dis. 2021, 15, e0009452. [Google Scholar] [CrossRef]
- Pantarotto, D.; Partidos, C.D.; Graff, R.; Hoebeke, J.; Briand, J.P.; Prato, M.; Bianco, A. Synthesis, structural characterization, and immunological properties of carbon nanotubes functionalized with peptides. J. Am. Chem. Soc. 2003, 125, 6160–6164. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, J.M.; Roitburd-Berman, A.; Siman-Tov, D.D.; Tarnovitski Freund, N.; Weiss, Y. Epitope mapping: The first step in developing epitope-based vaccines. BioDrugs 2007, 21, 145–156. [Google Scholar] [CrossRef]
- Changula, K.; Yoshida, R.; Noyori, O.; Marzi, A.; Miyamoto, H.; Ishijima, M.; Yokoyama, A.; Kajihara, M.; Feldmann, H.; Mweene, A.S.; et al. Mapping of conserved and species-specific antibody epitopes on the Ebola virus nucleoprotein. Virus Res. 2013, 176, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prickett, S.R.; Rolland, J.M.; O’Hehir, R.E. Immunoregulatory T cell epitope peptides: The new frontier in allergy therapy. Clin. Exp. Allergy 2015, 45, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.F.; Luo, Y.J.; Li, T.X.; Wang, H.L.; Hu, Z.H.; Zhang, F.C.; Zhang, Y.J.; Deng, F.; Sun, S.R. Serial expression of the truncated fragments of the nucleocapsid protein of CCHFV and identification of the epitope region. Virol. Sin. 2010, 25, 45–51. [Google Scholar] [CrossRef]
- Burt, F.J.; Samudzi, R.R.; Randall, C.; Pieters, D.; Vermeulen, J.; Knox, C.M. Human defined antigenic region on the nucleoprotein of Crimean-Congo hemorrhagic fever virus identified using truncated proteins and a bioinformatics approach. J. Virol. Methods 2013, 193, 706–712. [Google Scholar] [CrossRef]
- Shahhosseini, S.; Das, D.; Qiu, X.; Feldmann, H.; Jones, S.M.; Suresh, M.R. Production and characterization of monoclonal antibodies against different epitopes of Ebola virus antigens. J. Virol. Methods 2007, 143, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.E.; Hammarlund, E.; Slifka, M.K. Optimization of peptide-based ELISA for serological diagnostics: A retrospective study of human monkeypox infection. Vector Borne Zoonotic Dis. 2012, 12, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Moming, A.; Tuoken, D.; Yue, X.; Xu, W.; Guo, R.; Liu, D.; Li, Y.; Hu, Z.; Deng, F.; Zhang, Y.; et al. Mapping of B-cell epitopes on the N-terminal and C-terminal segment of nucleocapsid protein from Crimean-Congo hemorrhagic fever virus. PLoS ONE 2018, 13, e0204264. [Google Scholar] [CrossRef]
- Marriott, A.C.; Polyzoni, T.; Antoniadis, A.; Nuttall, P.A. Detection of human antibodies to Crimean-Congo haemorrhagic fever virus using expressed viral nucleocapsid protein. J. Gen. Virol. 1994, 75, 2157–2161. [Google Scholar] [CrossRef]
- Pellequer, J.L.; Westhof, E.; Van Regenmortel, M.H. Predicting location of continuous epitopes in proteins from their primary structures. Methods Enzymol. 1991, 203, 176–201. [Google Scholar] [PubMed]
- Walker, P.J.; Widen, S.G.; Wood, T.G.; Guzman, H.; Tesh, R.B.; Vasilakis, N. A global genomic characterization of nairoviruses identifies nine discrete genogroups with distinctive structural characteristics and host-vector associations. Am. J. Trop. Med. Hyg. 2016, 94, 1107–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, A.; Ma, B.; Kouidou, S.; Tang, Q.; Hang, C.; Antoniadis, A. Genetic characterization of the M RNA segment of Crimean Congo hemorrhagic fever virus strains, China. Emerg. Infect. Dis. 2002, 8, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Honig, J.E.; Osborne, J.C.; Nichol, S.T. The high genetic variation of viruses of the genus Nairovirus reflects the diversity of their predominant tick hosts. Virology 2004, 318, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casals, J.; Tignor, G.H. The Nairovirus genus: Serological relationships. Intervirology 1980, 14, 144–147. [Google Scholar] [CrossRef] [PubMed]
- Kalkan-Yazıcı, M.; Karaaslan, E.; Çetin, N.S.; Hasanoğlu, S.; Güney, F.; Zeybek, Ü.; Doymaz, M.Z. Cross-reactive anti-nucleocapsid protein immunity against Crimean-Congo hemorrhagic fever virus and Hazara virus in multiple species. J. Virol. 2021, 95, e02156-20. [Google Scholar] [CrossRef]
- Crabtree, M.B.; Sang, R.; Miller, B.R. Kupe virus, a new virus in the family bunyaviridae, genus nairovirus, Kenya. Emerg. Infect. Dis. 2009, 15, 147–154. [Google Scholar] [CrossRef]
- Ergünay, K.; Dinçer, E.; Kar, S.; Emanet, N.; Yalçınkaya, D.; Polat Dinçer, P.F.; Brinkmann, A.; Hacıoğlu, S.; Nitsche, A.; Özkul, A.; et al. Multiple orthonairoviruses including Crimean-Congo hemorrhagic fever virus, Tamdy virus and the novel Meram virus in Anatolia. Ticks Tick Borne Dis. 2020, 11, 101448. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Wiley, M.R.; Rodriguez, S.E.; Bào, Y.; Prieto, K.; Travassos da Rosa, A.P.; Guzman, H.; Savji, N.; Ladner, J.T.; Tesh, R.B.; et al. Genomic characterization of the genus Nairovirus (Family Bunyaviridae). Viruses 2016, 8, 164. [Google Scholar] [CrossRef]
- Kodama, F.; Yamaguchi, H.; Park, E.; Tatemoto, K.; Sashika, M.; Nakao, R.; Terauchi, Y.; Mizuma, K.; Orba, Y.; Kariwa, H.; et al. A novel nairovirus associated with acute febrile illness in Hokkaido, Japan. Nat. Commun. 2021, 12, 5539. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum and MIAF | CCHFV NP | Ch-NP | NSDV NP | |||
|---|---|---|---|---|---|---|
| CCN | CNN | NCC | NNC | |||
| Rabbit anti-CCHFV NP | + | + | − | + | − | − |
| Mouse anti-CCHFV NP | + | + | − | + | − | − |
| Mouse anti-NSDV | − | − | + | − | + | + |
| Mouse anti-DUGV | ± c | − | + | − | + | + |
| CCHFV-infected monkey | + | ND b | ND | ND | ND | − |
| CCHFV-infected patient #A | + | + | − | + | − | − |
| CCHFV-infected patient #B | + | + | − | + | − | − |
| CCHFV-infected patient #C | + | − | ± c | − | − | − |
| CCHFV-infected patient #D | + | + | − | + | − | − |
| CCHFV-infected patient #E | + | + | − | + | − | − |
| Group | mAb | ELISA (CCHFV NP) b | CCHFV NP a | Ch-NP a | NSDV NP a | Isotype | |||
|---|---|---|---|---|---|---|---|---|---|
| CCN | CNN | NCC | NNC | ||||||
| I | 09-2 | + | − | − | − | − | − | − | IgG2b kappa |
| 27-6 | + | − | − | − | − | − | − | IgG2b kappa | |
| 87-5 | + | − | − | − | − | − | − | IgG2b kappa | |
| II | 32-1 | + | + | − | − | − | − | − | IgG2b kappa |
| 74-2 | + | + | − | − | − | − | − | IgG2b kappa | |
| 86-3 | + | + | − | − | − | − | − | IgG2b kappa | |
| 114-2 | + | + | − | − | − | − | − | IgG2b kappa | |
| III | 17-3 | + | + | + | − | + | − | − | IgG2b kappa |
| 79-10 | + | + | + | − | + | − | − | IgG2a kappa | |
| 80-6 | + | + | + | − | + | − | − | IgG2a kappa | |
| 91-5 | + | + | + | − | + | − | − | IgG2a kappa | |
| 97-6 | + | + | + | − | + | − | − | IgG2a kappa | |
| Peptide Name (Amino Acid Position) | CCHFV-Infected Patient | CCHFV-Infected Monkey | MIAF | |||||
|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | NSDV | DUGV | ||
| P9 (81–100) | − | − | − | − | − | − | − | + |
| P14 (131–150) | + | + | − | + | − | + | + | − |
| P19 (181–200) | − | − | − | − | + | − | − | − |
| P22 (211–230) | + | − | − | + | − | + | + | + |
| P30 (291–310) | − | − | − | − | − | + | − | − |
| P34 (331–350) | − | − | − | + | − | − | − | − |
| P35 (341–360) | + | − | − | + | − | − | − | − |
| P42 (411–430) | + | − | − | − | + | − | − | − |
| P45 (441–460) | − | − | + | − | − | − | − | − |
| P46 (451–470) | − | − | + | − | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombe, B.P.; Saito, T.; Miyamoto, H.; Mori-Kajihara, A.; Kajihara, M.; Saijo, M.; Masumu, J.; Hattori, T.; Igarashi, M.; Takada, A. Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein. Viruses 2022, 14, 544. https://doi.org/10.3390/v14030544
Lombe BP, Saito T, Miyamoto H, Mori-Kajihara A, Kajihara M, Saijo M, Masumu J, Hattori T, Igarashi M, Takada A. Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein. Viruses. 2022; 14(3):544. https://doi.org/10.3390/v14030544
Chicago/Turabian StyleLombe, Boniface Pongombo, Takeshi Saito, Hiroko Miyamoto, Akina Mori-Kajihara, Masahiro Kajihara, Masayuki Saijo, Justin Masumu, Takanari Hattori, Manabu Igarashi, and Ayato Takada. 2022. "Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein" Viruses 14, no. 3: 544. https://doi.org/10.3390/v14030544
APA StyleLombe, B. P., Saito, T., Miyamoto, H., Mori-Kajihara, A., Kajihara, M., Saijo, M., Masumu, J., Hattori, T., Igarashi, M., & Takada, A. (2022). Mapping of Antibody Epitopes on the Crimean-Congo Hemorrhagic Fever Virus Nucleoprotein. Viruses, 14(3), 544. https://doi.org/10.3390/v14030544