
Human Brain Organoids as Models for Central Nervous System Viral Infection

 , , and
, , and
Abstract
:1. Introduction
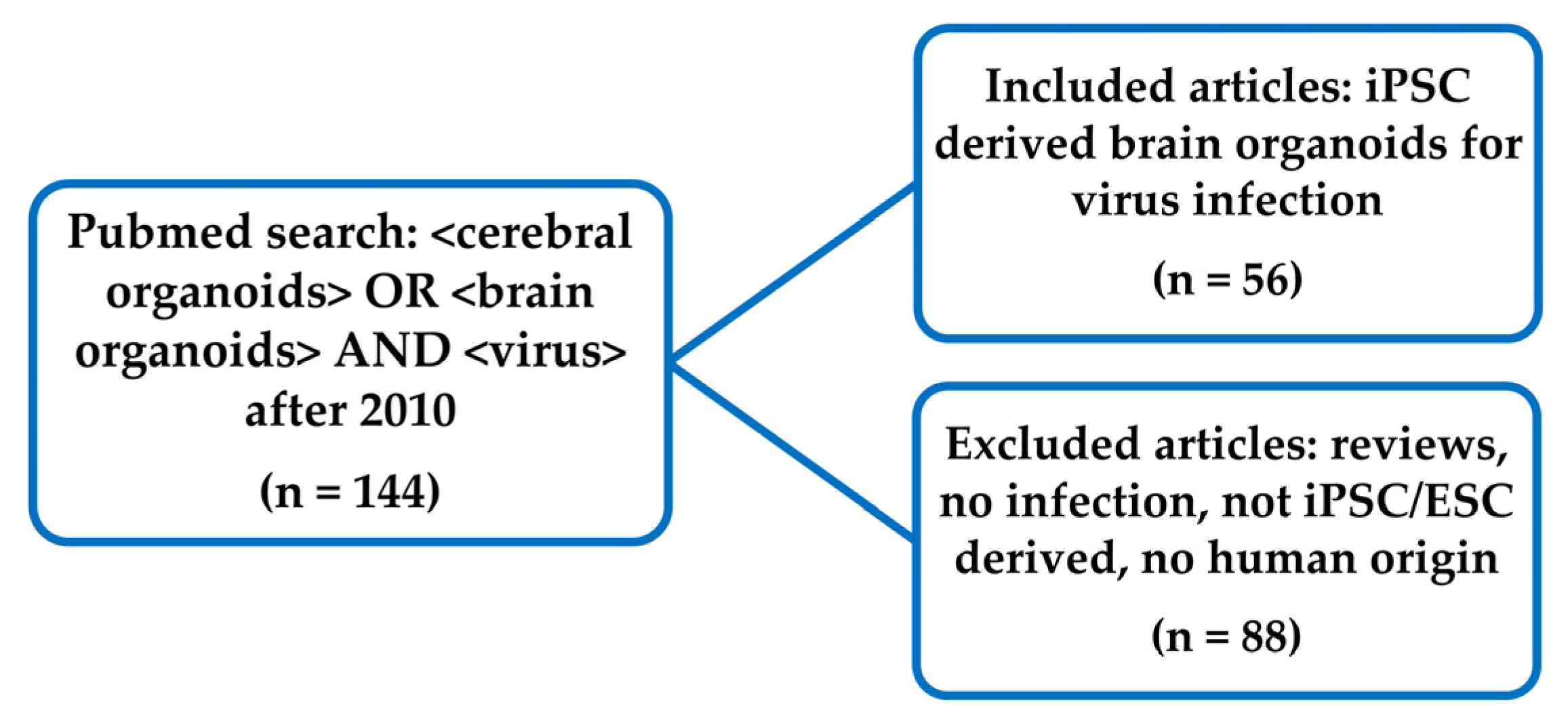
2. Methods
3. Results
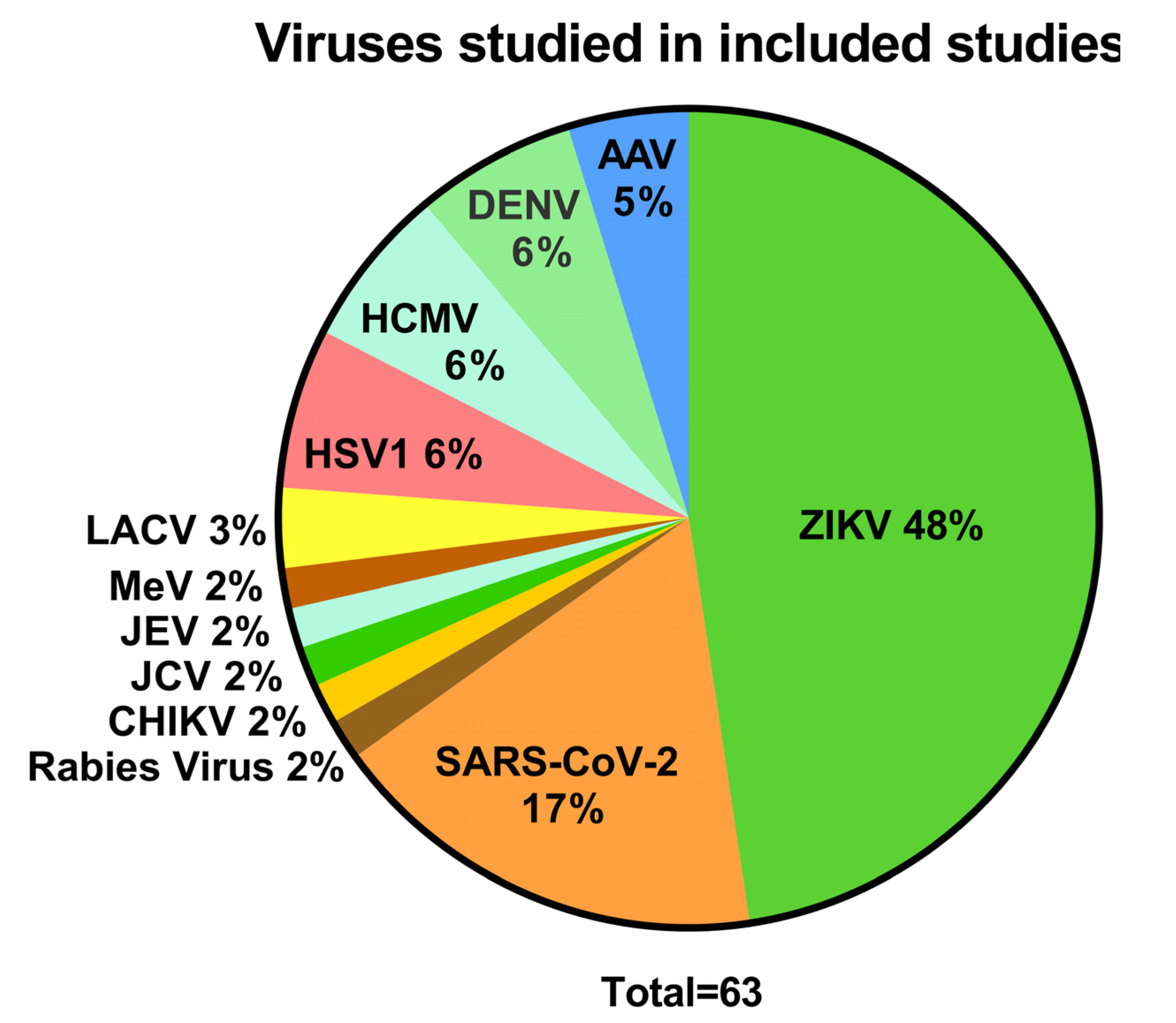
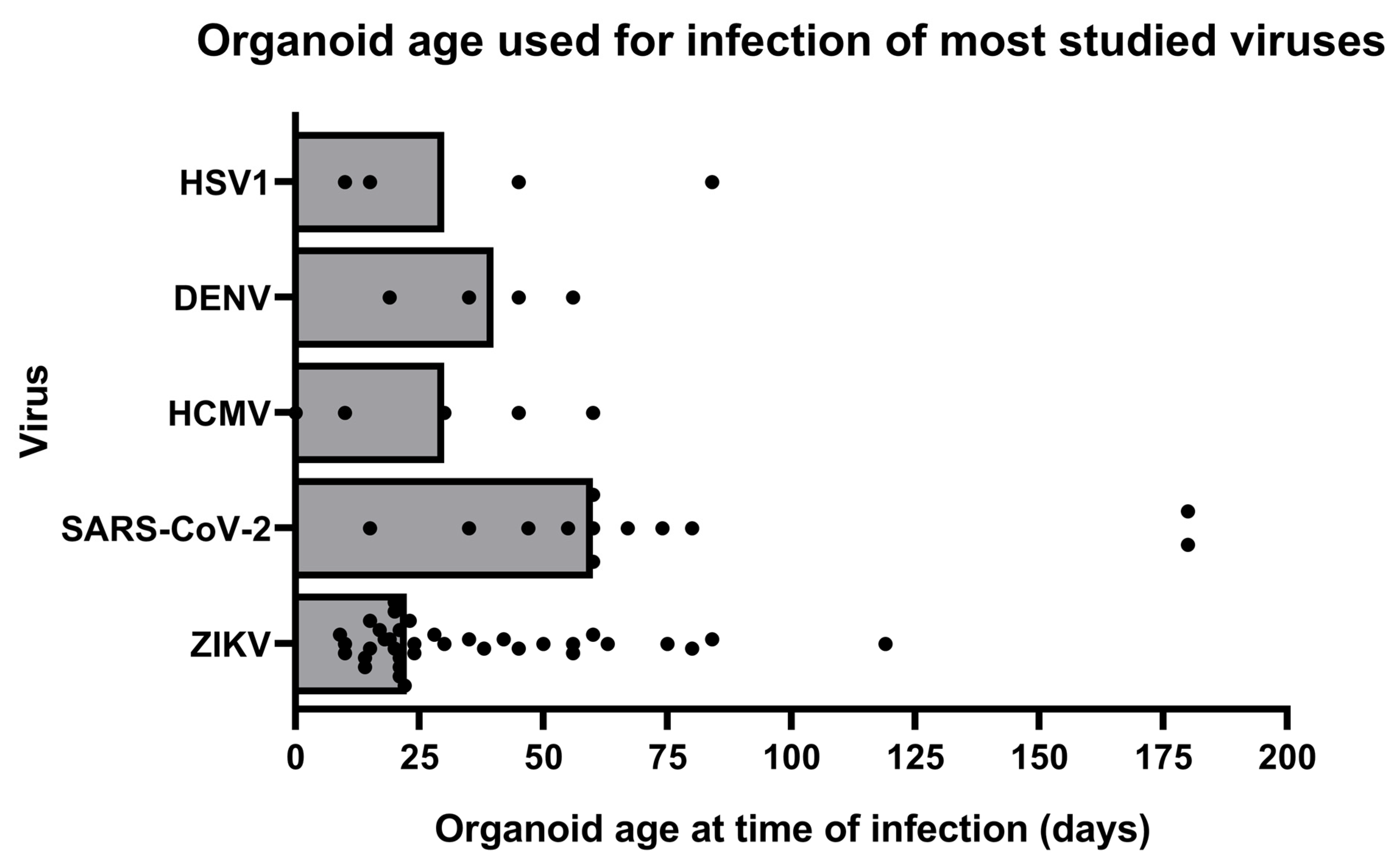
3.1. Viruses and Their Effects Studied in Brain Organoids
3.2. Outcomes of Virus Infections in Brain Organoids
3.2.1. ZIKV
3.2.2. SARS-CoV-2
3.2.3. HSV1
3.2.4. HCMV
3.2.5. DENV
3.2.6. Brain Organoid Studies on AAV, MeV, Rabies Virus, JEV and LACV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Swanson, P.A.; McGavern, D.B. Viral Diseases of the Central Nervous System. Curr. Opin. Virol. 2015, 11, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, L.L.; de Lima, R.G.; Boffi, L.C.; Bindilatti, R.N.; Garlipp, C.R.; Costa, S.C.B.; Bonon, S.H.A. Arbovirus, Herpesvirus, and Enterovirus Associated with Neurological Syndromes in Adult Patients of a University Hospital, 2017–2018. Rev. Soc. Bras. Med. Trop. 2021, 54, e0127. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, A.; Murphy, O.C. Viral Encephalitis. Neurol. Clin. 2018, 36, 705–724. [Google Scholar] [CrossRef]
- Hemachudha, P.; Petcharat, S.; Hinjoy, S.; Saraya, A.W.; Hemachudha, T. Encephalitis in Thailand: A Neglected Disease Increasingly Caused by Enterovirus. Trop. Med. Infect. Dis. 2021, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- Autore, G.; Bernardi, L.; Perrone, S.; Esposito, S. Update on Viral Infections Involving the Central Nervous System in Pediatric Patients. Children 2021, 8, 782. [Google Scholar] [CrossRef]
- Pașca, S.P. The Rise of Three-Dimensional Human Brain Cultures. Nature 2018, 553, 437–445. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, L.; Bloom, D.C.; Naciri, J.N.; Smith, A.; Edwards, T.G.; McClain, L.; Callio, J.A.; Jessup, M.; Wood, J.; Chowdari, K.; et al. Modeling Herpes Simplex Virus 1 Infections in Human Central Nervous System Neuronal Cells Using Two- and Three-Dimensional Cultures Derived from Induced Pluripotent Stem Cells. J. Virol. 2019, 93, e00111-19. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Klammer, A.M.; Naciri, J.N.; Yeung, J.; Demers, M.; Milosevic, J.; Kinchington, P.R.; Bloom, D.C.; Nimgaonkar, V.L.; D’Aiutoa, L. Patterns of Herpes Simplex Virus 1 Infection in Neural Progenitor Cells. J. Virol. 2020, 94, e00994-20. [Google Scholar] [CrossRef]
- Tekin, H.; Simmons, S.; Cummings, B.; Gao, L.; Adiconis, X.; Hession, C.C.; Ghoshal, A.; Dionne, D.; Choudhury, S.R.; Yesilyurt, V.; et al. Effects of 3D Culturing Conditions on the Transcriptomic Profile of Stem-Cell-Derived Neurons. Nat. Biomed. Eng. 2018, 2, 540–554. [Google Scholar] [CrossRef]
- Jacob, F.; Pather, S.R.; Huang, W.K.; Zhang, F.; Wong, S.Z.H.; Zhou, H.; Cubitt, B.; Fan, W.; Chen, C.Z.; Xu, M.; et al. Human Pluripotent Stem Cell-Derived Neural Cells and Brain Organoids Reveal SARS-CoV-2 Neurotropism Predominates in Choroid Plexus Epithelium. Cell Stem Cell 2020, 27, 937–950.e9. [Google Scholar] [CrossRef]
- Reddehase, M.J.; Lemmermann, N.A.W. Mouse Model of Cytomegalovirus Disease and Immunotherapy in the Immunocompromised Host: Predictions for Medical Translation That Survived the “Test of Time”. Viruses 2018, 10, 693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harschnitz, O.; Studer, L. Human Stem Cell Models to Study Host–Virus Interactions in the Central Nervous System. Nat. Rev. Immunol. 2021, 21, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Pollen, A.A.; Bhaduri, A.; Andrews, M.G.; Nowakowski, T.J.; Meyerson, O.S.; Mostajo-Radji, M.A.; Di Lullo, E.; Alvarado, B.; Bedolli, M.; Dougherty, M.L.; et al. Establishing Cerebral Organoids as Models of Human-Specific Brain Evolution. Cell 2019, 176, 743–756.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrazzo, P.; Cricca, M.; Nastasi, C. Are the Organoid Models an Invaluable Contribution to ZIKA Virus Research? Pathogens 2021, 10, 1233. [Google Scholar] [CrossRef]
- Pellegrini, L.; Albecka, A.; Mallery, D.L.; Kellner, M.J.; Paul, D.; Carter, A.P.; James, L.C.; Lancaster, M.A. SARS-CoV-2 Infects the Brain Choroid Plexus and Disrupts the Blood-CSF Barrier in Human Brain Organoids. Cell Stem Cell 2020, 27, 951–961.e5. [Google Scholar] [CrossRef]
- Watanabe, M.; Buth, J.E.; Vishlaghi, N.; de la Torre-Ubieta, L.; Taxidis, J.; Khakh, B.S.; Coppola, G.; Pearson, C.A.; Yamauchi, K.; Gong, D.; et al. Self-Organized Cerebral Organoids with Human-Specific Features Predict Effective Drugs to Combat Zika Virus Infection. Cell Rep. 2017, 21, 517–532. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, H.K.; Traverse, E.M.; Barr, K.L. Methodologies for Generating Brain Organoids to Model Viral Pathogenesis in the CNS. Pathogens 2021, 10, 1510. [Google Scholar] [CrossRef]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional Cortical Neurons and Astrocytes from Human Pluripotent Stem Cells in 3D Culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Cakir, B.; Xiang, Y.; Sullivan, G.J.; Park, I.-H. Synthetic Analyses of Single-Cell Transcriptomes from Multiple Brain Organoids and Fetal Brain. Cell Rep. 2020, 30, 1682–1689.e3. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.K.; Matsubayashi, M.; Sakaguchi, Y.M.; Hayashi, R.K.; Zheng, C.; Sugie, K.; Hasegawa, M.; Nakagawa, T.; Mori, E. Six-Month Cultured Cerebral Organoids from Human ES Cells Contain Matured Neural Cells. Neurosci. Lett. 2018, 670, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Ormel, P.R.; Vieira de Sá, R.; van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.M.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A.; et al. Microglia Innately Develop within Cerebral Organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef] [PubMed]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Song, H.; Ming, G.-L. Brain Organoids: Advances, Applications and Challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Boreland, A.J.; Li, X.; Erickson, C.; Jin, M.; Atkins, C.; Pang, Z.P.; Daniels, B.P.; Jiang, P. Developing Human Pluripotent Stem Cell-Based Cerebral Organoids with a Controllable Microglia Ratio for Modeling Brain Development and Pathology. Stem Cell Reports 2021, 16, 1923–1937. [Google Scholar] [CrossRef] [PubMed]
- Abreu, C.M.; Gama, L.; Krasemann, S.; Chesnut, M.; Odwin-Dacosta, S.; Hogberg, H.T.; Hartung, T.; Pamies, D. Microglia Increase Inflammatory Responses in IPSC-Derived Human BrainSpheres. Front. Microbiol. 2018, 9, 2766. [Google Scholar] [CrossRef]
- Caffrey, T.M.; Button, E.B.; Robert, J. Toward Three-Dimensional in Vitro Models to Study Neurovascular Unit Functions in Health and Disease. Neural Regen. Res. 2021, 16, 2132–2140. [Google Scholar] [CrossRef]
- Kook, M.G.; Lee, S.-E.; Shin, N.; Kong, D.; Kim, D.-H.; Kim, M.-S.; Kang, H.K.; Choi, S.W.; Kang, K.-S. Generation of Cortical Brain Organoid with Vascularization by Assembling with Vascular Spheroid. Int. J. Stem Cells 2022, 15, 85–94. [Google Scholar] [CrossRef]
- Wang, L.; Sievert, D.; Clark, A.E.; Lee, S.; Federman, H.; Gastfriend, B.D.; Shusta, E.V.; Palecek, S.P.; Carlin, A.F.; Gleeson, J.G. A Human Three-Dimensional Neural-Perivascular ‘Assembloid’ Promotes Astrocytic Development and Enables Modeling of SARS-CoV-2 Neuropathology. Nat. Med. 2021, 27, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Potjewyd, G.; Kellett, K.A.B.; Hooper, N.M. 3D Hydrogel Models of the Neurovascular Unit to Investigate Blood-Brain Barrier Dysfunction. Neuronal Signal. 2021, 5, NS20210027. [Google Scholar] [CrossRef]
- Fan, W.; Christian, K.M.; Song, H.; Ming, G.-L. Applications of Brain Organoids for Infectious Diseases. J. Mol. Biol. 2022, 434, 167243. [Google Scholar] [CrossRef]
- Su, X.; Yue, P.; Kong, J.; Xu, X.; Zhang, Y.; Cao, W.; Fan, Y.; Liu, M.; Chen, J.; Liu, A.; et al. Human Brain Organoids as an In Vitro Model System of Viral Infectious Diseases. Front. Immunol. 2022, 12, 792316. [Google Scholar] [CrossRef] [PubMed]
- Egilmezer, E.; Rawlinson, W.D. Review of Studies of Severe Acute Respiratory Syndrome Related Coronavirus-2 Pathogenesis in Human Organoid Models. Rev. Med. Virol. 2021, 31, e2227. [Google Scholar] [CrossRef] [PubMed]
- Ramani, A.; Pranty, A.-I.; Gopalakrishnan, J. Neurotropic Effects of SARS-CoV-2 Modeled by the Human Brain Organoids. Stem Cell Rep. 2021, 16, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.I.; Abdullah, M.; Ali, S.; Naqvi, I.H.; Ahmed, A.; Parveen, S. Zika Virus-Induced Microcephaly and Its Possible Molecular Mechanism. Intervirology 2016, 59, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Krenn, V.; Bosone, C.; Burkard, T.R.; Garcez, P.P.; Mirazimi, A.; Knoblich, A. Organoid Modeling of Zika and Herpes Simplex Virus 1 Infections Reveals Virus-Specific Responses Leading to Microcephaly. Cell Stem Cell 2021, 28, 1362–1379.e7. [Google Scholar] [CrossRef]
- Albanese, A.; Swaney, J.M.; Yun, D.H.; Evans, N.B.; Antonucci, J.M.; Velasco, S.; Sohn, C.H.; Arlotta, P.; Gehrke, L.; Chung, K. Multiscale 3D Phenotyping of Human Cerebral Organoids. Sci. Rep. 2020, 10, 21487. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Qin, Y.; Patil, V.S.; Eroshkin, A.M.; Rana, T.M. Zika Virus Depletes Neural Progenitors in Human Cerebral Organoids through Activation of the Innate Immune Receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain-Region-Specific Organoids Using Mini-Bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [Green Version]
- Salick, M.R.; Wells, M.F.; Eggan, K.; Kaykas, A. Modelling Zika Virus Infection of the Developing Human Brain In Vitro Using Stem Cell Derived Cerebral Organoids. J. Vis. Exp. 2017, e56404. [Google Scholar] [CrossRef] [Green Version]
- Setoh, Y.X.; Amarilla, A.A.; Peng, N.Y.G.; Griffiths, R.E.; Carrera, J.; Freney, M.E.; Nakayama, E.; Ogawa, S.; Watterson, D.; Modhiran, N.; et al. Determinants of Zika Virus Host Tropism Uncovered by Deep Mutational Scanning. Nat. Microbiol. 2019, 4, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.F.; Salick, M.R.; Wiskow, O.; Ho, D.J.; Worringer, K.A.; Ihry, R.J.; Kommineni, S.; Bilican, B.; Klim, J.R.; Hill, E.J.; et al. Genetic Ablation of AXL Does Not Protect Human Neural Progenitor Cells and Cerebral Organoids from Zika Virus Infection. Cell Stem Cell 2016, 19, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.P.; Qiu, Y.; Zhang, B.; Chen, G.; Chen, Q.; Wang, M.; Mo, F.; Xu, J.; Wu, J.; Zhang, R.R.; et al. Zika Virus Infection Induces RNAi-Mediated Antiviral Immunity in Human Neural Progenitors and Brain Organoids. Cell Res. 2019, 29, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcez, P.P.; Loiola, E.C.; Da Costa, R.M.; Higa, L.M.; Trindade, P.; Delvecchio, R.; Nascimento, J.M.; Brindeiro, R.; Tanuri, A.; Rehen, S.K. Zika Virus: Zika Virus Impairs Growth in Human Neurospheres and Brain Organoids. Science 2016, 352, 816–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Deng, Y.Q.; Wang, S.; Ma, F.; Aliyari, R.; Huang, X.Y.; Zhang, N.N.; Watanabe, M.; Dong, H.L.; Liu, P.; et al. 25-Hydroxycholesterol Protects Host against Zika Virus Infection and Its Associated Microcephaly in a Mouse Model. Immunity 2017, 46, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Muffat, J.; Omer, A.; Bosch, I.; Lancaster, M.A.; Sur, M.; Gehrke, L.; Knoblich, J.A.; Jaenisch, R. Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell 2017, 20, 385–396.e3. [Google Scholar] [CrossRef] [Green Version]
- Wiwanitkit, V. Neuroimaging Findings of Zika Virus Infection. Jpn. J. Radiol. 2017, 35, 40. [Google Scholar] [CrossRef]
- Pettke, A.; Tampere, M.; Pronk, R.; Wallner, O.; Falk, A.; Berglund, U.W.; Helleday, T.; Mirazimi, A.; Puumalainen, M.R. Broadly Active Antiviral Compounds Disturb Zika Virus Progeny Release Rescuing Virus-Induced Toxicity in Brain Organoids. Viruses 2021, 13, 37. [Google Scholar] [CrossRef]
- Cavalcante, B.R.R.; Aragão-França, L.S.; Sampaio, G.L.A.; Nonaka, C.K.V.; Oliveira, M.S.; Campos, G.S.; Sardi, S.I.; Dias, B.R.S.; Menezes, J.P.B.; Rocha, V.P.C.; et al. Betulinic Acid Exerts Cytoprotective Activity on Zika Virus-Infected Neural Progenitor Cells. Front. Cell. Infect. Microbiol. 2020, 10, 558324. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Z.; Zhang, X.; Li, S.; Hui, Y.; Feng, H.; Du, Y.; Jin, G.; Zhou, X.; Zhang, X. Protection of ZIKV Infection-Induced Neuropathy by Abrogation of Acute Antiviral Response in Human Neural Progenitors. Cell Death Differ. 2019, 26, 2607–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Tan, L.; Cederquist, G.Y.; Fan, Y.; Hartley, B.J.; Mukherjee, S.; Tomishima, M.; Brennand, K.J.; Zhang, Q.; Schwartz, R.E.; et al. High-Content Screening in HPSC-Neural Progenitors Identifies Drug Candidates That Inhibit Zika Virus Infection in Fetal-like Organoids and Adult Brain. Cell Stem Cell 2017, 21, 274–283.e5. [Google Scholar] [CrossRef] [PubMed]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian Zika Virus Strain Causes Birth Defects in Experimental Models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, E.; Ramani, A.; Karow, U.; Gottardo, M.; Natarajan, K.; Gooi, L.M.; Goranci-Buzhala, G.; Krut, O.; Peters, F.; Nikolic, M.; et al. Recent Zika Virus Isolates Induce Premature Differentiation of Neural Progenitors in Human Brain Organoids. Cell Stem Cell 2017, 20, 397–406.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, S.; Schotsaert, M.; Karnik, R.; Balasubramaniam, V.; Dejosez, M.; Meissner, A.; García-Sastre, A.; Zwaka, T.P. Zika Virus Alters DNA Methylation of Neural Genes in an Organoid Model of the Developing Human Brain. mSystems 2018, 31, e00219-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, S.; Schotsaert, M.; Manganaro, L.; Dejosez, M.; Simon, V.; García-Sastre, A.; Zwaka, T.P. FACS-Mediated Isolation of Neuronal Cell Populations From Virus-Infected Human Embryonic Stem Cell–Derived Cerebral Organoid Cultures. Curr. Protoc. Stem Cell Biol. 2019, 48, e65. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, D.; Li, G. The Role of Microglia in Viral Encephalitis: A Review. J. Neuroinflammation 2019, 16, 76. [Google Scholar] [CrossRef]
- Li, Z.; Xu, J.; Lang, Y.; Fan, X.; Kuo, L.; D’Brant, L.; Hu, S.; Samrat, S.K.; Trudeau, N.; Tharappel, A.M.; et al. JMX0207, a Niclosamide Derivative with Improved Pharmacokinetics, Suppresses Zika Virus Infection Both in Vitro and in Vivo. ACS Infect. Dis. 2020, 6, 2616–2628. [Google Scholar] [CrossRef]
- Li, Z.; Lang, Y.; Sakamuru, S.; Samrat, S.; Trudeau, N.; Kuo, L.; Rugenstein, N.; Tharappel, A.; D’Brant, L.; Koetzner, C.A.; et al. Methylene Blue Is a Potent and Broad-Spectrum Inhibitor against Zika Virus in Vitro and in Vivo. Emerg. Microbes Infect. 2020, 9, 2404–2416. [Google Scholar] [CrossRef]
- Sacramento, C.Q.; De Melo, G.R.; De Freitas, C.S.; Rocha, N.; Hoelz, L.V.B.; Miranda, M.; Fintelman-Rodrigues, N.; Marttorelli, A.; Ferreira, A.C.; Barbosa-Lima, G.; et al. The Clinically Approved Antiviral Drug Sofosbuvir Inhibits Zika Virus Replication. Sci. Rep. 2017, 7, 40920. [Google Scholar] [CrossRef]
- Xu, M.; Lee, E.M.; Wen, Z.; Cheng, Y.; Huang, W.K.; Qian, X.; Tcw, J.; Kouznetsova, J.; Ogden, S.C.; Hammack, C.; et al. Identification of Small-Molecule Inhibitors of Zika Virus Infection and Induced Neural Cell Death via a Drug Repurposing Screen. Nat. Med. 2016, 22, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Di Lullo, E.; Kriegstein, A.R. The Use of Brain Organoids to Investigate Neural Development and Disease. Nat. Rev. Neurosci. 2017, 18, 573–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meertens, L.; Labeau, A.; Dejarnac, O.; Cipriani, S.; Sinigaglia, L.; Bonnet-Madin, L.; Le Charpentier, T.; Hafirassou, M.L.; Zamborlini, A.; Cao-Lormeau, V.-M.; et al. Axl Mediates ZIKA Virus Entry in Human Glial Cells and Modulates Innate Immune Responses. Cell Rep. 2017, 18, 324–333. [Google Scholar] [CrossRef]
- Li, C.; Xu, D.; Ye, Q.; Hong, S.; Jiang, Y.; Liu, X.; Zhang, N.; Shi, L.; Qin, C. Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almutairi, M.M.; Sivandzade, F.; Albekairi, T.H.; Alqahtani, F.; Cucullo, L. Neuroinflammation and Its Impact on the Pathogenesis of COVID-19. Front. Med. 2021, 8, 745789. [Google Scholar] [CrossRef] [PubMed]
- Del Brutto, O.H.; Wu, S.; Mera, R.M.; Costa, A.F.; Recalde, B.Y.; Issa, N.P. Cognitive Decline among Individuals with History of Mild Symptomatic SARS-CoV-2 Infection: A Longitudinal Prospective Study Nested to a Population Cohort. Eur. J. Neurol. 2021, 28, 3245–3253. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.-H.; Sun, A.; Je, H.S.; Tan, E.-K. Unravelling Pathophysiology of Neurological and Psychiatric Complications of COVID-19 Using Brain Organoids. Neuroscientist 2021. [Google Scholar] [CrossRef]
- Chen, R.; Wang, K.; Yu, J.; Howard, D.; French, L.; Chen, Z.; Wen, C.; Xu, Z. The Spatial and Cell-Type Distribution of SARS-CoV-2 Receptor ACE2 in the Human and Mouse Brains. Front. Neurol. 2021, 11, 1860. [Google Scholar] [CrossRef]
- Bullen, C.K.; Hogberg, H.T.; Bahadirli-Talbott, A.; Bishai, W.R.; Hartung, T.; Keuthan, C.; Looney, M.M.; Pekosz, A.; Romero, J.C.; Sillé, F.C.M.; et al. Infectability of Human BrainSphere Neurons Suggests Neurotropism of SARS-CoV-2. ALTEX 2020, 37, 665–671. [Google Scholar] [CrossRef]
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S.; Lu, P.; Weizman, O.-E.; Liu, F.; Dai, Y.; et al. Neuroinvasion of SARS-CoV-2 in Human and Mouse Brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.K.; Wang, S.; Smith, D.; Carlin, A.F.; Rana, T.M. Revealing Tissue-Specific SARS-CoV-2 Infection and Host Responses Using Human Stem Cell-Derived Lung and Cerebral Organoids. Stem Cell Reports 2021, 16, 437–445. [Google Scholar] [CrossRef]
- Yi, S.A.; Nam, K.H.; Yun, J.; Gim, D.; Joe, D.; Kim, Y.H.; Kim, H.J.; Han, J.W.; Lee, J. Infection of Brain Organoids and 2D Cortical Neurons with SARS-CoV-2 Pseudovirus. Viruses 2020, 12, 1004. [Google Scholar] [CrossRef] [PubMed]
- McMahon, C.L.; Staples, H.; Gazi, M.; Carrion, R.; Hsieh, J. SARS-CoV-2 Targets Glial Cells in Human Cortical Organoids. Stem Cell Rep. 2021, 16, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Ramani, A.; Müller, L.; Ostermann, P.N.; Gabriel, E.; Abida-Islam, P.; Müller-Schiffmann, A.; Mariappan, A.; Goureau, O.; Gruell, H.; Walker, A.; et al. SARS -CoV-2 Targets Neurons of 3D Human Brain Organoids. EMBO J. 2020, 39, e106230. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, M.; Garcia, G.; Tian, E.; Cui, Q.; Chen, X.; Sun, G.; Wang, J.; Arumugaswami, V.; Shi, Y. ApoE-Isoform-Dependent SARS-CoV-2 Neurotropism and Cellular Response. Cell Stem Cell 2021, 28, 331–342.e5. [Google Scholar] [CrossRef]
- Zhang, B.Z.; Chu, H.; Han, S.; Shuai, H.; Deng, J.; Hu, Y.F.; Gong, H.R.; Lee, A.C.Y.; Zou, Z.; Yau, T.; et al. SARS-CoV-2 Infects Human Neural Progenitor Cells and Brain Organoids. Cell Res. 2020, 30, 928–931. [Google Scholar] [CrossRef]
- Pinninti, S.G.; Kimberlin, D.W. Neonatal Herpes Simplex Virus Infections. Semin. Perinatol. 2018, 42, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Guo, M.; Shang, J.; Zhao, W.; Wang, Z.; Liu, N.; Li, B.; Zhou, Y.; Wu, Y.; Chen, P. Herpes Simplex Virus Type 1 Infection Leads to Neurodevelopmental Disorder-Associated Neuropathological Changes. PLoS Pathog. 2020, 16, e1008899. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Zheng, W.; Muralidaran, V.; Ikonomovic, M.D.; Bloom, D.C.; Nimgaonkar, V.L.; D’Aiuto, L. Modeling Aβ42 Accumulation in Response to Herpes Simplex Virus 1 Infection: 2D or 3D? J. Virol. 2020, 95, e02219-20. [Google Scholar] [CrossRef]
- Boppana, S.B.; Ross, S.A.; Fowler, K.B. Congenital Cytomegalovirus Infection: Clinical Outcome. Clin. Infect. Dis. 2013, 57, S178–S181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sison, S.L.; O’Brien, B.S.; Johnson, A.J.; Seminary, E.R.; Terhune, S.S.; Ebert, A.D. Human Cytomegalovirus Disruption of Calcium Signaling in Neural Progenitor Cells and Organoids. J. Virol. 2019, 93, e00954-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Chiuppesi, F.; Chen, X.; Diamond, D.J.; Wussow, F.; Shi, Y.; Sun, G.; Chiuppesi, F.; Chen, X.; Wang, C.; et al. Modeling Human Cytomegalovirus-Induced Microcephaly in Human IPSC-Derived Brain Organoids. Cell Rep. Med. 2020, 1, 100002. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.M.; Rana, P.S.J.B.; Jaeger, H.K.; O’Dowd, J.M.; Balemba, O.B.; Fortunato, E.A. Human Cytomegalovirus Compromises Development of Cerebral Organoids. J. Virol. 2019, 93, e00957-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.-H.; Ning, Z.-J.; Liu, Y.-M.; Li, X.-H. Neurological Manifestations of Dengue Infection. Front. Cell. Infect. Microbiol. 2017, 7, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paixão, E.S.; Teixeira, M.G.; Costa, M. da C.N.; Barreto, M.L.; Rodrigues, L.C. Symptomatic Dengue during Pregnancy and Congenital Neurologic Malformations. Emerg. Infect. Dis. 2018, 24, 1748–1750. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Song, G.; Qian, X.; Pan, J.; Xu, D.; Rho, H.S.; Kim, N.S.; Habela, C.; Zheng, L.; Jacob, F.; et al. Zika-Virus-Encoded NS2A Disrupts Mammalian Cortical Neurogenesis by Degrading Adherens Junction Proteins. Cell Stem Cell 2017, 21, 349–358.e6. [Google Scholar] [CrossRef] [Green Version]
- Kunze, C.; Börner, K.; Kienle, E.; Orschmann, T.; Rusha, E.; Schneider, M.; Radivojkov-Blagojevic, M.; Drukker, M.; Desbordes, S.; Grimm, D.; et al. Synthetic AAV/CRISPR Vectors for Blocking HIV-1 Expression in Persistently Infected Astrocytes. Glia 2018, 66, 413–427. [Google Scholar] [CrossRef]
- Depla, J.A.; Sogorb-Gonzalez, M.; Mulder, L.A.; Heine, V.M.; Konstantinova, P.; van Deventer, S.J.; Wolthers, K.C.; Pajkrt, D.; Sridhar, A.; Evers, M.M. Cerebral Organoids: A Human Model for AAV Capsid Selection and Therapeutic Transgene Efficacy in the Brain. Mol. Ther.—Methods Clin. Dev. 2020, 18, 167–175. [Google Scholar] [CrossRef]
- Latour, Y.L.; Yoon, R.; Thomas, S.E.; Grant, C.; Li, C.; Sena-Esteves, M.; Allende, M.L.; Proia, R.L.; Tifft, C.J. Human GLB1 Knockout Cerebral Organoids: A Model System for Testing AAV9-Mediated GLB1 Gene Therapy for Reducing GM1 Ganglioside Storage in GM1 Gangliosidosis. Mol. Genet. Metab. Rep. 2019, 21, 100513. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Hudry, E.; Maguire, C.A.; Sena-esteves, M.; Breakefield, X.O.; Grandi, P.; Hospital, M.G.; Hospital, M.G. Viral Vectors for Therapy of Neurologic Diseases. Neuropharmacology 2018, 120, 63–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Shirogane, Y.; Sato, Y.; Hashiguchi, T.; Yanagi, Y. New Insights into Measles Virus Brain Infections. Trends Microbiol. 2019, 27, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Bovier, F.T.; Ferren, M.; Lieberman, N.A.P.; Predella, C.; Lalande, A.; Peddu, V.; Lin, M.J.; Addetia, A.; Patel, A.; et al. Molecular Features of the Measles Virus Viral Fusion Complex That Favor Infection and Spread in the Brain. MBio 2021, 12, e00799-21. [Google Scholar] [CrossRef] [PubMed]
- Schubert, R.; Trenholm, S.; Balint, K.; Kosche, G.; Cowan, C.S.; Mohr, M.A.; Munz, M.; Martinez-Martin, D.; Fläschner, G.; Newton, R.; et al. Virus Stamping for Targeted Single-Cell Infection in Vitro and in Vivo. Nat. Biotechnol. 2018, 36, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Schultz, E.M.; Jones, T.J.; Xu, S.; Dean, D.D.; Zechmann, B.; Barr, K.L. Cerebral Organoids Derived from a Parkinson’s Patient Exhibit Unique Pathogenesis from Chikungunya Virus Infection When Compared to a Non-Parkinson’s Patient. Pathogens 2021, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Barreras, P.; Pamies, D.; Monaco, M.C.; Muñoz, L.S.; Zhong, X.; Major, E.O.; Hogberg, H.T.; Hartung, T.; Pardo, C.A. A Human-Derived 3D Brain Organoid Model to Study JC Virus Infection. J. Neurovirol. 2022, 1–10. [Google Scholar] [CrossRef]
- Takahashi, K.; Sekizuka, T.; Fukumoto, H.; Nakamichi, K.; Suzuki, T.; Sato, Y.; Hasegawa, H.; Kuroda, M.; Katano, H. Deep-Sequence Identification and Role in Virus Replication of a JC Virus Quasispecies in Patients with Progressive Multifocal Leukoencephalopathy. J. Virol. 2017, 91, e01335-16. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.H.; Kardas, P.; Kranz, D.; Leboeuf, C. The Human JC Polyomavirus (JCPyV): Virological Background and Clinical Implications. APMIS 2013, 121, 685–727. [Google Scholar] [CrossRef]
- Zhang, B.; He, Y.; Xu, Y.; Mo, F.; Mi, T.; Shen, Q.S.; Li, C.; Li, Y.; Liu, J.; Wu, Y.; et al. Differential Antiviral Immunity to Japanese Encephalitis Virus in Developing Cortical Organoids. Cell Death Dis. 2018, 9, 719. [Google Scholar] [CrossRef]
- Vahey, G.M.; Lindsey, N.P.; Staples, J.E.; Hills, S.L. La Crosse Virus Disease in the United States, 2003-2019. Am. J. Trop. Med. Hyg. 2021, 105, 807–812. [Google Scholar] [CrossRef]
- Winkler, C.W.; Woods, T.A.; Groveman, B.R.; Carmody, A.B.; Speranza, E.E.; Martens, C.A.; Best, S.M.; Haigh, C.L.; Peterson, K.E. Neuronal Maturation Reduces the Type I IFN Response to Orthobunyavirus Infection and Leads to Increased Apoptosis of Human Neurons. J. Neuroinflammation 2019, 16, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojha, D.; Winkler, C.W.; Leung, J.M.; Woods, T.A.; Chen, C.Z.; Nair, V.; Taylor, K.; Yeh, C.D.; Tawa, G.J.; Larson, C.L.; et al. Rottlerin Inhibits La Crosse Virus-Induced Encephalitis in Mice and Blocks Release of Replicating Virus from the Golgi Body in Neurons. Nat. Microbiol. 2021, 6, 1398–1409. [Google Scholar] [CrossRef] [PubMed]
- Kathuria, A.; Lopez-Lengowski, K.; Watmuff, B.; Karmacharya, R. Comparative Transcriptomic Analysis of Cerebral Organoids and Cortical Neuron Cultures Derived from Human Induced Pluripotent Stem Cells. Stem Cells Dev. 2020, 29, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Shan, C.; Dunn, T.J.; Xie, X.; Xia, H.; Gao, J.; Allende Labastida, J.; Zou, J.; Villarreal, P.P.; Schlagal, C.R.; et al. Role of Microglia in the Dissemination of Zika Virus from Mother to Fetal Brain. PLoS Negl. Trop. Dis. 2020, 14, e0008413. [Google Scholar] [CrossRef]
- Vasek, M.J.; Garber, C.; Dorsey, D.; Durrant, D.M.; Bollman, B.; Soung, A.; Yu, J.; Perez-Torres, C.; Frouin, A.; Wilton, D.K.; et al. A Complement-Microglial Axis Drives Synapse Loss during Virus-Induced Memory Impairment. Nature 2016, 534, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, A.; Simmini, S.; Ribeiro, C.M.S.; Tapparel, C.; Evers, M.M.; Pajkrt, D.; Wolthers, K. A Perspective on Organoids for Virology Research. Viruses 2020, 12, 1341. [Google Scholar] [CrossRef]
- Dos Reis, R.S.; Sant, S.; Keeney, H.; Wagner, M.C.E.; Ayyavoo, V. Modeling HIV-1 Neuropathogenesis Using Three-Dimensional Human Brain Organoids (HBORGs) with HIV-1 Infected Microglia. Sci. Rep. 2020, 10, 15209. [Google Scholar] [CrossRef]
- Tanaka, Y.; Park, I.-H. Regional Specification and Complementation with Non-Neuroectodermal Cells in Human Brain Organoids. J. Mol. Med. 2021, 99, 489–500. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Virus | ZIKV | SARS-CoV-2 | DENV | HSV1 | HCMV | All Viruses |
|---|---|---|---|---|---|---|
| Viral Infection | 27/30 (90%) | 11/11 (100%) | 2/4 (50%) | 4/4 (100%) | 4/4 (100%) | 57/63 (90%) |
| Receptor expression | 2/30 (7%) | 8/11 (73%) | 0/4 (0%) | 0/4 (0%) | 1/4 (25%) | 11/63 (17%) |
| Organoid organization and size | 21/30 (70%) | 3/11 (27%) | 3/4 (75%) | 3/4 (75%) | 1/4 (25%) | 37/63 (59%) |
| Viral tropism | 14/30 (47%) | 10/11 (91%) | 1/4 (25%) | 3/4 (75%) | 2/4 (50%) | 34/63 (54%) |
| Cytopathic effect | 17/30 (57%) | 5/11 (45%) | 2/4 (50%) | 1/4 (25%) | 1/4 (25%) | 28/63 (44%) |
| Effect of infection on proliferation | 7/30 (23%) | 1/11 (9%) | 2/4 (50%) | 0/4 (0%) | 1/4 (25%) | 12/63 (19%) |
| Antiviral innate Immune response | 7/30 (23%) | 3/11 (27%) | 1/4 (25%) | 3/4 (75%) | 0/0 (0%) | 17/63 (27%) |
| Gene expression profile | 4/30 (13%) | 4/11 (36%) | 0/4 (0%) | 1/4 (25%) | 1/4 (25%) | 14/63 (22%) |
| Antiviral drug testing | 12/30 (40%) | 0/11 (0%) | 0/4 (0%) | 0/4 (25%) | 2/4 (50%) | 18/63 (29%) |
| Virus | Main Finding in Brain Organoids |
|---|---|
| ZIKV |
|
| SARS-CoV-2 |
|
| HSV1 |
|
| HCMV |
|
| DENV |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Depla, J.A.; Mulder, L.A.; de Sá, R.V.; Wartel, M.; Sridhar, A.; Evers, M.M.; Wolthers, K.C.; Pajkrt, D. Human Brain Organoids as Models for Central Nervous System Viral Infection. Viruses 2022, 14, 634. https://doi.org/10.3390/v14030634
Depla JA, Mulder LA, de Sá RV, Wartel M, Sridhar A, Evers MM, Wolthers KC, Pajkrt D. Human Brain Organoids as Models for Central Nervous System Viral Infection. Viruses. 2022; 14(3):634. https://doi.org/10.3390/v14030634
Chicago/Turabian StyleDepla, Josse A., Lance A. Mulder, Renata Vieira de Sá, Morgane Wartel, Adithya Sridhar, Melvin M. Evers, Katja C. Wolthers, and Dasja Pajkrt. 2022. "Human Brain Organoids as Models for Central Nervous System Viral Infection" Viruses 14, no. 3: 634. https://doi.org/10.3390/v14030634
APA StyleDepla, J. A., Mulder, L. A., de Sá, R. V., Wartel, M., Sridhar, A., Evers, M. M., Wolthers, K. C., & Pajkrt, D. (2022). Human Brain Organoids as Models for Central Nervous System Viral Infection. Viruses, 14(3), 634. https://doi.org/10.3390/v14030634