
Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses

 , , , , ,
, , , , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents
2.3. Fermentation, Isolation, and Identification of Exophillic Acid and Its Analogs
2.4. Preparation of Exophillic Acid Derivatives
2.5. HBV Preparation and Infection
2.6. Detection of HBs Antigens
2.7. Immunofluorescence Analysis
2.8. MTT Assay
2.9. HBV Replication Assay
2.10. RT-PCR and Real-Time PCR
2.11. preS1 Binding Assay
2.12. Immunoblot Analysis
2.13. Pull-Down Assay
2.14. NTCP Transporter Assay
2.15. HCV Pseudoparticle Assay
2.16. HDV Infection Assay
3. Results
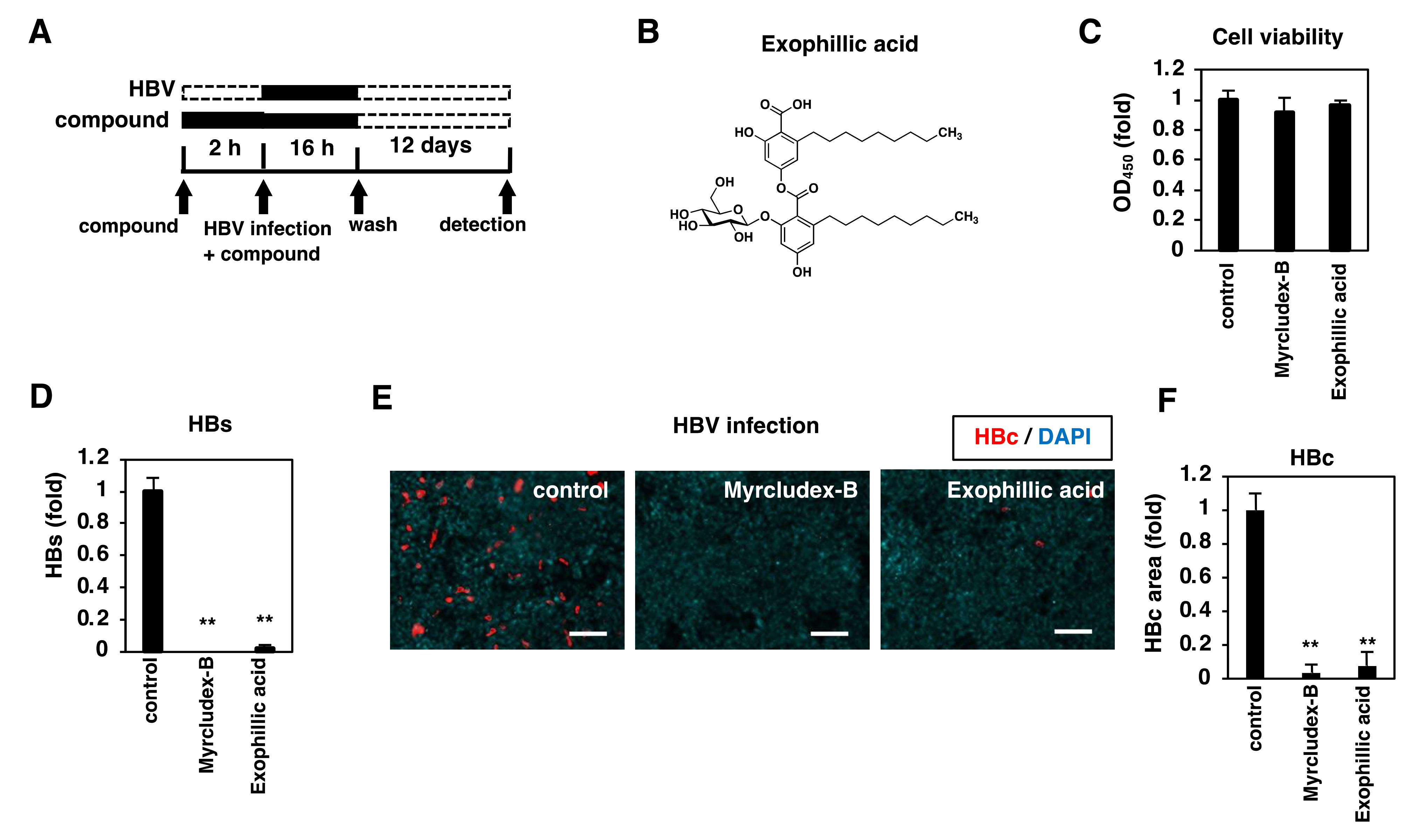
3.1. Exophillic Acid Inhibits HBV Infection
3.2. Anti-HBV Activity of Exophillic Acid in Primary Human Hepatocytes
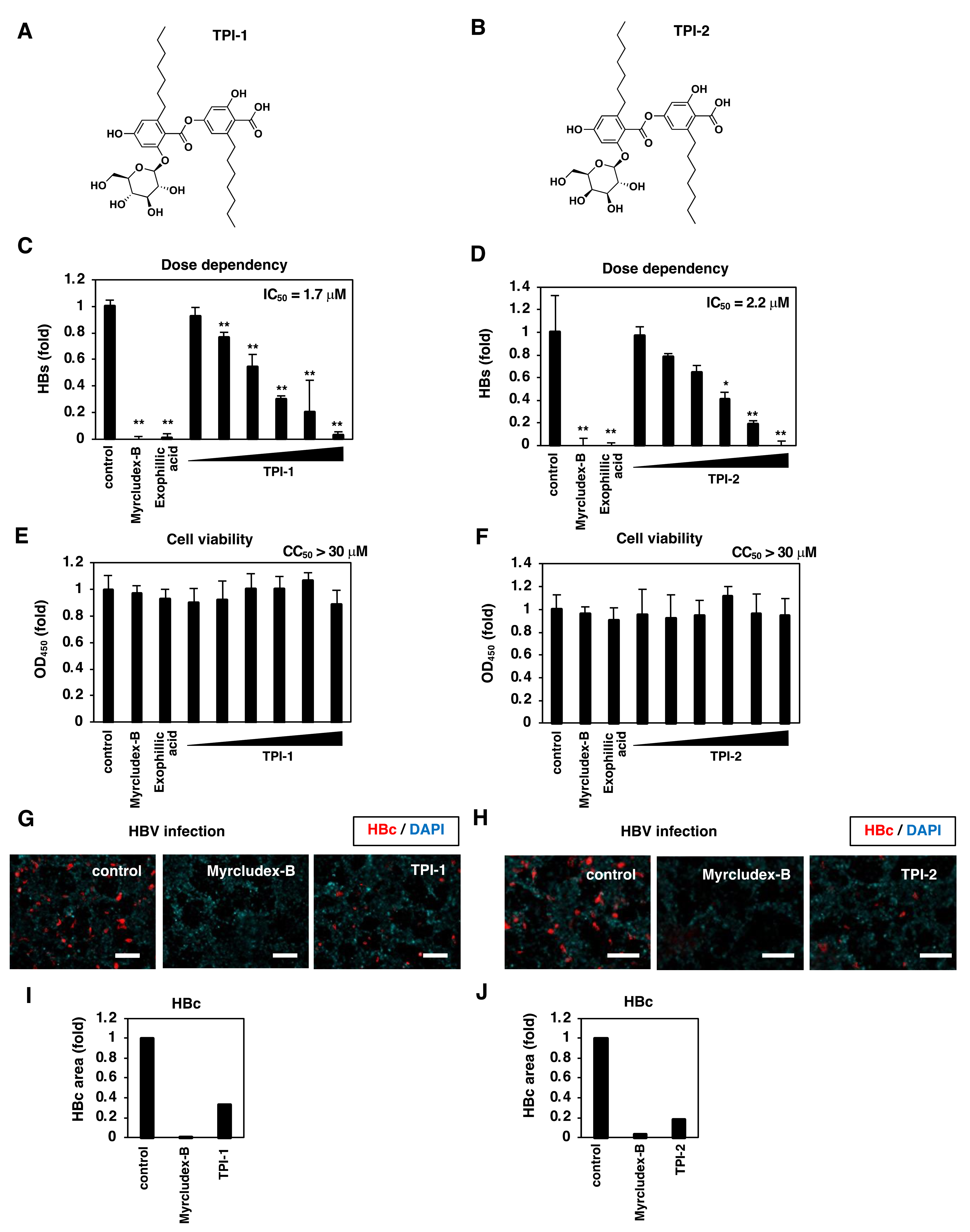
3.3. Exophillic Acid Analogs Inhibit HBV Infection
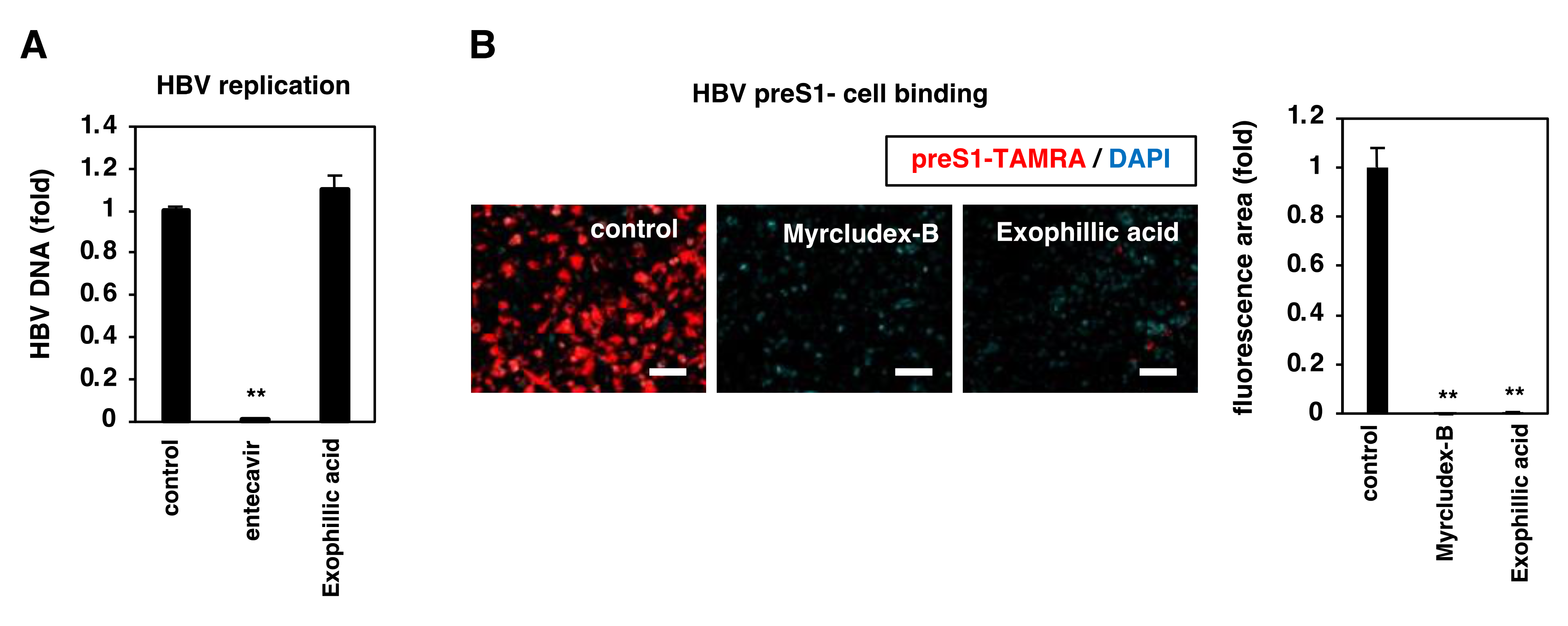
3.4. Exophillic Acid Inhibits HBV Attachment to Target Cells
3.5. Exophillic Acid Targets Host Cells to Reduce HBV Attachment
3.6. Interaction of Exophillic Acid with NTCP
3.7. Exophillic Acid Inhibits the Infection with HDV but Not HCV
3.8. Exophillic Acid Shows Pan-Genotypic Anti-HBV Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Health Organization Hepatitis B-Key Facts. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 21 January 2021).
- Martinez, M.G.; Villeret, F.; Testoni, B.; Zoulim, F. Can we cure hepatitis B virus with novel direct-acting antivirals? Liver Int. 2020, 40, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukuda, S.; Watashi, K. Hepatitis B virus biology and life cycle. Antivir. Res. 2020, 182, 104925. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.H.; et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, M.; Saso, W.; Nishioka, K.; Ohashi, H.; Sugiyama, R.; Ryo, A.; Ohki, M.; Yun, J.H.; Park, S.Y.; Ohshima, T.; et al. The machinery for endocytosis of epidermal growth factor receptor coordinates the transport of incoming hepatitis B virus to the endosomal network. J. Biol. Chem. 2020, 295, 800–807. [Google Scholar] [CrossRef]
- Heuschkel, M.J.; Baumert, T.F.; Verrier, E.R. Cell culture models for the study of hepatitis D virus entry and infection. Viruses 2021, 13, 1532. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, L.; Wedemeyer, H. New treatments for chronic hepatitis B virus/hepatitis D virus infection. Clin. Liver Dis. 2021, 25, 831–839. [Google Scholar] [CrossRef]
- Watashi, K.; Sluder, A.; Daito, T.; Matsunaga, S.; Ryo, A.; Nagamori, S.; Iwamoto, M.; Nakajima, S.; Tsukuda, S.; Borroto-Esoda, K.; et al. Cyclosporin A and its analogs inhibit hepatitis B virus entry into cultured hepatocytes through targeting a membrane transporter, sodium taurocholate cotransporting polypeptide (NTCP). Hepatology 2014, 59, 1726–1737. [Google Scholar] [CrossRef]
- Kaneko, M.; Watashi, K.; Kamisuki, S.; Matsunaga, H.; Iwamoto, M.; Kawai, F.; Ohashi, H.; Tsukuda, S.; Shimura, S.; Suzuki, R.; et al. A novel tricyclic polyketide, vanitaracin A, specifically inhibits the entry of hepatitis B and D viruses by targeting sodium taurocholate cotransporting polypeptide. J. Virol. 2015, 89, 11945–11953. [Google Scholar] [CrossRef] [Green Version]
- Okuyama-Dobashi, K.; Kasai, H.; Tanaka, T.; Yamashita, A.; Yasumoto, J.; Chen, W.; Okamoto, T.; Maekawa, S.; Watashi, K.; Wakita, T.; et al. Hepatitis B virus efficiently infects non-adherent hepatoma cells via human sodium taurocholate cotransporting polypeptide. Sci. Rep. 2015, 5, 17047. [Google Scholar] [CrossRef] [Green Version]
- Tsukuda, S.; Watashi, K.; Hojima, T.; Isogawa, M.; Iwamoto, M.; Omagari, K.; Suzuki, R.; Aizaki, H.; Kojima, S.; Sugiyama, M.; et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology 2017, 65, 1104–1116. [Google Scholar] [CrossRef] [Green Version]
- Shimura, S.; Watashi, K.; Fukano, K.; Peel, M.; Sluder, A.; Kawai, F.; Iwamoto, M.; Tsukuda, S.; Takeuchi, J.S.; Miyake, T.; et al. Cyclosporin derivatives inhibit hepatitis B virus entry without interfering with NTCP transporter activity. J. Hepatol. 2017, 66, 685–692. [Google Scholar] [CrossRef]
- Kaneko, M.; Futamura, Y.; Tsukuda, S.; Kondoh, Y.; Sekine, T.; Hirano, H.; Fukano, K.; Ohashi, H.; Saso, W.; Morishita, R.; et al. Chemical array system, a platform to identify novel hepatitis B virus entry inhibitors targeting sodium taurocholate cotransporting polypeptide. Sci. Rep. 2018, 8, 2769. [Google Scholar] [CrossRef] [Green Version]
- Saso, W.; Tsukuda, S.; Ohashi, H.; Fukano, K.; Morishita, R.; Matsunaga, S.; Ohki, M.; Ryo, A.; Park, S.Y.; Suzuki, R.; et al. A new strategy to identify hepatitis B virus entry inhibitors by AlphaScreen technology targeting the envelope-receptor interaction. Biochem. Biophys. Res. Commun. 2018, 501, 374–379. [Google Scholar] [CrossRef]
- Passioura, T.; Watashi, K.; Fukano, K.; Shimura, S.; Saso, W.; Morishita, R.; Ogasawara, Y.; Tanaka, Y.; Mizokami, M.; Sureau, C.; et al. De novo macrocyclic peptide inhibitors of hepatitis B virus cellular entry. Cell. Chem. Biol. 2018, 25, 906.e5–915.e5. [Google Scholar] [CrossRef]
- Miyakawa, K.; Matsunaga, S.; Yamaoka, Y.; Dairaku, M.; Fukano, K.; Kimura, H.; Chimuro, T.; Nishitsuji, H.; Watashi, K.; Shimotohno, K.; et al. Development of a cell-based assay to identify hepatitis B virus entry inhibitors targeting the sodium taurocholate cotransporting polypeptide. Oncotarget 2018, 9, 23681–23694. [Google Scholar] [CrossRef] [Green Version]
- Fukano, K.; Tsukuda, S.; Watashi, K.; Wakita, T. Concept of viral inhibitors via NTCP. Semin. Liver Dis. 2019, 39, 78–85. [Google Scholar] [CrossRef]
- Ito, K.; Okumura, A.; Takeuchi, J.S.; Watashi, K.; Inoue, R.; Yamauchi, T.; Sakamoto, K.; Yamashita, Y.; Iguchi, Y.; Une, M.; et al. Dual agonist of farnesoid X receptor and takeda G protein-coupled receptor 5 inhibits hepatitis B virus infection in vitro and in vivo. Hepatology 2021, 74, 83–98. [Google Scholar] [CrossRef]
- Ōmura, S. Microbial metabolites: 45 years of wandering, wondering and discovering. Tetrahedron 2011, 67, 6420–6459. [Google Scholar] [CrossRef]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef] [Green Version]
- Ogura, N.; Watashi, K.; Noguchi, T.; Wakita, T. Formation of covalently closed circular DNA in Hep38.7-Tet cells, a tetracycline inducible hepatitis B virus expression cell line. Biochem. Biophys. Res. Commun. 2014, 452, 315–321. [Google Scholar] [CrossRef]
- Watashi, K.; Liang, G.; Iwamoto, M.; Marusawa, H.; Uchida, N.; Daito, T.; Kitamura, K.; Muramatsu, M.; Ohashi, H.; Kiyohara, T.; et al. Interleukin-1 and tumor necrosis factor-alpha trigger restriction of hepatitis B virus infection via a cytidine deaminase activation-induced cytidine deaminase (AID). J. Biol. Chem. 2013, 288, 31715–31727. [Google Scholar] [CrossRef] [Green Version]
- Tsukuda, S.; Watashi, K.; Iwamoto, M.; Suzuki, R.; Aizaki, H.; Okada, M.; Sugiyama, M.; Kojima, S.; Tanaka, Y.; Mizokami, M.; et al. Dysregulation of retinoic acid receptor diminishes hepatocyte permissiveness to hepatitis B virus infection through modulation of sodium taurocholate cotransporting polypeptide (NTCP) expression. J. Biol. Chem. 2015, 290, 5673–5684. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, J.S.; Fukano, K.; Iwamoto, M.; Tsukuda, S.; Suzuki, R.; Aizaki, H.; Muramatsu, M.; Wakita, T.; Sureau, C.; Watashi, K. A single adaptive mutation in sodium taurocholate cotransporting polypeptide induced by hepadnaviruses determines virus species specificity. J. Virol. 2019, 93, 5. [Google Scholar] [CrossRef] [Green Version]
- Bartosch, B.; Dubuisson, J.; Cosset, F.L. Infectious hepatitis C virus pseudo-particles containing functional E1–E2 envelope protein complexes. J. Exp. Med. 2003, 197, 633–642. [Google Scholar] [CrossRef]
- Kuo, M.Y.; Chao, M.; Taylor, J. Initiation of replication of the human hepatitis delta virus genome from cloned DNA: Role of delta antigen. J. Virol. 1989, 63, 1945–1950. [Google Scholar] [CrossRef] [Green Version]
- Sureau, C.; Guerra, B.; Lee, H. The middle hepatitis B virus envelope protein is not necessary for infectivity of hepatitis delta virus. J. Virol. 1994, 68, 4063–4066. [Google Scholar] [CrossRef] [Green Version]
- Ishiyama, A.; Hokari, R.; Nonaka, K.; Chiba, T.; Miura, H.; Otoguro, K.; Iwatsuki, M. Diatretol, an α, α′-dioxo-diketopiperazine, is a potent in vitro and in vivo antimalarial. J. Antibiot. 2021, 74, 266–268. [Google Scholar] [CrossRef] [PubMed]
- Ondeyka, J.G.; Zink, D.L.; Dombrowski, A.W.; Polishook, J.D.; Felock, P.J.; Hazuda, D.J.; Singh, S.B. Isolation, structure and HIV-1 integrase inhibitory activity of exophillic acid, a novel fungal metabolite from Exophiala pisciphila. J. Antibiot. 2003, 56, 1018–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaginuma, S.; Awata, M.; Takada, M.; Kinoshita, K. Physilogically active substance TPI from Fr. Demande. FR 1986, 2579599 A1 19861003. Available online: https://patents.google.com/patent/FR2518546A1/en (accessed on 3 March 2022).
- Meier, A.; Mehrle, S.; Weiss, T.S.; Mier, W.; Urban, S. Myristoylated PreS1-domain of the hepatitis B virus L-protein mediates specific binding to differentiated hepatocytes. Hepatology 2013, 58, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Meertens, L.; Bertaux, C.; Dragic, T. Hepatitis C virus entry requires a critical postinternalization step and delivery to early endosomes via clathrin-coated vesicles. J. Virol. 2006, 80, 11571–11578. [Google Scholar] [CrossRef] [Green Version]
- Tu, T.; Urban, S. Virus entry and its inhibition to prevent and treat hepatitis B and hepatitis D virus infections. Curr. Opin. Virol. 2018, 30, 68–79. [Google Scholar] [CrossRef]
- Testoni, B.; Durantel, D.; Zoulim, F. Novel targets for hepatitis B virus therapy. Liver Int. 2017, 37, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Jeelani, G.; Masuda, Y.; Sakai, K.; Tsukui, K.; Waluyo, D.; Tarwadi; Watanabe, Y.; Nonaka, K.; Matsumoto, A.; et al. Identification of natural inhibitors of Entamoeba histolytica cysteine synthase from microbial secondary metabolites. Front. Microbiol. 2015, 6, 962. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, M.; Sureau, C.; Labonte, P. Use of FDA approved therapeutics with hNTCP metabolic inhibitory properties to impair the HDV lifecycle. Antivir. Res. 2014, 106, 111–115. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, C.; Watanabe, Y.; Oshima, M.; Hirose, T.; Yamasaki, M.; Iwamoto, M.; Iwatsuki, M.; Asami, Y.; Kuramochi, K.; Wakae, K.; et al. Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses. Viruses 2022, 14, 764. https://doi.org/10.3390/v14040764
Kobayashi C, Watanabe Y, Oshima M, Hirose T, Yamasaki M, Iwamoto M, Iwatsuki M, Asami Y, Kuramochi K, Wakae K, et al. Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses. Viruses. 2022; 14(4):764. https://doi.org/10.3390/v14040764
Chicago/Turabian StyleKobayashi, Chisa, Yoshihiro Watanabe, Mizuki Oshima, Tomoyasu Hirose, Masako Yamasaki, Masashi Iwamoto, Masato Iwatsuki, Yukihiro Asami, Kouji Kuramochi, Kousho Wakae, and et al. 2022. "Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses" Viruses 14, no. 4: 764. https://doi.org/10.3390/v14040764
APA StyleKobayashi, C., Watanabe, Y., Oshima, M., Hirose, T., Yamasaki, M., Iwamoto, M., Iwatsuki, M., Asami, Y., Kuramochi, K., Wakae, K., Aizaki, H., Muramatsu, M., Sureau, C., Sunazuka, T., & Watashi, K. (2022). Fungal Secondary Metabolite Exophillic Acid Selectively Inhibits the Entry of Hepatitis B and D Viruses. Viruses, 14(4), 764. https://doi.org/10.3390/v14040764