
No Evidence for Orthohepevirus C in Archived Human Samples in Germany, 2000–2020


Abstract
:1. Introduction
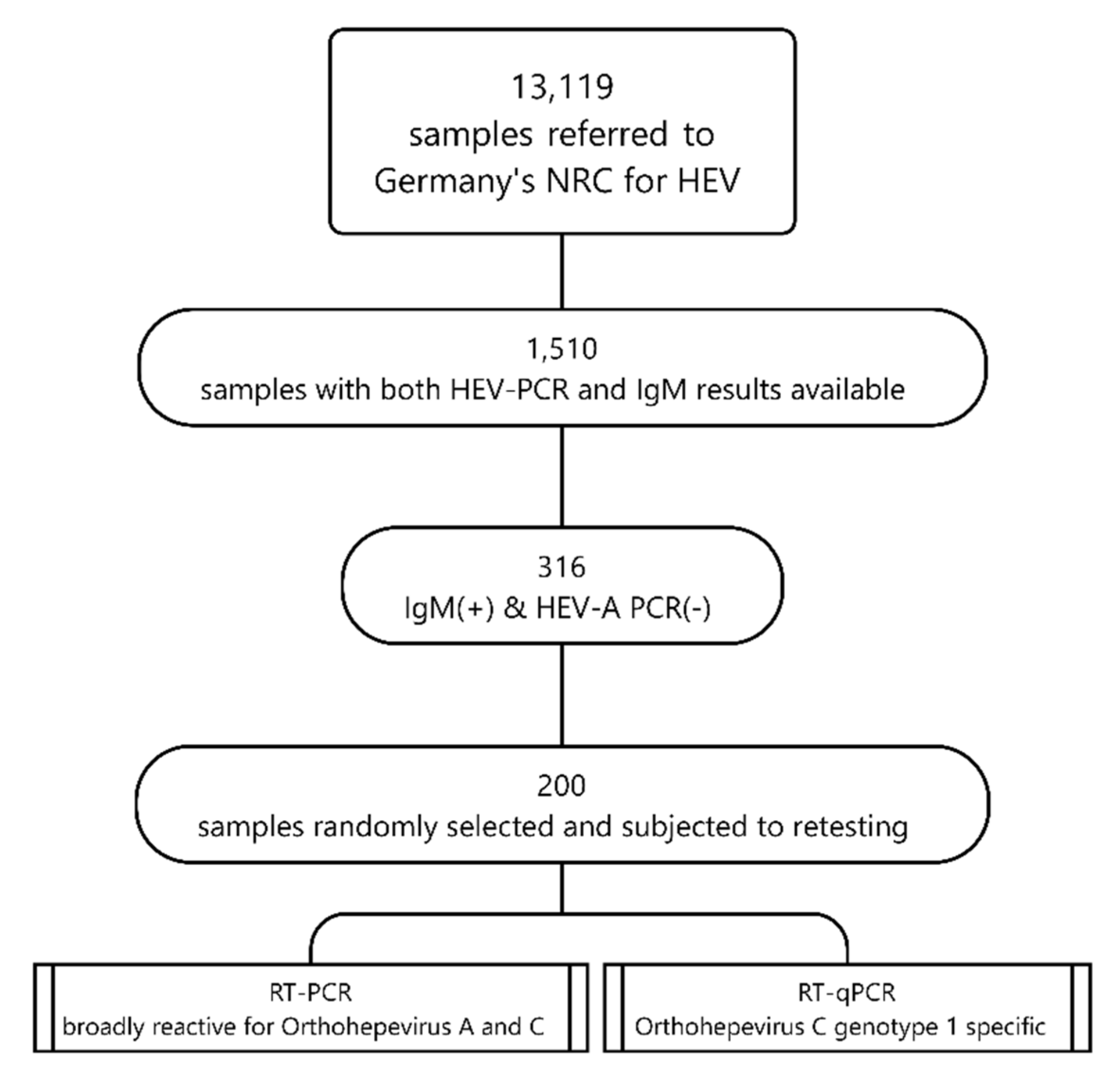
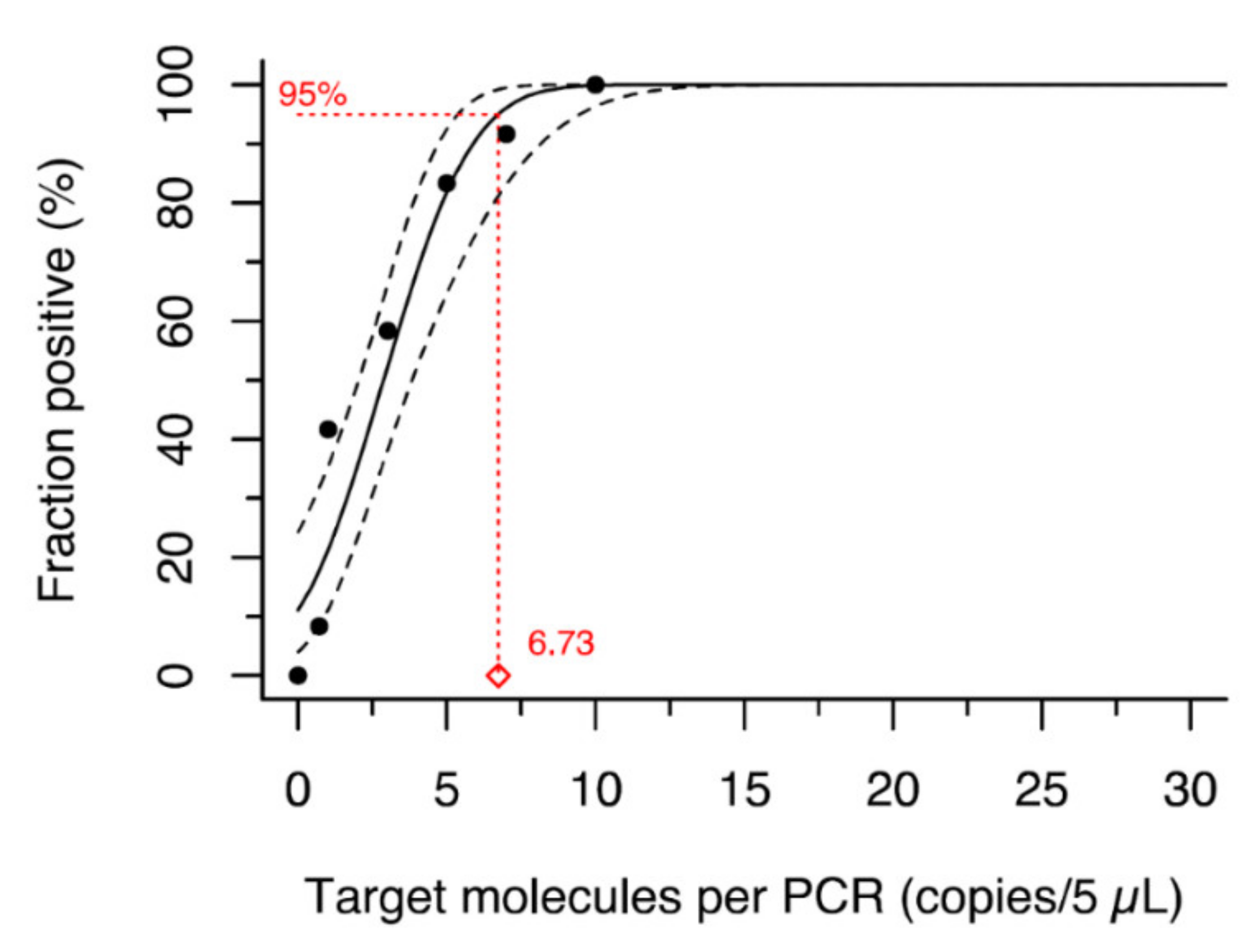
2. Materials and Methods
- an Orthohepevirus C genotype 1 specific RT-qPCR that was specifically established for this study.
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kamar, N.; Dalton, H.R.; Abravanel, F.; Izopet, J. Hepatitis E virus infection. Clin. Microbiol. Rev. 2014, 27, 116–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.-H.; Tan, B.-H.; Teo, E.C.-Y.; Lim, S.-G.; Dan, Y.-Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.; Tan, C.-K.; et al. Chronic Infection with Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Simmonds, P.; Members of The International Committee on the Taxonomy of Viruses Study G; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.-J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Johne, R.; Plenge-Bönig, A.; Hess, M.; Ulrich, R.G.; Reetz, J.; Schielke, A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J. Gen. Virol. 2010, 91, 750–758. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pankovics, P. Review of Hepatitis E Virus in Rats: Evident Risk of Species Orthohepevirus C to Human Zoonotic Infection and Disease. Viruses 2020, 12, 1148. [Google Scholar] [CrossRef]
- Wang, B.; Harms, D.; Yang, X.-L.; Bock, C.-T. Orthohepevirus C: An Expanding Species of Emerging Hepatitis E Virus Variants. Pathogens 2020, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Purcell, R.H.; Engle, R.E.; Rood, M.P.; Kabrane-Lazizi, Y.; Nguyen, H.T.; Govindarajan, S.; Claire, M.S.; Emerson, S.U. Hepatitis E Virus in Rats, Los Angeles, California, USA. Emerg. Infect. Dis. 2011, 17, 2216–2222. [Google Scholar] [CrossRef]
- Cossaboom, C.M.; Córdoba, L.; Sanford, B.J.; Piñeyro, P.; Kenney, S.P.; Dryman, B.A.; Wang, Y.; Meng, X.-J. Cross-species infection of pigs with a novel rabbit, but not rat, strain of hepatitis E virus isolated in the United States. J. Gen. Virol. 2012, 93, 1687–1695. [Google Scholar] [CrossRef]
- Andonov, A.; Robbins, M.; Borlang, J.; Cao, J.; Hatchette, T.; Stueck, A.; Deschaumbault, Y.; Murnaghan, K.; Varga, J.; Johnston, B. Rat Hepatitis E Virus Linked to Severe Acute Hepatitis in an Immunocompetent Patient. J. Infect. Dis. 2019, 220, 951–955. [Google Scholar] [CrossRef]
- Sridhar, S.; Yip, C.C.; Wu, S.; Cai, J.; Zhang, A.J.-X.; Leung, K.-H.; Chung, T.W.; Chan, J.F.; Chan, W.-M.; Teng, J.L.; et al. Rat Hepatitis E Virus as Cause of Persistent Hepatitis after Liver Transplant. Emerg. Infect. Dis. 2018, 24, 2241–2250. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Yip, C.C.; Wu, S.; Chew, N.F.; Leung, K.; Chan, J.F.; Zhao, P.S.; Chan, W.; Poon, R.W.; Tsoi, H.; et al. Transmission of Rat Hepatitis E Virus Infection to Humans in Hong Kong: A Clinical and Epidemiological Analysis. Hepatology 2021, 73, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Yip, C.C.-Y.; Lo, K.H.-Y.; Wu, S.; Situ, J.; Chew, N.F.-S.; Leung, K.-H.; Chan, H.S.-Y.; Wong, S.C.-Y.; Leung, A.W.-S.; et al. Hepatitis E Virus Species C Infection in Humans, Hong Kong. Clin. Infect. Dis. 2021, ciab919. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Perez, A.B.; Pineda, J.A.; Reina, G.; Fuentes-Lopez, A.; Freyre-Carrillo, C.; Ramirez-Arellano, E.; Alados, J.C.; Rivero, A. Orthohepevirus C infection as an emerging cause of acute hepatitis in Spain: First report in Europe. J. Hepatol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Situ, J.; Cai, J.-P.; Yip, C.C.-Y.; Wu, S.; Zhang, A.J.-X.; Wen, L.; Chew, N.F.-S.; Chan, W.-M.; Poon, R.W.-S.; et al. Multimodal investigation of rat hepatitis E virus antigenicity: Implications for infection, diagnostics, and vaccine efficacy. J. Hepatol. 2021, 74, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Pankovics, P.; Némethy, O.; Boros, Á.; Pár, G.; Szakály, P.; Reuter, G. Four-year long (2014–2017) clinical and laboratory surveillance of hepatitis E virus infections using combined antibody, molecular, antigen and avidity detection methods: Increasing incidence and chronic HEV case in Hungary. J. Clin. Virol. 2020, 124, 104284. [Google Scholar] [CrossRef] [PubMed]
- Parraud, D.; Lhomme, S.; Péron, J.M.; Da Silva, I.; Tavitian, S.; Kamar, N.; Izopet, J.; Abravanel, F. Rat Hepatitis E Virus: Presence in Humans in South-Western France? Front. Med. 2021, 8, 1485. [Google Scholar] [CrossRef]
- Garson, J.A.; Ferns, R.B.; Grant, P.R.; Ijaz, S.; Nastouli, E.; Szypulska, R.; Tedder, R. Minor groove binder modification of widely used TaqMan probe for hepatitis E virus reduces risk of false negative real-time PCR results. J. Virol. Methods 2012, 186, 157–160. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Wenzel, J.J.; Preiß, J.; Schemmerer, M.; Huber, B.; Plentz, A.; Jilg, W. Detection of hepatitis E virus (HEV) from porcine livers in Southeastern Germany and high sequence homology to human HEV isolates. J. Clin. Virol. 2011, 52, 50–54. [Google Scholar] [CrossRef]
- Li, T.-C.; Yang, T.; Yoshizaki, S.; Ami, Y.; Suzaki, Y.; Ishii, K.; Haga, K.; Nakamura, T.; Ochiai, S.; Takaji, W.; et al. Construction and characterization of an infectious cDNA clone of rat hepatitis E virus. J. Gen. Virol. 2015, 96, 1320–1327. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.-J.; Norder, H.; Okamoto, H.; Van Der Poel, W.H.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R. Finding Protein and Nucleotide Similarities with FASTA. Curr. Protoc. Bioinform. 2016, 53, 3.9.1–3.9.25. [Google Scholar] [CrossRef] [PubMed]
- Ryll, R.; Bernstein, S.; Heuser, E.; Schlegel, M.; Dremsek, P.; Zumpe, M.; Wolf, S.; Pépin, M.; Bajomi, D.; Müller, G.; et al. Detection of rat hepatitis E virus in wild Norway rats (Rattus norvegicus) and Black rats (Rattus rattus) from 11 European countries. Vet. Microbiol. 2017, 208, 58–68. [Google Scholar] [CrossRef]
- Dremsek, P.; Wenzel, J.; Johne, R.; Ziller, M.; Hofmann, J.; Groschup, M.H.; Werdermann, S.; Mohn, U.; Dorn, S.; Motz, M.; et al. Seroprevalence study in forestry workers from eastern Germany using novel genotype 3- and rat hepatitis E virus-specific immunoglobulin G ELISAs. Med. Microbiol. Immunol. 2011, 201, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Askar, M.; Stark, K. Case-control study on risk factors for acute hepatitis E in Germany, 2012 to 2014. Eurosurveillance 2018, 23, 17–00469. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, T.; Diekmann, J.; Knabbe, C.; Dreier, J. Hepatitis E virus blood donor NAT screening: As much as possible or as much as needed? Transfusion 2018, 59, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, S.; Faber, M.; Zimmermann, R.; Bock, C.T.; Offergeld, R.; Steffen, G. Epidemiology of viral hepatitis A to E in Germany. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2022, 65, 149–158. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Sequence | Location at NC_038504 |
|---|---|---|
| HEV-C1_fwd | 5′–TACTGCTAGAGAGGCCCAG–3′ | 49–67 |
| HEV-C1_rvs | 5′–GCTGTATCGGATGCGACC–3′ | 198–215 |
| HEV-C1_prb | 5′–6-FAM-ACCGCCTTTGCTAATGCT-NFQ-MGB–3′ | 86–103 |
| Characteristic | n | % |
|---|---|---|
| Sex | ||
| Female | 98 | 49 |
| Male | 94 | 47 |
| Unknown | 8 | 4 |
| Age group | ||
| ≤9 | 3 | 1.5 |
| 10–19 | 24 | 12 |
| 20–29 | 24 | 12 |
| 30–39 | 24 | 12 |
| 40–49 | 43 | 21.5 |
| 50–59 | 30 | 15 |
| 60–69 | 23 | 11.5 |
| 70–79 | 20 | 10 |
| ≥80 | 1 | 0.5 |
| Unknown | 8 | 4 |
| Place of residence | ||
| North | 26 | 13 |
| West | 48 | 24 |
| East | 25 | 12.5 |
| South | 100 | 50 |
| Unknown | 1 | 0.5 |
| Total | 200 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faber, M.; Wenzel, J.J.; Erl, M.; Stark, K.; Schemmerer, M. No Evidence for Orthohepevirus C in Archived Human Samples in Germany, 2000–2020. Viruses 2022, 14, 742. https://doi.org/10.3390/v14040742
Faber M, Wenzel JJ, Erl M, Stark K, Schemmerer M. No Evidence for Orthohepevirus C in Archived Human Samples in Germany, 2000–2020. Viruses. 2022; 14(4):742. https://doi.org/10.3390/v14040742
Chicago/Turabian StyleFaber, Mirko, Jürgen J. Wenzel, Monika Erl, Klaus Stark, and Mathias Schemmerer. 2022. "No Evidence for Orthohepevirus C in Archived Human Samples in Germany, 2000–2020" Viruses 14, no. 4: 742. https://doi.org/10.3390/v14040742
APA StyleFaber, M., Wenzel, J. J., Erl, M., Stark, K., & Schemmerer, M. (2022). No Evidence for Orthohepevirus C in Archived Human Samples in Germany, 2000–2020. Viruses, 14(4), 742. https://doi.org/10.3390/v14040742